
Fig. 6. A model for LR asymmetry in conjoined twins.
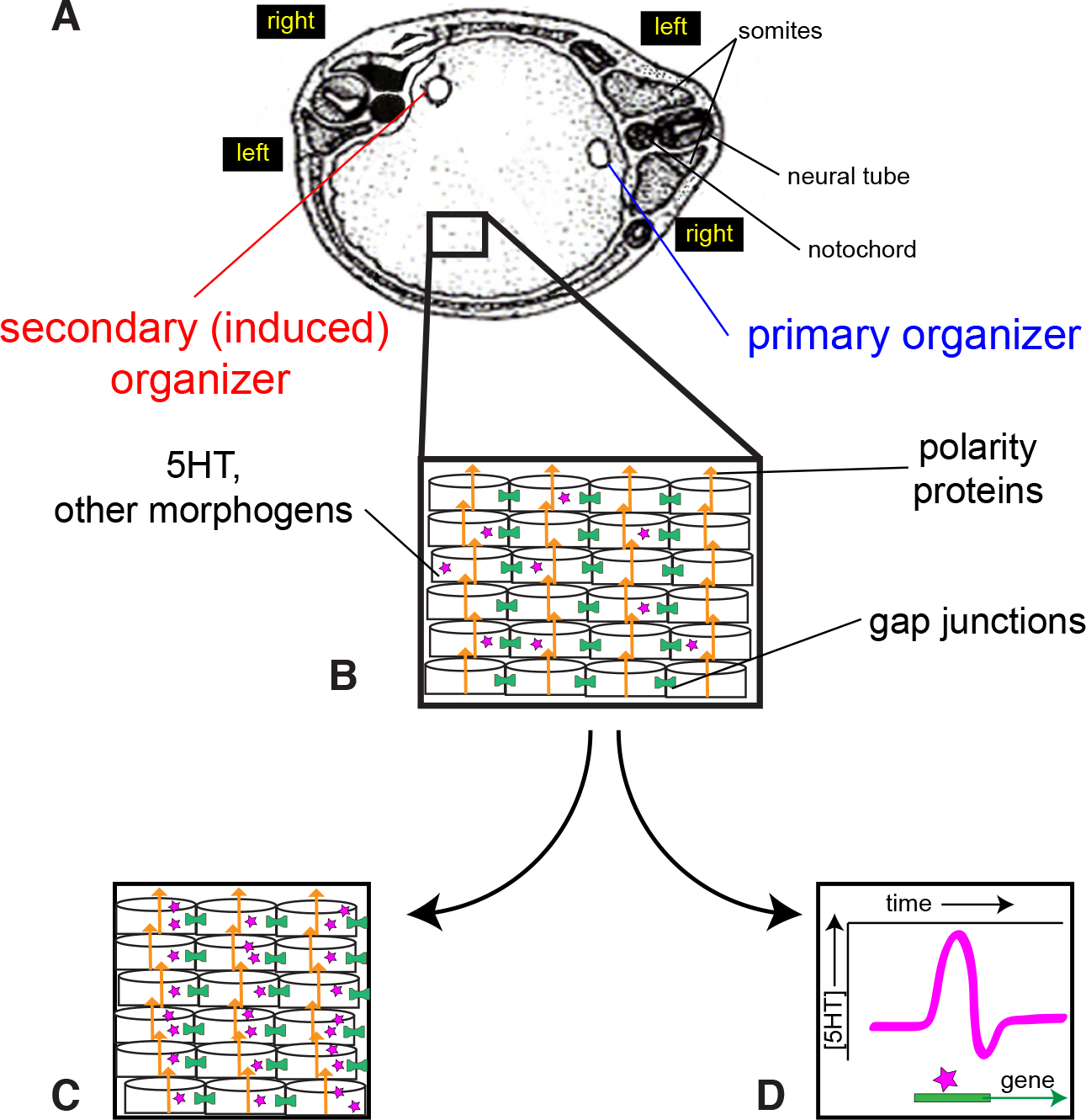
(A) The two organizers are positioned in a single blastoderm. The left and right sides of each twin are indicated. Note that the left side of the primary twin is across from the right side of the induced twin. (B) The cells of this blastoderm are oriented by polarity proteins (orange arrows) including planar cell polarity and apical basal polarity proteins. These polarity proteins are themselves oriented with respect to the anterior-posterior and dorsal-ventral axes, as described previously (Vandenberg and Levin, 2012). Within this highly organized blastoderm, both gap junctions (green cylinders) and small signaling molecules like 5-HT (purple stars) are required for proper LR patterning in the induced twin. Two roles for 5-HT are proposed. (C) One hypothesis is that 5-HT maintains subcellular asymmetries, providing an amplification of the asymmetries established by the polarity proteins. Thus, each cell in the blastoderm, or perhaps only specific cells in a late LR “organizer”, has its own physiological and biochemical LR axis. (D) The second hypothesis is that a surge of 5-HT at a specific time, and perhaps again only in specific cells in a late LR “organizer”, are sufficient to induce consistent LR gene expression. Thus, asymmetric expression is not required, but a temporal pulse is needed.