
Fig. 5. An intact brain is critical for driving cilia reorientation.
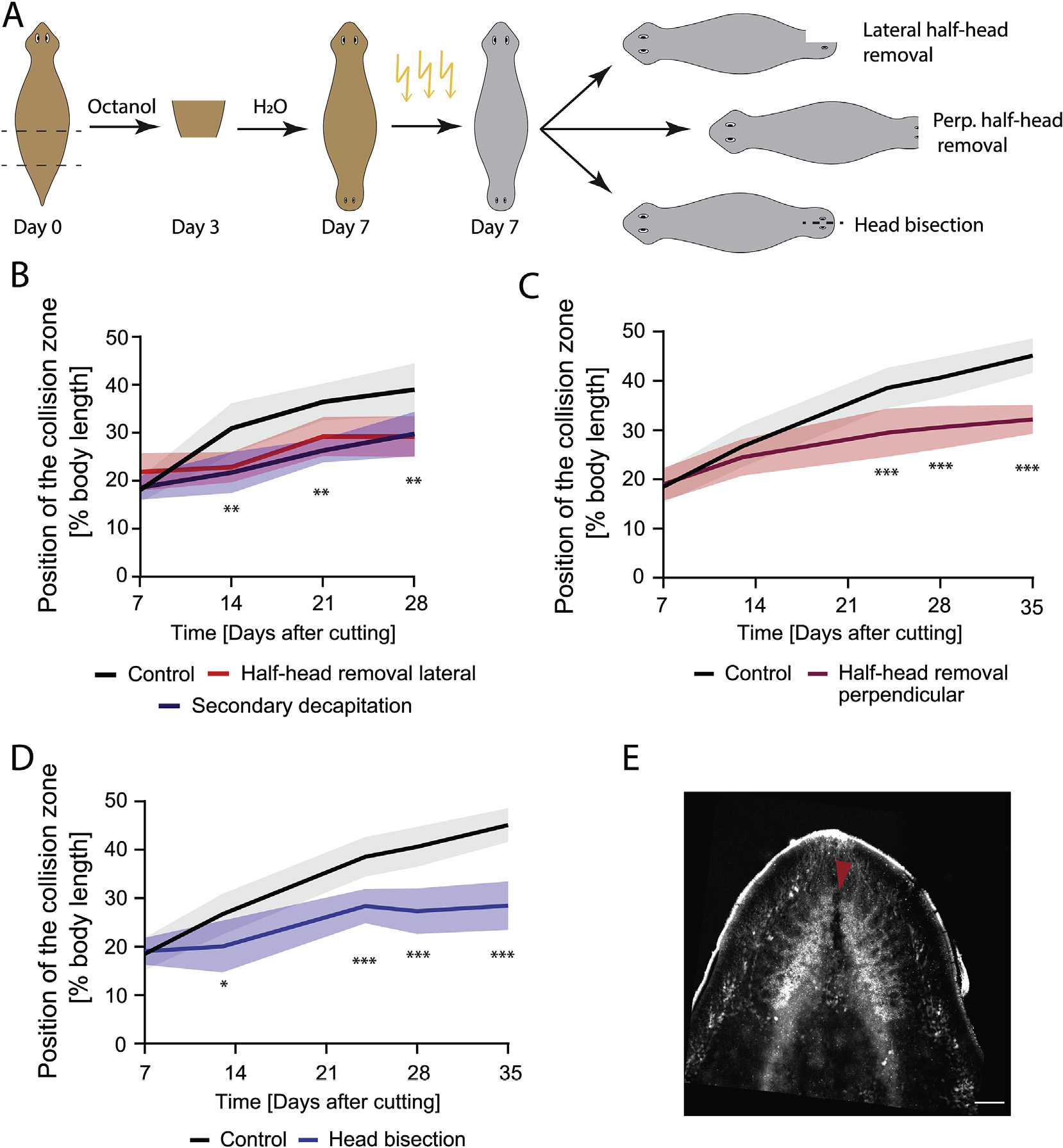
A) Experimental scheme showing induction of the DHs via Octanol treatment, followed by irradiation to remove neoblasts at Day 7. Cuts to remove parts of the brain or bisect the head were performed several hours after irradiation. B) DHs with the lateral half of the secondary head removed following irradiation at Day 7 (red) showed significantly reduced cilia reorientation speed compared to control (black) to the same extent as animals with the entire secondary head removed (blue). C) DHs with the frontal half of the secondary head removed following irradiation at Day 7 (red) showed significantly reduced cilia reorientation speed compared to control (black). D) DHs with secondary head bisected laterally between the eyes without tissue removal following irradiation at Day 7 (blue) reoriented their cilia significantly slower compared to controls (black). E) Synapsin stain of brain following head bisection without tissue removal. Red arrow highlighting disruption of the brain due to the head bisecting cut. *p < 0.01, **p < 0.001, ***p < 0.0001. N = 12. Plotted is mean of all samples, with standard deviation as shaded area for all experiments with their age-matched controls. Scale bar 100 μm.