
Figure 4.
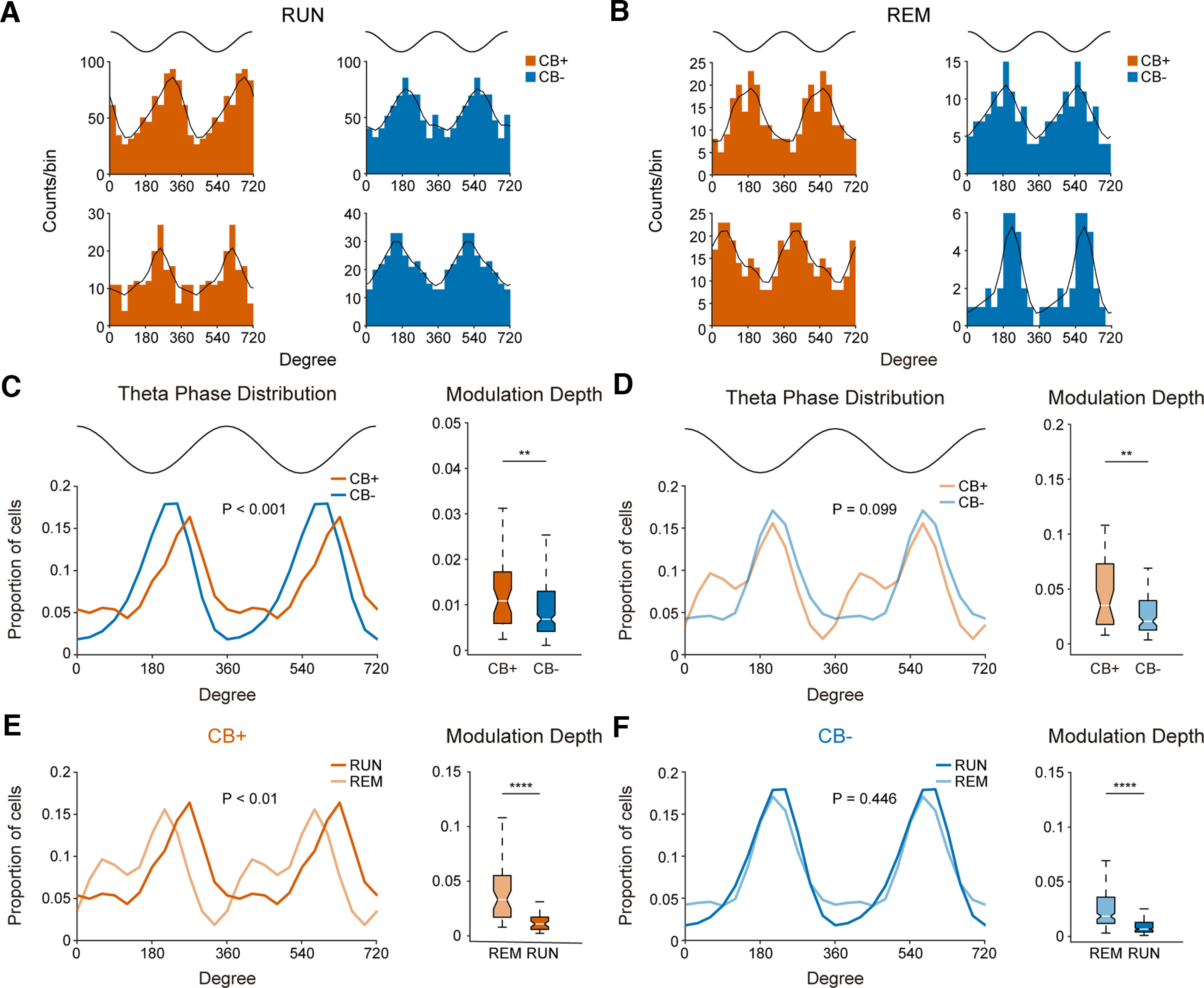
A subset of CB+ PNs shifted their theta firing phase during REM sleep. A, Example theta firing phase distribution of both PN subtypes during RUN state (theta peak = 0°, 360°, theta trough = 180°; bin size: 30°). B, Same as in A, but for preferred theta firing phase during REM sleep theta oscillations. C, Left, Distribution of preferred theta phase of both PN subtypes during RUN state (CB+ PNs, n = 43; CB− PNs, n = 199; p = 2.2e-4, Watson’s U2 test; bin size: 30°). Top trace indicates idealized reference theta cycle. Right, Theta modulation depth of CB+ and CB− PNs during RUN (**p = 8.6e-3 Mann–Whitney test). D, Same as in C, but for population distribution of preferred theta firing phase during REM sleep. Note the bimodal distribution of theta phase preference of CB+ PNs (p = 0.099, Watson’s U2 test; bin size: 30°). Right, Theta modulation depth of both PN subtypes during REM sleep (**p = 1.4e-3, Mann–Whitney test). E, Left, Comparison of preferred theta firing phase of CB+ PNs during RUN and REM states. The preferred theta firing phase shifted significantly between the two states (p = 9.1e-3, Watson’s U2 test, bin size: 30°). Right, Modulation depth of CB+ PNs during REM sleep theta is significantly deeper than RUN theta states (****p = 2.3e-8, Mann–Whitney test). F, Same as in E, but for comparison of preferred theta firing phase of CB− PNs during RUN and REM states. No significant theta phase shift was observed between the two theta states (p = 0.446, Watson’s U2 test, bin size: 30°). Right, Theta modulation depth of CB− PNs during RUN and REM states (****p = 1.7e-26, Mann–Whitney test).