
Figure 2. The flavonol biosynthetic machinery is localized to the root epidermis to regulate root hair initiation.
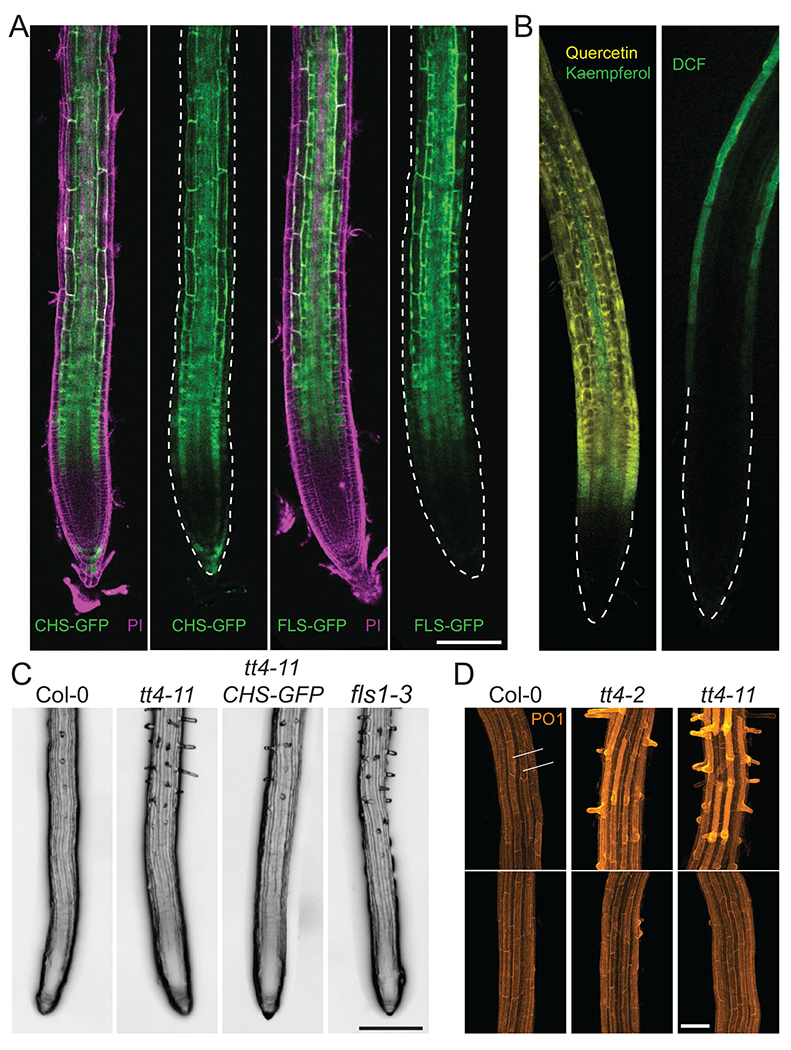
(a) Fluorescence of transgenic Arabidopsis lines containing CHS-GFP and FLS1-GFP reporters (green) stained with the cell wall probe propidium iodide (PI) (magenta) show that flavonol biosynthetic enzymes localize to the root epidermis. Images without PI channel have epidermal tissues outlined by dashed lines. Scale bar =100 μm (b) LSCM images of WT Arabidopsis roots treated with either the flavonol-selective probe, DPBA, with yellow fluorescence of quercetin and green fluorescence of kaempferol (left image), or the fluorescence of the general ROS sensor, dichlorofluorescein (DCF) (green, right image), reveals ROS accumulate in higher levels in root epidermal tissues where flavonols are less abundant. (c) Representative images of root hair number in WT Arabidopsis and mutants with impaired flavonol accumulation, tt4-ll and fls1-3, and tt4-11 complemented with a CHS-GFP transgene (ttd-11 CHS-GFP) show increased root hair numbers in mutants with defects in flavonol synthesis. Scale bar = 200 μm (d) Confocal images of WT, tt4-2, and tt4-11 Arabidopsis lines stained with the H2O2 probe, Peroxy orange 1, display elevated H2O2 accumulation in root hair forming cells (denoted as 1, 3, and 5) relative to nonhair cells (2 and 4). H2O2 accumulation is increased in tt4-2 and tt4-11, though this phenotype can be restored to WT levels through treatment with the flavonol precursor, naringenin. Scale bar = 50 μm. Adapted from Ref. [64].