
Figure 3. IFNγ-producing Tfh cells are required for influenza-specific lung-BRM responses.
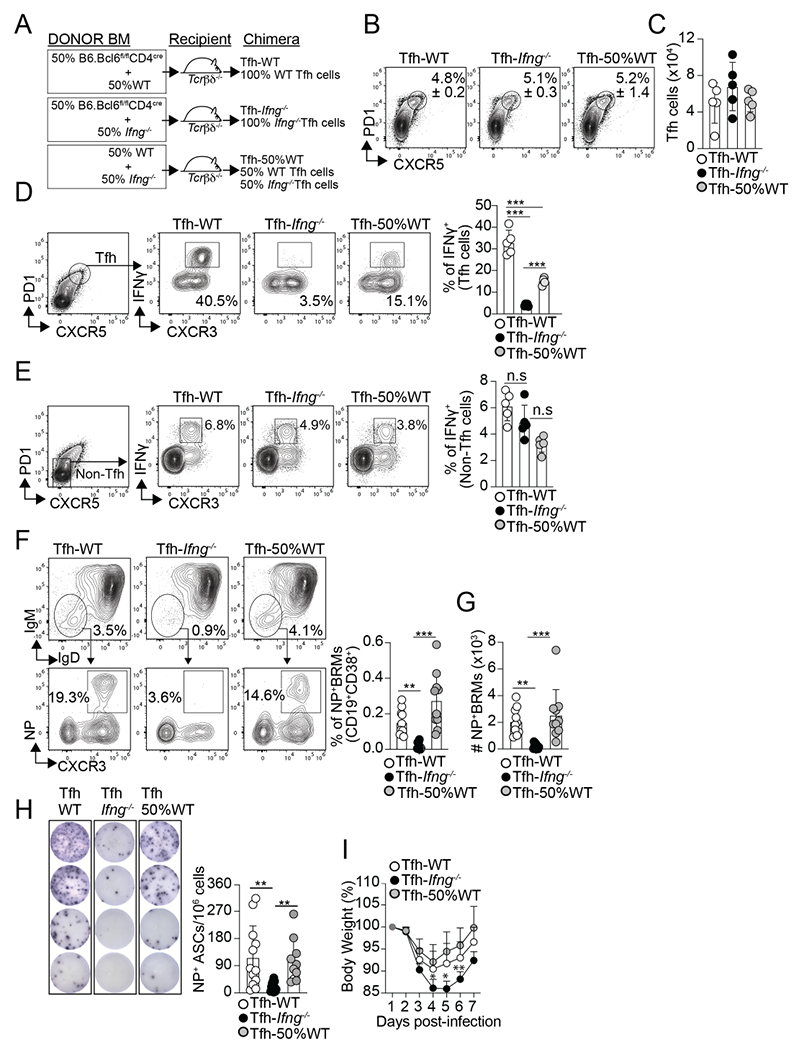
(A) Diagram showing the Tfh-WT, Tfh- Ifng−/−, and Tfh-50%WT BM chimeras design. Frequency (B) and number (C) of Tfh cells in the med-LN on day 12 after PR8 infection. Frequency of IFNγ+ cells within the Tfh cells (D) and non-Tfh cells (E) on day 12 after infection. Data are representative of three independent experiments (n=5 mice/group). Frequency (F) and number (G) of class-switched NP-specific BRMs in the lungs on day 60 after infection. Data were pooled from two independent experiments (n=10 mice/group). Data are representative of four independent experiments. (H and I) The chimeric mice were infected with PR8 and challenged with X31 on day 30. (H) IgG NP-specific ELISPOTs in the lungs on day 6 after rechallenge. Data were pooled from two independent experiments (n=10-12 mice/group). (I) Body weight loss after rechallenge (n=5-6 mice). Data are representative of two independent experiments. All P values were determined by one-way ANOVA with a post- hoc Kruskal–Wallis comparison test. Data shown as the mean ± SD. *p < 0.05, **p < 0.01, ***p < 0.001; ns, not significant.