

Abstract
Introduction
This study aimed to examine the anti-biofilm activity and mechanism of gallic acid (GA), kaempferol-7-O-glucoside (K7G) and apigenin-7-O-glucoside (A7G) against Staphylococcus aureus and Escherichia coli.
Methods
The antibacterial activity of the natural compounds was determined by serial dilution method. The inhibitory activity of natural compounds on biofilms was determined by crystal violet staining method. The effects and mechanisms of natural compounds on bacterial biofilms were analyzed by atomic force microscopy.
Results
In our study, compared with GA and K7G, A7G was found to exhibit the strongest anti-biofilm and antibacterial activities. The minimum biofilm inhibitory concentration (MBIC) of A7G against S. aureus and E. coli was 0.20 mg/mL and 0.10 mg/mL, respectively. The inhibition rates of 1/2 MIC of A7G on biofilms of S. aureus and E. coli were 88.9%, and 83.2% respectively. Moreover, atomic force microscope (AFM) images showed the three-dimensional biofilm morphology of S. aureus and E. coli, and the results indicated that A7G was highly effective in biofilm inhibition.
Discussion
It was found that the inhibition of A7G on biofilm was achieved through inhibiting on exopolysaccharides (EPS), quorum sensing (QS), and cell surface hydrophobicity (CSH). A7G exerted strong anti-biofilm activities by inhibiting EPS production, QS, and CSH. Hence, A7G, as a natural substance, could be a promising novel antibacterial and anti-biofilm agent for control of biofilm in food industry.
Keywords: apigenin-7-O-glucoside, anti-biofilm, exopolysaccharide, quorum sensing, cell surface hydrophobicity, atomic force microscope
Introduction
Food safety is closely related to public health and has received increasing attention from researchers in recent years. Food-borne spoilage and new food-borne diseases caused by pathogens are one of the major food safety challenges.1 A potential cause of microbial contamination in food is via their presence in biofilms either on the raw material2,3 or formed during processing in a food production environment.4,5 Bacterial biofilms are amorphous and dynamic mono- or multi-microbial structures attached to living or non-living surfaces.6–8 To develop biofilm, microbial communities encased in a layer of exopolysaccharides (EPS), adhered to biotic or abiotic surfaces.9,10 The typical biofilm formation begins with the adhesion of planktonic bacteria, followed by cell growth and EPS production.11 Biofilm formation of microbial communities is influenced by QS. It has been reported that QS plays a central role in bacterial social life.12,13 Besides, the ability of bacteria attached to the interface is directly related to biofilm formation. Cell surface hydrophobicity, appears to be an important factor in bacterial adhesion.14,15 Therefore, EPS production, \quorum sensing system and the ability of bacteria to adhere to surfaces all affect the formation of biofilm.
Staphylococcus aureus is a common foodborne pathogen. As a typical representative of the QS system of Gram-positive bacteria, its Accessory gene regulator (Agr) QS system regulates the secretion of virulence and the formation of biofilm through a two-component system. Pathogenic Escherichia coli regulates virulence secretion and cell colonization through QS system, causing intestinal metabolism and immune imbalance, leading to bacterial diarrhea, dysentery and other diseases. Escherichia coli can regulate the biofilm formation ability of bacteria and the synthesis of virulence factors through QS system, so as to promote the pathogen to overcome the host immune system, spread virulence factors and cause infectious diarrhea and acute enteritis. EPS allows the bacteria in the biofilm to gather together and play a stabilizing and protective role. EPS can affect water content, density, adsorption properties, porosity, charge and other properties in biofilms, so EPS can directly affect the environment necessary for bacterial life.16 The proportion of components in EPS is closely related to the type of biofilm.17 In S. aureus, EPS includes polysaccharides, proteins, glycoproteins, glycolipids and so on.
To minimize the risk of contamination from S. aureus and E. coli, many synthetic additives are used in industrial food processing. However, consumers might be worried about being exposed to synthetic chemicals in their daily diet and the potential side effects of some synthetic additives, such as sodium benzoate, potassium sorbate and phosphoric acid.18–20 Therefore, from this perspective, the search for novel, safer and more effective strategies for controlling food pathogens in food processing plants remains a very relevant issue.21,22 Natural antimicrobial agents are a potentially interesting solution for treatment and prevention of biofilm formation on the surface of food and processing surfaces, particularly the extracts from edible plant tissues because these extracts are considered almost non-toxic. Our previous study20 showed the inhibitive activity of phenolics of peony flowers in the eluent fraction of silica gel column chromatography (PPF-ESGCC) on bacterial biofilm was high, and phenolics were mainly composed of apigenin-7-O-glucoside (A7G), gallic acid (GA) and kaempferol-7-O-glucoside (K7G). However, the information how the phenolics in PPF-ESGCC contribute to anti-biofilm activity and the action mechanism of phenolics needs further study.
Therefore, in this study, the antibacterial activities and anti-biofilm activities of A7G, GA and K7G were compared, and the substance with the strongest anti-biofilm activity was selected. Then the inhibitory mechanism of sub-minimum inhibitory concentration (sub-MIC) of the substance with the strongest anti-biofilm activity was studied.
Materials and Methods
Materials
Apigenin-7-O-glucoside (A7G, 98%), gallic acid (GA, 98%) and kaempferol-7-O-glucoside (K7G, 98%) were purchased from Shanghai Yuanye Co., Ltd, China. Escherichia coli ATCC 25922 (GIM1.559) and Staphylococcus aureus ATCC 6538 (GIM1.142) were purchased from Guangdong Microbial Species Preservation Center, China. Chromobacterium violaceum CV026 was purchased from Spanish Type Culture Collection (CECT-5999).
Determination of Minimum Inhibitory Concentration (MICs)
Determination of MICs was carried out according to previous report.23 A7G, GA and K7G were added to the wells of a sterile 96-well plate, and serially diluted in LB broth, and their concentrations ranged from 0.14 to 6.84mg/mL. And then each well was inoculated 2% of bacterial suspension (106 cfu/mL), while the culture medium without the bacterial suspension was used as control, and cultured at 37°C for 24 h. The samples were taken at 600 nm at every two hours using a microplate reader (Molecular Devices, M5, USA) to measure the absorbance. MIC refers to the lowest concentration that inhibits bacterial growth after 24 hours of incubation at 37 °C.
Determination of Growth Inhibitory Activity of Apigenin-7-O-Glucoside (A7G)
The growth rates of Escherichia coli ATCC 25922 and Staphylococcus aureus ATCC 6538 treated with 1/2 MIC of A7G and untreated Escherichia coli ATCC 25922 and Staphylococcus aureus ATCC 6538 were determined24 (Yang et al 2016). Briefly, LB broth with and without 1/2 MIC of A7G were incubated at 37°C for 24 h. Then, the samples were taken every two hours for measuring the absorbance at 600 nm with a microplate reader (Molecular Devices, M5, USA).
Biofilm Formation Inhibition Assay
Bacterial biofilm inhibition rate was measured using the 24-well plate method.25,26 The lowest sample concentration at which the bacterial biofilm inhibition rate was 100% was defined as the minimum concentration on biofilm inhibition (MBIC). Once inoculated with 10 μL of bacterial cultures (106 cfu/mL), LB broth with altered concentrations of A7G, GA and K7G was incubated at normal room temperature for 24 hours. Deionized water was used to remove the lightly attached cells after incubation while the cells stacked on the wall were tainted for 10 minutes with 0.3% crystal violet solution. The excessive stains were removed with deionized water, after which an addition of 1 mL of 95% ethanol was applied to melt the crystal violet and then incubated at normal room temperature for 30 minutes, and its absorption was measured at 595 nm. To determine the inhibition rate, the formula below was employed (Eq. (1)):
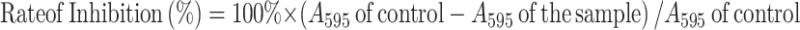 |
Quantification of Violacein Production
Because Chromobacterium violaceum CV026 is a mature indicator to evaluate the QS inhibitory activity of natural components against bacteria, CV026 was used to determine the quorum-sensing inhibitory activity of A7G.27 Since most Gram-negative bacteria have N-Acyl homoserine lactone (AHL) quorum-sensing system, the use of CV026 to evaluate the quorum-sensing inhibitory activity of A7G has certain universality. CV026 was grown in 10 mL LB in test tubes with different A7G levels, C6-HSL was introduced and then grown 24 hours in 200 rpm shakers at 37°C. The 1 mL of material was put into a centrifugal tube (5 mL), and centrifuged at 13,000 rpm / min for 10 minutes. Afterwards the supernatant was discarded and 500 μL DMSO was applied to the pellet, and then a vortex shaker aided in dissolving the violacein. The mixture was eventually centrifuged, and its absorbance values were measured at 585 nm for the supernatant containing violacein.28
Exopolysaccharides (EPS) Production Inhibition Assay
The inhibition rate of EPS production in A7G was determined according to the method defined by Huston.29 Inoculated with 1% bacterial cultures (106 cfu/mL), the LB broth with and without different concentrations of A7G was incubated at 37°C for 24 hours. After incubation, 1 mL of the mixture was centrifuged for 10 min (15,000 rpm, 4 °C), then the supernatant (0.22 μm) was filtered. The associated proteins in the supernatant were removed, using the Sevag method.30 After removing the Sevag reagent, four volumes frozen ethanol was added to the water phase and stored at 4°C for 24 hours to precipitate the EPS. The precipitated EPS were centrifuged at 15,000 rpm (4°C) for 10 min, and the supernatant was discarded. Deionized water (1 mL) was added to dissolve the EPS. The EPS content was calculated at 490 nm using phenol-sulfuric acid procedure with glucose as the reference. The standard curve equation was computed as y=0.0581x+0.0913 (R2= 0.9969). The inhibition rate for the EPS was calculated using the following equation (Eq.(2)):
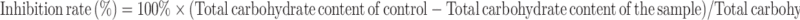 |
Cell Surface Hydrophobicity (CSH) Assay
The cell surface hydrophobicity was measured as published previously by Zhang et al.31 Briefly, for each strain tested, 4 mL of cell suspension was placed into a clean glass tube and centrifuged for 10 min (15,000 rpm, 4°C). Sediments were washed twice by PBS (pH = 7.0) and overlaid with 1 mL of toluene. The samples were vortexed thoroughly for 3 min and then the phases were allowed to separate. The aqueous phase was removed and was measured to determine the cell density in the phase. Each strain was tested in triplicate. The relative CSH was calculated according to the following equation (Eq. (3)):
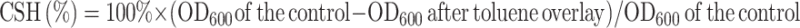 |
Atomic Force Microscope (AFM) Analysis of Bacterial Biofilm
The preparation of bacterial biofilms was carried out according to the method of Nithya.32,33 The specific procedure was as follows: 1 mL of LB broth with different concentrations of apigenin-7-O-glucoside (1/10 MIC to 1/2 MIC) was added to the sterile 24-well plates containing sterile silicon chip (5 mm×5 mm), and 1% of bacterial cultures was inoculated at 37 °C for 24 h. After incubation, the silicon chips were rinsed 3 times with 0.9% NaCl solution to remove planktonic cells on the silicon chips, and finally the biofilms were observed by AFM (MuLtimode 8, Germany).
Statistical Analysis
The experiments were repeated at least three times. Data is expressed as mean ± SD (n = 3). ANOVA and Tukey’s Single-way Test (SPSS 22) was used to determine significant differences. Differences were deemed significant at P < 0.05.
Results and Discussion
MICs and MBICs of A7G, GA and K7G
In our study, MICs of A7G, GA and K7G were determined, and the results are shown in Table 1. The MIC of A7G for E. coli and S. aureus are 0.14 mg/mL, and 0.28 mg/mL, respectively, and the MIC of A7G was lower than that of GA and K7G. Hence, A7G exhibited stronger antibacterial activities.
Table 1.
MICs and MBICs of A7G, GA and K7G (Unit: mg/mL)
| Components | E. coli | S. aureus | ||
|---|---|---|---|---|
| MIC | MBIC | MIC | MBIC | |
| A7G | 0.14 | 0.10 | 0.28 | 0.20 |
| GA | 2.00 | 1.50 | 4.00 | 2.50 |
| K7G | 1.71 | 1.00 | 3.42 | 2.00 |
In addition, the MBICs of A7G, GA and K7G were also determined. The MBIC of A7G for E. coli and S. aureus are 0.10 mg/mL, and 0.20 mg/mL, respectively, and the value is lower than the MBICs of K7G and GA. Among three compounds, GA with a MBIC (2.50 mg/mL) for S. aureus exhibited the weakest anti-biofilm activities. Therefore, A7G exhibited the strongest anti-biofilm activities and A7G was chosen for following experiment.
In our previous study, the phenolics extracted from peony flowers can effectively inhibit biofilm formation, and phenolics are mainly composed of apigenin-7-O-glucoside (A7G), gallic acid (GA) and kaempferol-7-O-glucoside (K7G). However, the information how these phenolics contribute to the anti-biofilm activity were not evaluated. In Table 1, the MIC of A7G is consistent with the experimental results of Cottigli et al.34 GA with a MIC (2 mg/mL) for E. coli and S. aureus35,36 and K7G with a MIC (2 mg/mL) for E. coli and S. aureus were consistent with our results. Moreover, MBIC of GA is lower than that of previous reports that the MBIC of GA for S. aureus was 4 mg/mL,36 and there is few reports about MBIC of A7G and K7G. In this paper, the MBIC of A7G and K7G were studied, and the results indicated that A7G possessed more effective anti-microbial and anti-biofilm activities against S. aureus and E. coli, compared to GA and K7G. Hence, A7G was chosen for subsequent experiment.
Effect of A7G on Bacterial Growth
In order to evaluate whether the inhibitory effect of A7G on bacterial biofilm is through inhibiting the growth of bacteria, the effect of A7G on the growth of bacteria was studied. As shown in Figure 1, after incubation at 37°C for 12 h, the growth of E. coli and S. aureus treated with and without 1/2 MIC of A7G was stable, which showed that 1/2 MIC of A7G had no effect on the growth of E. coli and S. aureus, and it was consistent with previous reports.24,37
Figure 1.

Effect of A7G on bacterial growth and biofilm formation of E. coli and S. aureus at different concentrations. (A) Effect on bacterial growth of E. coli, (B) effect on bacterial growth of S. aureus, (C) effect on biofilm formation Bars labeled with different letters indicated significant differences at P < 0.05.
It was found that the inhibitory effect of A7G on bacterial biofilm is not through inhibiting the growth of bacteria (Figure 1). Hence, sub-MICs of A7G were selected for the following experiment.38,39
Inhibitory Effect of Sub-MICs of A7G on Biofilm Formation
The results of different sub-MICs of A7G which influenced biofilm formation activities of E. coli and S. aureus is showed in Figure 1. For S. aureus and E. coli, 1/2 MIC of A7G had better inhibitory effect on biofilm formation at 88.9%, and 83.2%, and as the concentration of A7G increased, the biofilm inhibition rate also increased. The results indicated that A7G had a strong anti-biofilm activity against S. aureus and E. coli.
Inhibition Mechanism of A7G on Biofilm Formation
Quorum Sensing (QS) Inhibition
In order to evaluate the inhibition mechanism of A7G on bacterial biofilm, the effect of A7G on QS was studied (Figure 2). In Figure 2A, sub-MICs of A7G have no effect on the growth of CV026. The results indicated that the inhibitory effect of sub-MICs of A7G on the bacterial quorum sensing is not produced by inhibiting the growth of CV026. Figure 2B showed that with A7G concentration changing from 1/10 MIC to 1/2 MIC, the inhibition on violacein production increased significantly (P < 0.05) and reached 81.50% at the 1/2 MIC of A7G.
Figure 2.

The inhibitory effect of A7G on Quorum sensing of CV026. (A) Effect of A7G on CV026 growth; (B) inhibition on violacein production of CV026 by A7G. Bars labeled with different letters indicated significant differences at P < 0.05.
CV026 and E. coli have similar N-Acyl homoserine lactone (AHL) mediated quorum-induction system,40 meanwhile, cv026 and Staphylococcus aureus and Escherichia coli all have Autoinducer 2 (AI-2) mediated quorum-induction system.40,41 A7G showed significant inhibitory activity against QS of quorum-sensing indicator bacterium CV026, so A7G may have certain inhibitory effects on quorum-sensing of Escherichia coli and Staphylococcus aureus.
Biofilm development processes are regulated by QS,42,43 and there is increasing evidence that QS plays a crucial role in the maturation of biofilms.44 A7G was shown to have a considerable influence on bacterial QS (Figure 2). The results suggested that A7G could effectively inhibit the production of violacein. In the same manner as in the studies by Yin et al45 and by Choo et al,46 the results indicated that phenolics can inhibit violacein production. Vikram et al47 studied the suppression of bacterial cell–cell signalling, biofilm formation and type III secretion system by citrus flavonoids, and they also found a potential modulation of bacterial cell–cell communication, E. coli O157:H7 biofilm and V. harveyi virulence, by flavonoids especially naringenin, and apigenin. Therefore, A7G can be a potential quorum sensing inhibitor (QSI) to influence biofilm formation.
Inhibition of EPS Production
As shown in Figure 3, the EPS production of E. coli and S. aureus decreased significantly (P < 0.05) in the presence of A7G. And at a concentration of 1/2 MIC A7G, the EPS of S. aureus and E. coli was inhibited by 74.5% and 71.2%, respectively.
Figure 3.

Inhibition of A7G (1/10, 1/5, and 1/2 MIC) on EPS production of E. coli and S. aureus. Bars labeled with different letters (a, b, c or A, B, C) indicated significant differences at P < 0.05.
It has been proved that EPS plays an important role in the maintenance of biofilm structure.10,48 Biofilms are mainly composed of extracellular polymers secreted by microorganisms and themselves, and the main components of extracellular polymers are polysaccharide protein complexes. In the process of biofilm formation, ESPs play an important role, such as the adhesion of microorganisms on the carrier surface, the aggregation between microorganisms, and the maintenance of biofilm structure.48 In addition, the overproduction of EPS results in the change of biofilm structure, which is related to the increase of cell resistance to osmotic pressure and oxidative stress. In the present study, the total amount of biofilm EPS was also reduced with the increasement of A7G concentration (Figure 3). Moola et al reported that phenolics disrupted biofilms by extracting the polysaccharides, which are essential for the biofilm formation. It is likely that E. coli and S. aureus biofilms are also disrupted through similar mechanisms. To the best of our knowledge, this is the first report about the effect of A7G on EPS production.
Effect of A7G on Cell Surface Hydrophobicity
To explore how AG inhibits biofilm formation, the effect of A7G on cell surface hydrophobicity was investigated. Figure 4 shows that a high and statistically significant decrease (P < 0.05) occurred in percentages of cell surface hydrophobicity with the increasing of A7G concentrations.
Figure 4.

Effect of sub-MICs of A7G (1/10 MIC, 1/5 MIC, 1/2 MIC) on cell surface hydrophobicity. Bars labeled with different letters (a, b, c or A, B, C) indicated significant differences at P < 0.05.
Cell surface hydrophobicity (CSH) is identified as an important virulence factors that have significant roles in the pathogenesis of S. aureus and E. coli. Previous studies49 have shown positive correlation between CSH and adhesion of S. aureus and E. coli. The adhesion of S. aureus and E. coli is an initial and critical step in biofilm formation. The present study showed the ability of A7G to influence CSH of S. aureus and E. coli in a dose-dependent manner (Figure 4). In the studies of Nithyanand et al50 and Polaquini et al,51 their results obtained in the hydrophobicity assays showed a decrease in the CSH of the two strains after the treatment by bacteriostats. The results indicated that lower hydrophobicity resulted in less bacterial adhesion and less biofilm. Thus, A7G can inhibit biofilm formation through reducing CSH.
Atomic Force Microscope (AFM) Analysis of Bacterial Biofilm
The attachment of S. aureus and E. coli cells to abiotic substrate and the cell-to-cell attachment in the biofilms were examined by AFM. The AFM images reveal differences in surface morphology, as shown in Figures 5 and 6. Figures 5 and 6 show three-dimensional topography, deflection image, AFM topography, and line profile of height, which is indicated in topography, and the control groups show higher thickness of biofilm. Compared to other groups, the control groups have 60.00 ± 26.28 nm of average height for S. aureus biofilm and 52.00 ± 2.208 nm of average height for E. coli biofilm (Table 2). In Figures 5B and 6B, it appears that cells are aggregated with EPS. Moreover, in Figure 5D, the line profile indicated some of single bacteria attached to the substrate, showing 60–80 nm height, and in Figure 5D, it produced its highest peak (83 nm).
Figure 5.

Three-dimensional topography, deflection image, AFM topography, and height line profile of S. aureus biofilm. (A) 3-D topography of biofilm without A7G, (B) deflection image of biofilm without A7G, (C) AFM topography of biofilm without A7G, (D) height profile of biofilm without A7G, (E) 3-D topography of biofilm treated by A7G at 1/10 MIC, (F) deflection image of biofilm treated by A7G at 1/10 MIC, (G) AFM topography of biofilm treated by A7G at 1/10 MIC, (H) height profile of biofilm treated by A7G at 1/10 MIC, (I) 3-D topography of biofilm treated by A7G at 1/5 MIC, (J) deflection image of biofilm treated by A7G at 1/5 MIC, (K) AFM topography of biofilm treated by A7G at 1/5 MIC, (L) height profile of biofilm treated by A7G at 1/5 MIC, (M) 3-D topography of biofilm treated by A7G at 1/2 MIC, (N) deflection image of biofilm treated by A7G at 1/2 MIC, (O) AFM topography of biofilm treated by A7G at 1/2 MIC, (P) height profile of biofilm treated by A7G at 1/2 MIC. All the images show an area of 1×1 μm2.
Figure 6.

Three-dimensional topography, deflection image, AFM topography, and height line profile of E. coli biofilm. (A) 3-D topography of biofilm without A7G, (B) deflection image of biofilm without A7G, (C) AFM topography of biofilm without A7G, (D) height profile of biofilm without A7G, (E) 3-D topography of biofilm treated by A7G at 1/10 MIC, (F) deflection image of biofilm treated by A7G at 1/10 MIC, (G) AFM topography of biofilm treated by A7G at 1/10 MIC, (H) height profile of biofilm treated by A7G at 1/10 MIC, (I) 3-D topography of biofilm treated by A7G at 1/5 MIC, (J) deflection image of biofilm treated by A7G at 1/5 MIC, (K) AFM topography of biofilm treated by A7G at 1/5 MIC, (L) height profile of biofilm treated by A7G at 1/5 MIC, (M) 3-D topography of biofilm treated by A7G at 1/2 MIC, (N) deflection image of biofilm treated by A7G at 1/2 MIC, (O) AFM topography of biofilm treated by A7G at 1/2 MIC, (P) height profile of biofilm treated by A7G at 1/2 MIC. All the images show an area of 1×1 μm2.
Table 2.
The Average Height of E. coli and S. aureus Biofilm at Different Concentrations of A7G (0, 1/10, 1/5, and 1/2 MIC)
| Samples | Average Height (nm) | |
|---|---|---|
| S. aureus | E. coli | |
| Control | 60.00 ± 26.28 | 52.00 ± 2.20 |
| 1/10 MIC | 11.40 ± 3.39 | 11.70 ± 3.60 |
| 1/5 MIC | 5.31 ± 0.32 | 6.52 ± 0.51 |
| 1/2 MIC | 3.34 ± 0.65 | 2.43 ± 0.34 |
To remove the biofilm on the silicon, we applied different concentrations of A7G (1/10, 1/5, and 1/2 MIC) in the process of incubation. Figures 5F-N and 6F-N show that the number of bacteria on the silicon is fewer than the control groups. As the concentrations of A7G increased, it was found that less number of bacterial cells attached to the surface. This phenomenon might be attributed to extracellular polymers which were inhibited during incubation, and the loss of extracellular polymers on bacterial surface induced detachment of bacteria from the silicon.52
The images obtained by AFM, shown in Figures 5 and 6, confirm the use of AFM as a tool for the visualisation of biofilm. The images clearly show how A7G inhibits the formation of S. aureus and E. coli biofilm at concentrations of 1/10, 1/5, and 1/2 MIC. Bonez et al reported the similar AFM results of biofilm which treated with different concentrations of A22 compound.53
In China, peony is a new food raw material, so A7G isolated from peony may have high safety. The results suggest that A7G, as a natural and safe substance, could be potentially used to control the food contaminations caused by pathogenic bacteria, thus providing an effective natural antibiofilm alternative for food industry.
Acknowledgment
This work was supported by national first-class discipline program of Food Science and Technology (JUFSTR20180204) and 13th 5 years National Key Research and Development Program Project: Research on Core Technologies of Green Processing of Tea Food and Product Creation (2017YFD0400803).This work was supported by the Open Project Program of State Key Laboratory of Dairy Biotechnology (No. SKLDB2022-002).
Data Sharing Statement
The original contributions presented in the study are included in the article, further inquiries can be directed to the corresponding authors.
Ethical Approval
This article does not contain any studies with animals performed by any of the authors.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Disclosure
The authors declare that they have no conflict of interest.
References
- 1.Zhang YB, Liu XY, Wang YF, et al. Antibacterial activity and mechanism of cinnamon essential oil against Escherichia coli and Staphylococcus aureus. Food Control. 2016;59:282–289. doi: 10.1016/j.foodcont.2015.05.032 [DOI] [Google Scholar]
- 2.Kubota H, Senda S, Nomura N, et al. Biofilm Formation by Lactic Acid Bacteria and Resistance to Environmental Stress. J Biosci Bioeng. 2008;106:381–386. doi: 10.1263/jbb.106.381 [DOI] [PubMed] [Google Scholar]
- 3.Hossain MI, Rahaman Mizan MF, Toushik SH, et al. Antibiofilm effect of nisin alone and combined with food-grade oil components (thymol and eugenol) against Listeria monocytogenes cocktail culture on food and food-contact surfaces. Food Control. 2022;135:108796. doi: 10.1016/j.foodcont.2021.108796 [DOI] [Google Scholar]
- 4.Somers EB, Johnson ME, Wong ACL. Biofilm formation and contamination of cheese by nonstarter lactic acid bacteria in the dairy environment. J Dairy Sci. 2001;84:1926–1936. doi: 10.3168/jds.S0022-0302(01)74634-6 [DOI] [PubMed] [Google Scholar]
- 5.Kirtonia K, Salauddin M, Bharadwaj KK, et al. Bacteriocin: a new strategic antibiofilm agent in food industries. Biocatalysis Agr Biotechnol. 2021;36:102141. doi: 10.1016/j.bcab.2021.102141 [DOI] [Google Scholar]
- 6.Prateeksha SK, Singh BN. Nanoemulsion-loaded hydrogel coatings for inhibition of bacterial virulence and biofilm formation on solid surfaces. Sci Rep-UK. 2019;9:548. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Kirtonia K, Salauddin M, Bharadwaj KK, et al. Bacteriocin: a new strategic antibiofilm agent in food industries. Biocatal Agr Biotech. 2021;36. [Google Scholar]
- 8.Hossain MI, Mizan MFR, Toushik SH, et al. p Antibiofilm effect of nisin alone and combined with food-grade oil components (thymol and eugenol) against Listeria monocytogenes cocktail culture on food and food-contact surfaces. Food Control. 2022;1:135. [Google Scholar]
- 9.Frederick MR, Kuttler C, Hense BA, et al. A mathematical model of quorum sensing regulated EPS production in biofilm communities. Theor Biol Med Model. 2011;8. doi: 10.1186/1742-4682-8-8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Liu F, Jin P, Gong H, et al. Antibacterial and antibiofilm activities of thyme oil against foodborne multiple antibiotics-resistant Enterococcus faecalis. Poult Sci. 2020;99:5127–5136. doi: 10.1016/j.psj.2020.06.067 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Liu S, Cao S, Guo J, et al. Graphene oxide-silver nanocomposites modulate biofilm formation and extracellular polymeric substance (EPS) production. Nanoscale. 2018;10:19603–19611. doi: 10.1039/C8NR04064H [DOI] [PubMed] [Google Scholar]
- 12.Ponnusamy K, Paul D, Kweon JH. Inhibition of Quorum Sensing Mechanism and Aeromonas hydrophila Biofilm Formation by Vanillin. Environ Eng Sci. 2009;26:1359–1363. doi: 10.1089/ees.2008.0415 [DOI] [Google Scholar]
- 13.Rashmi M, Meena H, Meena C, et al. Anti-quorum sensing and antibiofilm potential of Alternaria alternata, a foliar endophyte of Carica papaya, evidenced by QS assays and in-silico analysis. Fungal Biol. 2018;122:998–1012. doi: 10.1016/j.funbio.2018.07.003 [DOI] [PubMed] [Google Scholar]
- 14.Anil S, Ellepola AN, Samaranayake LP. The impact of chlorhexidine gluconate on the relative cell surface hydrophobicity of oral Candida albicans. Oral Dis. 2001;7:119–122. doi: 10.1034/j.1601-0825.2001.70210.x [DOI] [PubMed] [Google Scholar]
- 15.Zammuto V, Rizzo MG, Spanò A, et al. In vitro evaluation of antibiofilm activity of crude extracts from macroalgae against pathogens relevant in aquaculture. Aquaculture. 2022;549:737729. doi: 10.1016/j.aquaculture.2021.737729 [DOI] [Google Scholar]
- 16.Villain M, Simon S, Bourven I, et al. The use of a new mobile phase, with no multivalent cation binding properties, to differentiate extracellular polymeric substances (EPS), by size exclusion chromatography (SEC), from biomass used for wastewater treatment. Process Biochemistry. 2010;45:1415–1421. doi: 10.1016/j.procbio.2010.05.018 [DOI] [Google Scholar]
- 17.da Silva EP, De Martinis ECP. Current knowledge and perspectives on biofilm formation: the case of Listeria monocytogenes. Appl Microbiol Biotechnol. 2013;97:957–968. doi: 10.1007/s00253-012-4611-1 [DOI] [PubMed] [Google Scholar]
- 18.Di Bonaventura G, Stepanovic S, Picciani C, et al. Effect of environmental factors on biofilm formation by clinical Stenotrophomonas maltophilia isolates. Folia Microbiol (Praha). 2007;52:86–90. doi: 10.1007/BF02932144 [DOI] [PubMed] [Google Scholar]
- 19.Ren X, Tuo Y, Li M, et al. Effects of Environmental Factors on Biofilm Formation by Lactobacillus pentosus. Trans Chine Soc Agr Machinery. 2014;45:230. [Google Scholar]
- 20.Li CC, Jiang CY, Jing HJ, et al. Separation of phenolics from peony flowers and their inhibitory activities and action mechanism on bacterial biofilm. Appl Microbiol Biotechnol. 2020;104:4321–4332. doi: 10.1007/s00253-020-10540-z [DOI] [PubMed] [Google Scholar]
- 21.Shi XM, Zhu XN. Biofilm formation and food safety in food industries. Trends Food Sci Tech. 2009;20:407–413. doi: 10.1016/j.tifs.2009.01.054 [DOI] [Google Scholar]
- 22.Toushik SH, Park J-H, Kim K, et al. Antibiofilm efficacy of Leuconostoc mesenteroides J.27-derived postbiotic and food-grade essential oils against Vibrio parahaemolyticus, Pseudomonas aeruginosa, and Escherichia coli alone and in combination, and their application as a green preservative in the seafood industry. Food Res Int. 2022;156:111163. doi: 10.1016/j.foodres.2022.111163 [DOI] [PubMed] [Google Scholar]
- 23.Jadhav S, Shah R, Bhave M, et al. Inhibitory activity of yarrow essential oil on Listeria planktonic cells and biofilms. Food Control. 2013;29:125–130. doi: 10.1016/j.foodcont.2012.05.071 [DOI] [Google Scholar]
- 24.Yang YB, Chen JQ, Zhao YL, et al. Sub-MICs of Azithromycin Decrease Biofilm Formation of Streptococcus suis and Increase Capsular Polysaccharide Content of S. suis. Front Microbiol. 2016;7. doi: 10.3389/fmicb.2016.01659 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Quave CL, Plano LR, Pantuso T, et al. Effects of extracts from Italian medicinal plants on planktonic growth, biofilm formation and adherence of methicillin-resistant Staphylococcus aureus. J Ethnopharmacol. 2008;118:418–428. doi: 10.1016/j.jep.2008.05.005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Limsuwan S, Voravuthikunchai SP. Boesenbergia pandurata (Roxb.) Schltr., Eleutherine americana Merr. and Rhodomyrtus tomentosa (Aiton) Hassk. as antibiofilm producing and antiquorum sensing in Streptococcus pyogenes. FEMS Immunol Med Microbiol. 2008;53:429–436. doi: 10.1111/j.1574-695X.2008.00445.x [DOI] [PubMed] [Google Scholar]
- 27.Taganna JC, Quanico JP, Perono R, et al. Tannin-rich fraction from Terminalia catappa inhibits quorum sensing (QS) in Chromobacterium violaceum and the QS-controlled biofilm maturation and LasA staphylolytic activity in Pseudomonas aeruginosa. J Ethnopharmacol. 2011;134:865–871. doi: 10.1016/j.jep.2011.01.028 [DOI] [PubMed] [Google Scholar]
- 28.Lopes SC, Blanco YC, Justo GZ, et al. Violacein extracted from Chromobacterium violaceum inhibits Plasmodium growth in vitro and in vivo. Antimicrob Agents Chemother. 2009;53:2149–2152. doi: 10.1128/AAC.00693-08 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Huston AL, Methe B, Deming JW. Purification, characterization, and sequencing of an extracellular cold-active aminopeptidase produced by marine psychrophile Colwellia psychrerythraea strain 34H. Appl Environ Microbiol. 2004;70:3321–3328. doi: 10.1128/AEM.70.6.3321-3328.2004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Navarini L, Gilli R, Gombac V, et al. Polysaccharides from hot water extracts of roasted Coffea arabica beans: isolation and characterization. Carbohydr Polym. 1999;40:71–81. doi: 10.1016/S0144-8617(99)00032-6 [DOI] [Google Scholar]
- 31.Zhang XS, Garcia-Contreras R, Wood TK. YcfR (BhsA) influences Escherichia coli biofilm formation through stress response and surface hydrophobicity. J Bacteriol. 2007;189:3051–3062. doi: 10.1128/JB.01832-06 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Lin L, Wang XL, Cui HY. Synergistic efficacy of pulsed magnetic fields and Litseacubeba essential oil treatment against Escherichia coli O157:H7 in vegetable juices. Food Control. 2019;106. [Google Scholar]
- 33.Nithya C, Aravindraja C, Pandian SK. Bacillus pumilus of Palk Bay origin inhibits quorum-sensing-mediated virulence factors in Gram-negative bacteria. Res Microbiol. 2010;161:293–304. doi: 10.1016/j.resmic.2010.03.002 [DOI] [PubMed] [Google Scholar]
- 34.Cottiglia F, Loy G, Garau D, et al. Antimicrobial evaluation of coumarins and flavonoids from the stems of Daphne gnidium L. Phytomedicine. 2001;8:302–305. doi: 10.1078/0944-7113-00036 [DOI] [PubMed] [Google Scholar]
- 35.Lima VN, Oliveira-Tintino CDM, Santos ES, et al. Antimicrobial and enhancement of the antibiotic activity by phenolic compounds: gallic acid, caffeic acid and pyrogallol. Microb Pathogenesis. 2016;99:56–61. doi: 10.1016/j.micpath.2016.08.004 [DOI] [PubMed] [Google Scholar]
- 36.Liu MH, Wu XX, Li JK, et al. The specific anti-biofilm effect of gallic acid on Staphylococcus aureus by regulating the expression of the ica operon. Food Control. 2017;73:613–618. doi: 10.1016/j.foodcont.2016.09.015 [DOI] [Google Scholar]
- 37.Sheng LN, Rasco B, Zhu MJ. Cinnamon Oil Inhibits Shiga Toxin Type 2 Phage Induction and Shiga Toxin Type 2 Production in Escherichia coli O157:H7. Appl Environ Microb. 2016;82:6531–6540. doi: 10.1128/AEM.01702-16 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Laspidou CS, Rittmann BE. A unified theory for extracellular polymeric substances, soluble microbial products, and active and inert biomass. Water Res. 2002;36:2711–2720. doi: 10.1016/S0043-1354(01)00413-4 [DOI] [PubMed] [Google Scholar]
- 39.Nagar N, Aswathanarayan JB, Vittal RR. Anti-quorum sensing and biofilm inhibitory activity of Apium graveolens L. oleoresin. J Food Sci Tech Mys. 2020;57:2414–2422. doi: 10.1007/s13197-020-04275-y [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Papenfort K, Bassler BL. Quorum sensing signal–response systems in Gram-negative bacteria. Nat Rev Microbiol. 2016;14:576–588. doi: 10.1038/nrmicro.2016.89 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Waters CM, Bassler BL. QUORUM SENSING: cell-to-Cell Communication in Bacteria. Annu Rev Cell Dev Biol. 2005;21:319–346. doi: 10.1146/annurev.cellbio.21.012704.131001 [DOI] [PubMed] [Google Scholar]
- 42.Wang JF, Liu QJ, Li XH, et al. In-situ monitoring AHL-mediated quorum-sensing regulation of the initial phase of wastewater biofilm formation. Environ Int. 2020;2:135. [DOI] [PubMed] [Google Scholar]
- 43.Whiteley M, Diggle SP, Greenberg EP. Progress in and promise of bacterial quorum sensing research. Nature. 2017;551:313–320. doi: 10.1038/nature24624 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.De Kievit TR, Gillis R, Marx S, et al. Quorum-sensing genes in Pseudomonas aeruginosa biofilms: their role and expression patterns. Appl Environ Microb. 2001;67:1865–1873. doi: 10.1128/AEM.67.4.1865-1873.2001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Yin HG, Deng YF, Wang HF, et al. Tea polyphenols as an antivirulence compound Disrupt Quorum-Sensing Regulated Pathogenicity of Pseudomonas aeruginosa. Sci Rep-UK. 2015;5:6836. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Choo JH, Rukayadi Y, Hwang JK. Inhibition of bacterial quorum sensing by vanilla extract. Lett Appl Microbiol. 2006;42:637–641. doi: 10.1111/j.1472-765X.2006.01928.x [DOI] [PubMed] [Google Scholar]
- 47.Vikram A, Jayaprakasha GK, Jesudhasan PR, et al. Suppression of bacterial cell-cell signalling, biofilm formation and type III secretion system by citrus flavonoids. J Appl Microbiol. 2010;109:515–527. doi: 10.1111/j.1365-2672.2010.04677.x [DOI] [PubMed] [Google Scholar]
- 48.Jaisi DP, Dong HL, Kim J, et al. Nontronite particle aggregation induced by microbial Fe(III) reduction and exopolysaccharide production. Clay Clay Miner. 2007;55:96–107. doi: 10.1346/CCMN.2007.0550108 [DOI] [Google Scholar]
- 49.LewisOscar F, Nithya C, Bakkiyaraj D, et al. Biofilm Inhibitory Effect of Spirulina platensis Extracts on Bacteria of Clinical Significance. Proce National Acad Sci. 2015;87:537–544. doi: 10.1007/s40011-015-0623-9 [DOI] [Google Scholar]
- 50.Nithyanand P, Thenmozhi R, Rathna J, et al. Inhibition of Streptococcus pyogenes Biofilm Formation by Coral-Associated Actinomycetes. Curr Microbiol. 2010;60:454–460. doi: 10.1007/s00284-009-9564-y [DOI] [PubMed] [Google Scholar]
- 51.Polaquini SRB, Svidzinski TIE, Kemmelmeier C, et al. Effect of aqueous extract from Neem (Azadirachta indica A. Juss) on hydrophobicity, biofilm formation and adhesion in composite resin by Candida albicans. Arch Oral Biol. 2006;51:482–490. doi: 10.1016/j.archoralbio.2005.11.007 [DOI] [PubMed] [Google Scholar]
- 52.Oh YJ, Lee NR, Jo W, et al. Effects of substrates on biofilm formation observed by atomic force microscopy. Ultramicroscopy. 2009;109:874–880. doi: 10.1016/j.ultramic.2009.03.042 [DOI] [PubMed] [Google Scholar]
- 53.Bonez PC, Rossi GG, Bandeira JR, et al. Anti-biofilm activity of A22 ((S-3,4-dichlorobenzyl) isothiourea hydrochloride) against Pseudomonas aeruginosa: influence on biofilm formation, motility and bioadhesion. Microb Pathogenesis. 2017;111:6–13. doi: 10.1016/j.micpath.2017.08.008 [DOI] [PubMed] [Google Scholar]