

Abstract
To date, 11 thermosensitive transient receptor potential (thermo-TRP) channels have been identified. Recent studies have characterized the mechanism of thermosensing by thermo-TRPs and the physiological role of thermo-TRPs in energy metabolism. In this review, we highlight the role of various thermo-TRPs in energy metabolism and hormone secretion. In the pancreas, TRPM2 and other TRPs regulate insulin secretion. TRPV2 expressed in brown adipocytes contributes to differentiation and/or thermogenesis. Sensory nerves that express TRPV1 promote increased energy expenditure by activating sympathetic nerves and adrenaline secretion. Here, we first show that capsaicin-induced adrenaline secretion is completely impaired in TRPV1 knockout mice. The thermogenic effects of TRPV1 agonists are attributable to brown adipose tissue (BAT) activation in mice and humans. Moreover, TRPA1- and TRPM8-expressing sensory nerves also contribute to potentiation of BAT thermogenesis and energy expenditure in mice. Together, thermo-TRPs are promising targets for combating obesity and metabolic disorders.
Keywords: TRP channel, Insulin, Brown adipose adipocyte, UCP1, Adrenaline, Energy expenditure
Introduction
Most transient receptor potential (TRP) channels are non-selective cation channels. The name TRP comes from the prototypical member in Drosophila, in which a mutation resulted in abnormal transient receptor potential to continuous light [1]. TRP channels are now divided into seven subfamilies: TRPC (canonical), TRPV (vanilloid), TRPM (melastatin), TRPML (mucolipin), TRPP (polycystin), TRPA (ankyrin) and TPRN (NomPC). In mammals, there are six TRP subfamilies and 28 channels. TRP channels are expressed in many tissues and have a wide variety of physiological functions, including detection of various physical and chemical stimuli in vision, taste, olfaction, hearing, touch, and thermosensation [2, 3]. The gene encoding the capsaicin receptor as a noxious heat sensor, which is now called TRPV1, was isolated from a rodent sensory neuron cDNA library in 1997 and was considered to be a breakthrough for research concerning temperature sensing [4]. Since then, several TRP channels having thermosensitive abilities have been identified in mammals, with 11 thermosensitive TRP (thermo-TRP) channels reported in mammals to date (Table 1). These channels belong to the TRPV, TRPM, TRPA, and TRPC subfamilies, and their temperature thresholds for activation are in the range of physiological temperatures, which we can discriminate. TRPV1 and TRPV2 are activated by elevated temperatures, whereas TRPM8 and TRPA1 are activated by cool and cold temperatures. TRPV3, TRPV4, TRPM2, TRPM4, and TRPM5 are activated by warm temperatures. In addition, TRPM3 was shown to be a sensor for noxious heat and TRPC5 was identified as a candidate cold sensor [5, 6]. Thermo-TRP channels usually function as ‘multimodal receptors’ that respond to various chemical and physical stimuli. For example, TRPV1, activated by noxious heat (>42 °C), is also a receptor for several pungent agents such as capsaicin, an active ingredient in chili peppers, as well as by low pH. Activation of these channels could contribute to changes in intracellular Ca2+ concentrations ([Ca2+]i) and control of membrane potentials in many cell types, except TRPM4 and TRPM5, which are not permeable of divalent cations. Thermo-TRP channels expressed in sensory neurons and skin can act as ambient temperature sensors. On the other hand, thermo-TRP channels are also expressed in tissues that are not exposed to dynamic temperature changes, suggesting that these channels have other physiological roles that are unrelated to sensation of temperature changes.
Table 1.
Properties of thermosensitive TRP channels
| Temperature threshold | Tissue distribution | Other stimuli | ||
|---|---|---|---|---|
| Heat | TRPV1 | >42 °C | Sensory neuron, brain, skin | Capsaicin, proton, capsiate, gingerol, shogaol, allicin, shanshool, camphor, resiniferatoxin, vanillotoxin, 2-APB, propofol, anandamide, arachidonic acid metabolic products (by lipoxygenases), monoacylglycerol, NO, extracellular cation |
| TRPV2 | >52 °C | Sensory neuron, brain, spinal cord, lung, liver, spleen, colon, heart, immunocyte | Probenecid, 2-APB, cannabidiol, mechanical stimulation | |
| Warm | TRPV3 | >32 °C | Skin, sensory neuron, brain, spinal cord, stomach, colon | Camphor, carvacrol, menthol, eugenol, thymol, 2-APB |
| TRPV4 | >27–41 °C | Skin, sensory neuron, brain, kidney, lung, inner ear, bladder | 4α-PDD, bisandrographolide, citric acid, arachidonic acid metabolic products (by epoxygenases), anandamide, hypoosmolality, mechanical stimulation | |
| TRPM2 | >36 °C | Brain, immunocyte, pancreas etc. | (cyclic) ADPribose, β-NAD, H2O2, intracellular Ca2+ | |
| TRPM3 | Warm-heat | Brain, sensory neuron, pancreas, eye | Ca2+ store depletion, pregnenolone sulfate, nifedipine, clotrimazole | |
| TRPM4 | Warm | Heart, liver, immunocyte, pancreas etc. | Intracellular Ca2+ | |
| TRPM5 | Warm | Taste cell, pancreas | Intracellular Ca2+ | |
| Cold | TRPM8 | <27 °C | Sensory neuron | Menthol, icilin, eucalyptol |
| TRPC5 | Cold | Brain, sensory neuron, liver, heart, kidney | Gq/11-coupled receptors, diacylglycerol, Gd3+ | |
| TRPA1 | <17 °C | Sensory neuron, inner cell | Allyl isothiocyanate, carvacrol, cinnamaldehyde, allicin, dially trisulfide, miogadial, miogatrial, capsiate, acrolein, icilin, tetrahydrocannabinol, menthol (10-100 μM), formalin, H2O2, alkalization, intracellular Ca2+, NSAIDs, propofol/isoflurane/desflurane/etomidate/octanol/hexanol etc. |
2-APB 2-aminoethoxydiphenyl borate, NO nitric oxide, 4α-PDD 4α-phorbol-didecanoate, ADPribose adenosine diphosphate ribose, β-NAD β-nicotinamide adenine dinucleotide, H 2 O 2 hydrogen peroxide, NSAIDs non-steroidal anti-inflammatory drugs
The prevalence of excess weight and obesity is increasing at an alarming rate worldwide. According to the World Health Organization, in 2014, ~39% (1.9 billion) and 13% (600 million) of adults were overweight and obese, respectively [7]. These numbers are expected to increase in the future [8]. Obesity is a major risk factor for metabolic syndromes, including type 2 diabetes, as well as for cardiovascular and cerebral diseases. The fundamental cause of excess weight and obesity is an energy imbalance between energy intake and energy expenditure. Recent studies showed that several thermo-TRP channels are key molecules in the regulation of energy metabolism. In this review, we focus on the involvement of thermo-TRP channels, especially those expressed in the pancreas, brown adipocytes and sensory nerves, in energy metabolism and the secretion of the metabolically important hormones insulin and adrenaline.
Thermo-TRP in the pancreas and regulation of insulin secretion
TRPM2 channel in β-cells
In pancreatic β-cells, glucose metabolism-induced closure of ATP-sensitive K+ (KATP) channels and membrane depolarization trigger opening of voltage-dependent Ca2+ channels, which induces Ca2+ influx and subsequent insulin secretion. During glucose-stimulated insulin secretion in β-cells, induction of background inward current promoted by the opening of non-selective cation channels (NSCCs) might facilitate depolarization after glucose metabolism-induced closure of the KATP channels. Glucose metabolism evokes not only KATP channel inhibition but also increases NSCC currents. We reported that the NSCC transient receptor potential melastatin 2 (TRPM2) channel in β-cells plays an essential role in glucose-induced and incretin-potentiated insulin secretion [9]. TRPM2 is expressed in β-cells [10] and the increase in glucose-induced NSCC activity is due to opening of TRPM2 channels, since this glucose effect was attenuated in β-cells from TRPM2-deficient mice [11]. These effects of glucose on KATP channel inhibition and NSCC (TRPM2) activation may synergistically and effectively depolarize the β-cell membrane to trigger acute insulin secretion. This mechanism may contribute to the priming of insulin release from β-cells, since glucose-induced TRPM2 activation occurs before glucose-induced KATP channel inhibition [11].
Intestinal incretin hormones, such as glucagon-like peptide-1 (GLP-1) secreted from L-cells and glucose-dependent insulinotropic polypeptide (GIP) secreted from K-cells after meal, potentiate glucose-induced insulin release by cytosolic cAMP production via Gs-coupled receptors [12, 13]. GLP-1 activates NSCC currents and depolarizes the membrane potential through cAMP production in β-cells. In wild-type mice, the non-selective TRPM2 blocker 2-aminoethoxydiphenyl borate (2-APB) inhibits GLP-1-mediated increases in NSCC currents. Furthermore, GLP-1 has no effect on insulin secretion in TRPM2-deficient mice [11, 14]. These results demonstrate that TRPM2 activation is an important pathway for incretin-potentiated insulin secretion.
Considering the mechanism of TRPM2 channel stimulation mediated by cAMP signaling via Gs-coupled receptors, Gi/Go-mediated inhibition of cAMP production is expected to attenuate TRPM2 channel activity. Ghrelin, an acylated 28-amino acid peptide produced predominantly in the stomach, was discovered as the endogenous ligand for the growth hormone secretagogue-receptor (GHS-R), which is widely expressed throughout the body. Ghrelin inhibits glucose-stimulated insulin secretion in vitro in perfused pancreas tissue and isolated islets [15, 16]. We found that the insulinostatic action of ghrelin is produced via pertussis toxin-sensitive Gi-proteins in β-cells that in turn attenuate cAMP and [Ca2+]i signaling in β-cells and insulin release from islets [17, 18]. Moreover, ghrelin markedly counteracts glucose (8.3 mM)-induced activation of TRPM2 currents in islet β-cells from wild-type mice but not TRPM2 knockout (TRPM2-KO) mice [19]. These results suggest that ghrelin suppresses glucose-induced insulin secretion at least partly by inhibiting TRPM2 channels. Furthermore, ghrelin potently attenuates GLP-1-induced cAMP generation and insulin release from islet β-cells, whereas ghrelin receptor antagonists potentiate GLP-1-induced cAMP generation and insulin release [20]. Consistent with ghrelin signaling, we recently found that adrenaline attenuates TRPM2 activation via Gi-mediated inhibition of cAMP signaling in β-cells [21].
The gastric hormones GLP-1 and ghrelin have reciprocal actions on cAMP levels and TRPM2 channel activity in islet β-cells. GLP-1 and ghrelin are released in a reciprocal pattern: following a meal, the GLP-1 plasma level rises while ghrelin levels fall [22]. These changes may collaborate to effectively elevate cAMP and activate TRPM2 channels in β-cells, leading to rapid and efficient insulin release for regulated postprandial glucose disposal. As such, the development of approaches that would specifically intervene in TRPM2 signaling in β-cells might provide a potential therapeutic tool to treat patients with type 2 diabetes.
Other TRP channels in β-cells
Several TRP family members are reportedly expressed in β-cells, although their physiological role is unclear and mechanistic insights into the regulation of these channels during insulin release are limited. TRPM3 expressed in β-cells functions as an ionotropic steroid receptor that links insulin release [23], although the inhibition of TRPM3 channels does not affect glucose-induced insulin secretion [24]. TRPM4 is a Ca2+-activated NSCC that may play a key role in controlling the membrane potential and electrical activity of insulin-secreting INS1 cells [25]. TRPM5 is also activated by Ca2+ and plays a role in insulin release [26]. GLP-1 stimulates insulin secretion in part by promoting TRPM4 and TRPM5 activation [27]. Activation of TRPA1 channels expressed in β-cells by the agonist allyl isothiocyanate stimulates insulin release from insulinoma cells and primary β-cells [28]. TRPA1-mediated depolarization may act synergistically with KATP channel blockade to facilitate insulin release in β-cells. Interestingly, the anti-diabetic drug glibenclamide inhibits KATP channels but activates human TRPA1 channels [29], suggesting that part of the insulin-secreting action of glibenclamide could be attributed to TRPA1 activation.
Roles of thermo-TRP channels in brown adipose tissue
Trpv2
TRPV2 was initially reported to be activated by noxious heat (temperature threshold >52 °C) [30]. TRPV2 was also found to act as a mechano-sensor that is activated by membrane stretch and cell swelling [31]. 2-APB and lysophosphatidylcholine (LPC) act as TRPV2 agonists [32, 33], whereas ruthenium red and SKF96365 antagonize TRPV2 activity [32], although these ligands are not specific for TRPV2. TRPV2 is dominantly expressed in the central and peripheral nervous systems and is involved in axon outgrowth in developing neurons and intestinal movement [34–36]. In addition, the phenotype of TRPV2 knockout (TRPV2-KO) mice suggests that TRPV2 is involved in phagocytosis of macrophages [37].
TRPV2 was recently shown to be expressed in brown adipose tissue (BAT) and primary brown adipocytes. In particular, the increase in TRPV2 expression levels during brown adipocyte differentiation suggests that TRPV2 could have important roles in differentiated brown adipocytes. A study by Sun et al. showed that TRPV2-KO mice have alterations in the mRNA levels for genes that are related to mitochondrial oxidative metabolism [38]. For instance, mice lacking TRPV2 have decreased amounts of mRNA of UCP1, which is a mitochondrial transporter protein and plays important roles in energy balance and regulation, and peroxisome proliferator-activated receptor gamma coactivator 1-alpha (PERM1) expression that are accompanied by increases in expression of genes related to lipid accumulation, such as lipoprotein lipase (LPL), and cluster of differentiation 36 (CD36). Furthermore, in morphological terms, TRPV2-KO BAT cells were larger overall and had larger lipid droplets compared to wild-type BAT. These differences could be explained by the impaired thermogenic activity seen for TRPV2-KO BAT. TRPV2-KO mice are unable to maintain body temperature upon exposure to cold, but these mice had locomotor activity and sympathetic nerve activity that were similar to wild-type mice. In addition, increases in UCP1 mRNA and protein in BAT were impaired in TRPV2-KO mice exposed to cold temperature (4 °C). TRPV2 expression levels in BAT were also increased following exposure of wild-type mice to cold. Thermogenesis in mice can be assessed by measuring interscapular BAT (iBAT) temperature using an inserted temperature probe. The iBAT temperature could be increased by systemic administration of the β3-adrenergic receptor agonist BRL37344 to anesthetized wild-type mice, but not TRPV2-KO mice. Consistent with this impairment, TRPV2-KO mice showed heavier white adipose tissue (WAT) and increased accumulation of lipid droplets in BAT. Moreover, TRPV2-KO mice fed a high-fat diet had a significant increases in body weight and metabolically active tissues.
These findings raise several questions: (1) how is TRPV2 activated downstream of sympathetic nerve activation? (2) what are the mechanisms involved in increased TRPV2 expression? and (3) what is the involvement of calcium signaling in BAT thermogenesis? Stimuli that activate TRPV2 include membrane stretch (mechanical tension), as well as the chemical agonists LPC, LPI, and the endocannabinoids [31, 33, 39, 40]. Insulin growth factor-1 (IGF-1) is reported to enhance translocation of TRPV2 from intracellular compartments to the plasma membrane following stretch-mediated activation of TRPV2 [41]. In brown adipocytes, TRPV2 could be activated downstream of adenylyl cyclase (AC), based on the finding that enhancement of UCP1 mRNA expression mediated by the AC activator forskolin was almost abolished in primary brown adipocytes from TRPV2-KO mice [38]. Although the precise mechanism of TRPV2 activation in BAT is unclear, several studies indicated that TRPV2 agonists and stimulations could work synergistically both in vitro and in vivo. There is little evidence for the dependence of calcium influx on sympathetic nerve activation of brown adipocytes, although increases in intracellular calcium concentrations have been observed in these cells. Calcium is thought to be released from mitochondria or the ER followed by store-operated calcium entry. On the other hand, activation of TRPM8 by menthol in brown adipocytes reportedly enhances calcium-dependent PKA phosphorylation (also described below). This result suggests that a calcium influx pathway that promotes entry of extracellular calcium directly into the cell may exist, and TRPV2 could be a candidate channel that promotes this entry. Another important event in brown adipocyte function is PKA phosphorylation. Given that chelation of intracellular calcium by BAPTA-AM attenuates increases in UCP1 expression in brown adipocytes treated with the β-adrenergic receptor agonist isoproterenol [38], it is possible that calcium influx could modulate UCP1 expression increase through PKA phosphorylation. Furthermore, TRPV2 activation could enhance PKA phosphorylation levels [42], although further experiments are necessary to clarify the precise mechanisms of thermogenesis mediated by TRPV2 activation.
TRPV2 mRNA expression can also be seen in primary pre-adipocytes, but its expression is significantly increased in differentiated brown adipocytes compared to pre-adipocytes [43]. Pre-adipocytes isolated from mouse iBAT showed impaired differentiation to brown adipocytes in the presence of non-selective TRPV2 agonists (2-APB and LPC). However, application of the TRPV2 agonists 3 days after differentiation began had no effect, indicating that TRPV2 activation could be critical for the early stages of pre-adipocyte differentiation [43]. Mechanical stimulation, which could activate TRPV2, also inhibited brown adipocyte differentiation. In addition, brown adipocyte differentiation was enhanced in TRPV2-KO mice. Calcium influx is reported to suppress brown adipocyte differentiation through a calcineurin-dependent pathway [44]. Indeed, the calcineurin inhibitors cyclosporine A and FK506 partially recovered TRPV2 activation-induced inhibition of brown adipocyte differentiation [43]. These results demonstrated that TRPV2 is involved in the differentiation of brown adipocytes, and could regulate the number of brown adipocytes. Although the phenotype related to impaired differentiation is not seen in TRPV2-KO BAT, a compensatory effect could be present in these mice, particularly given the importance of BAT functions.
Taken together, these findings suggest that TRPV2 plays two different roles related to the developmental stages of brown adipocytes. First, in pre-adipocytes, TRPV2 could inhibit the differentiation of brown adipocytes, and second, in differentiated brown adipocytes TRPV2 could facilitate thermogenesis.
Other TRP channels
TRPV1 is reported to be expressed in the 3T3-L1 and HB2 adipocyte cell lines, brown adipocytes, and BAT [45–47]. Application of the TRPV1 agonist (capsaicin) to 3T3-L1 adipocytes caused upregulation of the expression of genes related to thermogenesis and “browning” [46]. However, the physiological roles of TRPV1 in BAT have not been well clarified.
TRPM8 is also expressed in BAT and menthol-induced activation of TRPM8 expressed in brown adipocytes upregulates UCP1 expression, which requires the activation of PKA [48]. Chronic dietary application of menthol significantly increases the core body temperature and locomotor activity in wild-type mice, whereas these effects are absent in both TRPM8-KO and UCP1-KO mice [48]. In addition, TRPM8 was demonstrated to be expressed in a human white adipocyte cell line and its expression level is elevated during the differentiation of adipocyte. TRPM8 activation induced UCP1 expression, mitochondrial activation and heat production [49]. Although this study suggests that TRPM8 stimulation enhances non-shivering thermogenesis, the roles of TRPM8 in BAT are still largely unclear [48].
TRPV4 is expressed in both BAT and WAT [50]. Reduction of TRPV4 expression enhances the expression of genes related to thermogenesis such as PGC-1α and UCP1 without changing adipogenesis in 3T3-F442A adipocytes [50]. TRPV4 activation causes a rapid phosphorylation of ERK1/2 and JNK1/2, which further suppresses the expression of thermogenic genes [51]. Consistent with this finding, TRPV4-KO mice exhibit increased muscle oxidative capacity and resistance to diet-induced obesity [50]. Another report indicated that TRPV4 is expressed in WAT and BAT [50]. Interestingly, induction of thermogenic gene expression upon TRPV4 inhibition by GSK205 leads to the development of metabolically active brown fat-like features in WAT [50]. Calcium influx through TRPV4 has an opposite effect to that seen for TRPV2 in BAT thermogenesis. Thus, how and when calcium influx occurs during thermogenesis in BAT could have important regulatory implications.
Activation of thermo-TRP channels in sensory nerves increases energy expenditure via sympathetic nerve activation and via enhancement of adrenaline secretion
Findings on the effects and mechanisms of thermo-TRP from animal studies
It is empirically known that consumption of spicy foods or drinks can enhance thermogenesis by increasing energy expenditure. In traditional Chinese medicine, many spicy foods were shown to induce warming sensations in the body. In 1986, Henry and Emery provided evidence to link consumption of hot foods and enhanced energy expenditure, in that individuals who consumed spicy foods containing chili or mustard sauces had an approximately twofold increase in O2 consumption after the meal [52]. Red hot peppers (Capsicum sp.) are a representative spicy food, and its pungent ingredient is capsaicin. Also in 1986, Kawada et al. reported that intraperitoneal injection of capsaicin increased O2 consumption and that the addition of capsaicin to a high-fat diet prevented accumulation of visceral WAT and obesity in rats [53, 54]. The effect of capsaicin on increasing energy expenditure has since been supported by numerous studies involving both humans and animals [55, 56].
After cloning and identification of the capsaicin receptor TRPV1 in 1997 [4], the mechanism of action by which capsaicin and TRPV1 agonists increase energy expenditure and thermogenesis has been determined in greater detail. Many pungent ingredients derived from spices activate TRPV1 [57], including capsaicin in hot peppers [4], piperine and its analog in black pepper [58, 59], and gingerol and shogaol in ginger [60, 61]. Interestingly, some compounds with no or very low pungency have been identified as TRPV1 agonists, such as the capsaicin analog capsiate present in “CH-19 sweet” peppers [62, 63], [10] -shogaol from ginger [61], and 1-monoacylglycerol that has various acyl moieties in wheat, mioga (Zingiber mioga) and onion [64]. TRPV1 agonists with no or low pungency can also have high lipophilicity, which could render these molecules unable to access the termini of trigeminal nerves in the oral cavity that is covered with epithelium [61, 63, 64].
Oral administration of the TRPV1 agonists capsaicin and capsiate increases energy expenditure (O2 consumption) and core body temperature, and these responses are significantly blunted by a TRPV1 antagonist [65] and abolished in TRPV1-KO mice [66], suggesting that TRPV1 is an essential receptor for increasing energy expenditure and heat production. TRPV1 is markedly expressed in peripheral sensory neurons derived from the dorsal root ganglion, the trigeminal ganglion and the nodose ganglion [4, 67, 68]. Denervation of TRPV1-expressing sensory nerves by systemic pretreatment with excess capsaicin can completely block enhancement of O2 consumption and increases in body temperature [69]. Therefore, TRPV1 agonists induce incremental energy expenditure and thermogenesis via TRPV1-expressing sensory nerves.
Adrenaline produced by the adrenal medulla is an important stimulating hormone that increases energy expenditure and thermogenesis, as well as possibly contributing to diet-induced energy expenditure. Meal intake induces increases in plasma adrenaline and O2 consumption, and meal-induced energy expenditures can be blunted by the β-blocker propranolol [70]. Administration of capsaicin enhances adrenal sympathetic nerve activity [71] and adrenaline secretion from the adrenal medulla [72]. Pretreatment with a β-blocker and adrenal demedullation largely attenuate capsaicin-induced increases in O2 consumption [54, 69]. Capsaicin-induced adrenaline secretion is inhibited by chemical denervation of sensory nerves [73], pretreatment with the TRPV1 antagonist capsazepine [74], and in TRPV1-KO mice (Fig. 1). In Fig. 1, basal adrenaline level in adrenal vein in TRPV1-KO mice were lower than that in wild-type mice. Previous reports indicate that TRPV1-KO mice show a decreased sympathetic activity although basal body temperature, heart rate and blood pressure are normal [75–77], therefore the lowering of adrenaline secretion in TRPV1-KO might be due to decreasing activity of sympathoadrenal nerves. Moreover, not only the pungent TRPV1 agonist capsaicin but also the low pungency TRPV1 agonists capsiate [78] and [10] -shogaol [61] could induce adrenaline secretion. These results indicate that peripheral administration of TRPV1 agonists can evoke adrenaline secretion via sensory—central—sympathoadrenal reflexes to increase energy expenditure (Fig. 2). In addition, Osaka et al. reported that the rostral ventrolateral medulla, which is the site of the premotor area that contains sympathoadrenal preganglionic neurons, is a critical locus for capsaicin-mediated increases in energy expenditure [79].
Fig. 1.
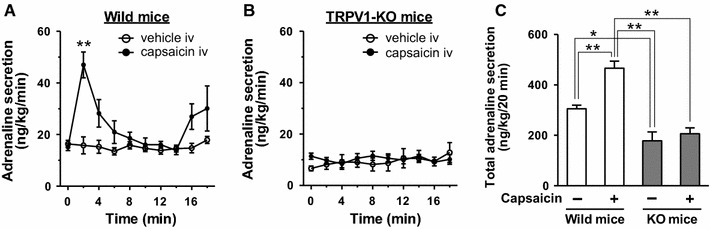
Capsaicin-induced adrenaline secretion is completely impaired in TRPV1 knockout mice. We investigated the effect of capsaicin on adrenaline secretion in male adult C57BL/6 wild-type mice and TRPV1 knockout mice (obtained from Dr. D. Julius, University of California, San Francisco) according to the previous report with a slight modification [89]. A mouse anesthetized with α-chloralose and urethane (0.1 and 1 g/kg, respectively) was placed on heated pad, and the rectal temperature was maintained at 36.5–37.5 °C. Heparinized saline (500 IU/ml, 25 µl) was injected through the femoral vein, then sampling of adrenal blood from the adrenal vein was started with a 2-min interval. Immediately after collecting the first fraction, capsaicin (0.05 mg/kg) or vehicle (saline containing 2% ethanol and 10% Tween-80) was administered into the femoral vein. Plasma adrenaline was purified with active alumina and measured by HPLC-electrochemical detection. Intravenous administration of capsaicin significantly increased adrenaline secretion in wild-type mice (a) but not TRPV1 knockout mice (b). There was a significant difference between vehicle and capsaicin in a but not b (treatment effect, p < 0.01 by two-way ANOVA). **p < 0.01 by Bonferroni’s test. c Total adrenaline secretion for 20 min in a and b. *p < 0.05, **p < 0.01 by one-way ANOVA followed by Tukey’s test. Each value is the mean ± SEM (n = 5)
Fig. 2.
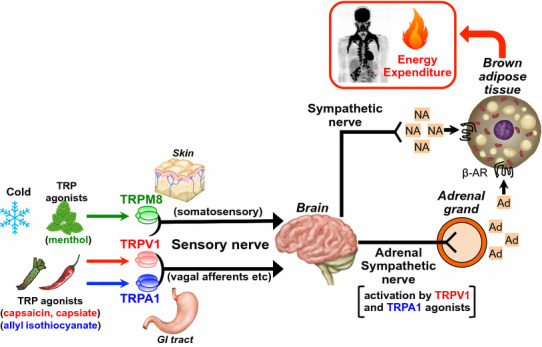
Mechanisms of increasing energy expenditure and thermogenesis by sensory nerves expressing thermo-TRP channels. Ad adrenaline, NA noradrenaline, GI tract gastrointestinal tract, β-AR β-adrenergic receptor.
Modified with permission from Ref. [126]
Increases in O2 consumption by capsaicin are partially retained in adrenal-demedullated rats [69], suggesting that mechanisms other than those involving adrenaline could underlie the increase in energy expenditure. Capsaicin and capsiate activate sympathetic efferent nerves innervating iBAT, thereby inducing expression of UCP1 and heat production [66, 69, 80, 81]. Sympathetic denervation of iBAT partly attenuates capsaicin-induced energy expenditure [69]. The administration of capsaicin or capsiate into the GI tract increases the activity of sympathetic efferent nerves innervating iBAT and induces thermogenesis in iBAT. These effects can be impaired by the denervation of vagal afferents and extrinsic nerves connected to the jejunum [66, 81]. Subchronic intake of the non-pungent TRPV1 agonist capsiate or 1-monoolein elevates UCP1 expression in iBAT and prevents accumulation of visceral fat promoted by a high-fat diet [64, 80]. Furthermore, the anti-obesity effects of capsiate are completely abolished in UCP1-KO mice [82]. Together, these findings indicate that TRPV1-expressing sensory nerves, especially vagal afferent sensory nerves, participate in regulating activity of sympathetic nerves innervating the iBAT as well as energy expenditure (Fig. 2).
Many spicy foods contain both TRPV1 agonists and TRPA1 agonists [57]. For example, allyl isothiocyanate in mustard [83], cinnamaldehyde in cinnamon [83], and allicin and diallyl trisulfide in garlic [84, 85] are TRPA1 agonists. Additionally, we identified miogatrial from mioga (Zingiber mioga) as a potent and low pungency TRPA1 agonist [86]. TRPA1 is co-expressed with TRPV1 in dorsal root ganglion neurons and nodose ganglion neurons [87, 88], and similar to that of TRPV1 agonists, the TRPA1 agonists allyl isothiocyanate and cinnamaldehyde enhance adrenaline secretion via the sensory—central—sympathoadrenal reflex [89]. Allyl isothiocyanate and cinnamaldehyde induce UCP1 expression in iBAT [90–92] and heat production [93]. Therefore, sensory neurons expressing both TRPV1 and TRPA1 might be a crucial subclass that regulates energy expenditure and thermogenesis (Fig. 2).
Cold exposure is known to enhance thermogenesis to maintain body temperature. TRPM8 is a cold receptor that is activated by moderate cold (<25–28 °C) and “cooling-mimetic” compounds such as menthol from mint [94, 95]. TRPM8 is abundantly expressed in different subpopulations of sensory neurons (from dorsal root, trigeminal and nodose ganglion) that express TRPV1 and TRPA1 [87, 96–98]. Previous reports using TRPM8-deficient mice show that TRPM8 mediates cold sensations to induce avoidance behavior towards innocuous cold [99–101]. Moreover, recent studies demonstrated that TRPM8 is a crucial channel for cold-defense thermoregulation. The core body temperature in wild-type mice is maintained during cold exposure, but the core temperature of TRPM8-KO mice and wild-type mice treated with a TRPM8 antagonist is decreased [102, 103]. Moreover, activation of TRPM8 by cold stimulation or menthol administration increases core temperature in wild-type mice but not TRPM8-KO mice [93, 102]. Cold exposure activates and increases c-Fos expression in the lateral parabrachial nucleus, which is relay area for cutaneous cold signals from primary sensory neurons to the preoptic hypothalamus [104, 105]; c-Fos expression is blunted by treatment with TRPM8 antagonists [103]. Taken together with these data, activation of TRPM8, presumably on sensory nerves, increases heat production (Fig. 2). TRPM8 expression has also been detected outside of sensory nerves in tissues such as the bladder, prostate, brown adipocyte, liver, gastrointestinal mucosa and several types of tumors. As such, the above-mentioned results using TRPM8-KO mice and TRPM8 antagonists might contribute to an understanding of TRPM8 mechanisms beyond that in sensory nerves. Future studies using site-specific loss of function approaches could clarify the mechanism by which TRPM8 in sensory nerves regulates thermogenesis.
TRP-activated brown fat thermogenesis in humans
In humans, with the exception of newborns, the prevalence of BAT has long been believed to be negligible. However, recent radionuclide imaging studies revealed the existence of considerable amounts of BAT in healthy adults [106, 107]. The metabolic activity of human BAT can be assessed by fluorodeoxyglucose (FDG)-positron emission tomography (PET) combined with X-ray computed tomography (CT), which is a powerful diagnostic tool for malignant tumors. Although the principal substrate for BAT thermogenesis is fatty acids, glucose utilization is greatly enhanced in parallel with the activation of UCP1, a key molecule of BAT thermogenesis [108]. Thus, glucose utilization assessed by FDG uptake could serve as an index of BAT thermogenic activity.
Cold exposure is the most powerful and physiological stimulus for BAT activation. Cold acts on TRP expressed in sensory nerves to enhance sympathetic nerve activity and trigger β-adrenergic receptor-mediated intracellular cascades in brown adipocytes, and finally to activate UCP1 and thermogenesis (Fig. 2) [104, 109]. In fact, the activity of BAT as assessed by FDG-PET/CT is greatly increased after acute cold exposure or administration of β-adrenergic receptor agonists, but is reduced under warm conditions or by pretreatment with a β-adrenergic blocker [106, 110, 111]. BAT activity is positively associated with cold-induced non-shivering thermogenesis (CIT), suggesting that BAT contributes to whole-body energy expenditure in humans [112]. BAT activity decreases with age and these decreases are associated with excessive accumulation of body fat with age [113]. Inactivation and reduction of BAT are now accepted to be associated with obesity and insulin resistance [114, 115]. As such, methods that re-activate and recruit BAT could promote reductions in body fat. Indeed, repeated mild cold exposure at 16 °C for 2 h every day for 6 weeks results in increases in BAT activity and CIT in healthy lean subjects [116]. More importantly, such cold acclimation decreases body fat mass, in parallel with the changes in BAT activity and CIT. This result is in line with a recent report showing increased BAT mass and CIT after cold acclimation in obese subjects [117]. Thus, BAT is a significant anti-obesity target in humans [118].
Nevertheless, increased exposure to cold would be difficult and uncomfortable in daily life. As mentioned above, some thermo-TRP channels are activated by various chemical substances, including food ingredients. Given that chemical activation of TRPV1 in the gastrointestinal tract by capsaicin and capsiate activates UCP1 in BAT in mice [82], oral administration of these ingredients may be a more feasible method to recruit BAT in humans. We found that single oral ingestion of capsinoids, which include capsiate, dihydrocapsiate, and nordihydrocapsiate, increases whole-body energy expenditure in human individuals with metabolically active BAT, but not in those without active BAT [119]. Furthermore, daily ingestion of capsinoids augments BAT activity [120] and CIT [116] even in individuals with low BAT activities. Thermogenic and fat-reducing effects of capsinoids have also been shown in an obese population [55, 121, 122]. Taken together, the anti-obesity effects of capsinoids as TRPV1 agonists may be attributable to the thermogenic activity of recruited BAT.
The mechanism of capsinoid-induced activation of BAT has been characterized for the most part by studies in small rodents. As noted above, the primary action site of capsinoids would be TRPV1 on sensory nerves in the gastrointestinal tract. Consistent with the crucial role of TRPV1 in capsinoid-mediated effects in mice [66], the beneficial effects of capsinoids are greatly attenuated in individuals who carry a mutated (Val585Ile) TRPV1 [121]. Although TRPV1 is expressed in brown adipocytes, the direct action of capsinoids toward TRPV1 in human BAT may be unlikely because orally ingested capsinoids are rapidly hydrolyzed, and thus are usually undetectable in the general circulation in humans. Although TRPV1 activators affect BAT and body fat similarly to cold exposure, it should be noted that TRPV1 is not a cold sensor but instead is a sensor of noxious stimuli, including exposure to temperatures above 42 °C. Thus, human BAT is likely to be activated by nociceptive stimuli such as high temperature and capsinoids. In agreement with this idea, Sidossis et al. [123] recently demonstrated in humans that chronic adrenergic stress induced by burn trauma results in browning of WAT.
In addition to capsinoids, several dietary substances activate TRPV1 and BAT thermogenesis. For example, dietary supplementation with eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA) up-regulates UCP1 expression in both BAT and WAT, thereby preventing diet-induced obesity [124]. These effects are largely dependent on the presence of TRPV1 and activation of sympathetic nervous system (SNS) [76]. Despite the evidence in mice, effects of EPA and DHA on human BAT have not been determined. Additionally, as noted above, TRPV1-expressing sensory nerves also express cold-sensitive TRPA1, which is involved in BAT activation induced by cold exposure. TRPA1 is activated by various pungent compounds, such as allyl- and benzyl-isothiocyanates in wasabi (Japanese horse radish) and cinnamaldehyde in cinnamon. These compounds are known to increase thermogenesis and UCP1 expression in small rodents [125]. Furthermore, TRPM8 is also the most likely candidate receptor to sense lower temperatures and is known to be involved in BAT activation [125]. A representative TRPM8 agonist is menthol, a cooling and flavor compound in mint [93]. Using TRPM8- and UCP1-deficient mice, Ma et al. documented that dietary menthol activates UCP1-dependent thermogenesis in a TRPM8-dependent manner [48]. These findings concerning the chemical activation of the TRPs-SNS-BAT axis in small rodents provide further impetus for the identification of common food ingredients that can activate and recruit BAT in humans.
Conclusions and perspectives
The obesity pandemic is a serious global health problem because it is a major risk factor for metabolic syndromes including insulin resistance, impaired insulin secretion, hyperglycemia, dyslipidemia, and hypertension. Normal regulation of glucose and lipid metabolism is indispensable for healthy biological activity. Moreover, enhancement of energy expenditure together with a reduction in food intake is effective in reducing obesity. Eleven thermo-TRP channels have been identified in the 20 years since the first thermo-TRP channel, TRPV1, was cloned. Furthermore, these thermo-TRPs not only sense temperature but also regulate events related to energy metabolism, such as insulin secretion by the pancreas, differentiation and/or thermogenesis in brown adipocytes, and energy expenditure mediated by sensory nerve—brain—sympathetic reflexes. We anticipate that further studies on the physiological and pathophysiological roles of thermo-TRPs may open novel avenues for treating metabolic disorders, including obesity and diabetes.
Acknowledgements
We would like to thank Manabu Tanabe at University of Shizuoka for supporting the experiment of adrenaline measurements in TRPV1 knockout mice.
Compliance with ethical standards
Conflict of interest
All authors have no conflicts of interest related to this manuscript.
Funding
This work was supported in part by a Grant-in-Aid for Research Fellowship (17-5490) from the Japan Society for the Promotion of Science to Y. Iwasaki.
Ethical approval
All animal experiments were approved by the Animal Care and Use Committee of University of Shizuoka.
Footnotes
Kunitoshi Uchida, Katsuya Dezaki, and Takeshi Yoneshiro contributed equally to this work.
Contributor Information
Kunitoshi Uchida, Phone: +81-92-801-0411, Email: uchida@college.fdcnet.ac.jp.
Yusaku Iwasaki, Phone: +81-285-58-7320, Email: ysk-iwasaki@jichi.ac.jp.
References
- 1.Montell C, Rubin GM. Molecular characterization of the Drosophila trp locus: a putative integral membrane protein required for phototransduction. Neuron. 1989;2:1313–1323. doi: 10.1016/0896-6273(89)90069-X. [DOI] [PubMed] [Google Scholar]
- 2.Venkatachalam K, Montell C. TRP channels. Annu Rev Biochem. 2007;76:387–417. doi: 10.1146/annurev.biochem.75.103004.142819. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Hilton JK, Rath P, Helsell CV, Beckstein O, Van Horn WD. Understanding thermosensitive transient receptor potential channels as versatile polymodal cellular sensors. Biochemistry. 2015;54:2401–2413. doi: 10.1021/acs.biochem.5b00071. [DOI] [PubMed] [Google Scholar]
- 4.Caterina MJ, Schumacher MA, Tominaga M, Rosen TA, Levine JD, et al. The capsaicin receptor: a heat-activated ion channel in the pain pathway. Nature. 1997;389:816–824. doi: 10.1038/39807. [DOI] [PubMed] [Google Scholar]
- 5.Vriens J, Owsianik G, Hofmann T, Philipp SE, Stab J, et al. TRPM3 is a nociceptor channel involved in the detection of noxious heat. Neuron. 2011;70:482–494. doi: 10.1016/j.neuron.2011.02.051. [DOI] [PubMed] [Google Scholar]
- 6.Zimmermann K, Lennerz JK, Hein A, Link AS, Kaczmarek JS, et al. Transient receptor potential cation channel, subfamily C, member 5 (TRPC5) is a cold-transducer in the peripheral nervous system. Proc Natl Acad Sci USA. 2011;108:18114–18119. doi: 10.1073/pnas.1115387108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.World Health Organization. Obesity and overweight: fact sheet N311. http://www.who.int/mediacentre/factsheets/fs311/en/. Accessed June 2016
- 8.Kelly T, Yang W, Chen CS, Reynolds K, He J. Global burden of obesity in 2005 and projections to 2030. Int J Obes. 2008;32:1431–1437. doi: 10.1038/ijo.2008.102. [DOI] [PubMed] [Google Scholar]
- 9.Uchida K, Tominaga M. The role of TRPM2 in pancreatic β-cells and the development of diabetes. Cell Calcium. 2014;56:332–339. doi: 10.1016/j.ceca.2014.07.001. [DOI] [PubMed] [Google Scholar]
- 10.Togashi K, Hara Y, Tominaga T, Higashi T, Konishi Y, et al. TRPM2 activation by cyclic ADP-ribose at body temperature is involved in insulin secretion. EMBO J. 2006;25:1804–1815. doi: 10.1038/sj.emboj.7601083. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Yosida M, Dezaki K, Uchida K, Kodera S, Lam NV, et al. Involvement of cAMP/EPAC/TRPM2 activation in glucose- and incretin-induced insulin secretion. Diabetes. 2014;63:3394–3403. doi: 10.2337/db13-1868. [DOI] [PubMed] [Google Scholar]
- 12.Ahren B. Islet G protein-coupled receptors as potential targets for treatment of type 2 diabetes. Nat Rev Drug Discov. 2009;8:369–385. doi: 10.1038/nrd2782. [DOI] [PubMed] [Google Scholar]
- 13.Campbell JE, Drucker DJ. Pharmacology, physiology, and mechanisms of incretin hormone action. Cell Metab. 2013;17:819–837. doi: 10.1016/j.cmet.2013.04.008. [DOI] [PubMed] [Google Scholar]
- 14.Uchida K, Dezaki K, Damdindorj B, Inada H, Shiuchi T, et al. Lack of TRPM2 impaired insulin secretion and glucose metabolisms in mice. Diabetes. 2011;60:119–126. doi: 10.2337/db10-0276. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Dezaki K, Hosoda H, Kakei M, Hashiguchi S, Watanabe M, et al. Endogenous ghrelin in pancreatic islets restricts insulin release by attenuating Ca2+ signaling in β-cells: implication in the glycemic control in rodents. Diabetes. 2004;53:3142–3151. doi: 10.2337/diabetes.53.12.3142. [DOI] [PubMed] [Google Scholar]
- 16.Dezaki K, Sone H, Koizumi M, Nakata M, Kakei M, et al. Blockade of pancreatic islet-derived ghrelin enhances insulin secretion to prevent high-fat diet-induced glucose intolerance. Diabetes. 2006;55:3486–3493. doi: 10.2337/db06-0878. [DOI] [PubMed] [Google Scholar]
- 17.Dezaki K, Kakei M, Yada T. Ghrelin uses Gαi2 and activates voltage-dependent K+ channels to attenuate glucose-induced Ca2+ signaling and insulin release in islet β-cells: novel signal transduction of ghrelin. Diabetes. 2007;56:2319–2327. doi: 10.2337/db07-0345. [DOI] [PubMed] [Google Scholar]
- 18.Dezaki K, Damdindorj B, Sone H, Dyachok O, Tengholm A, et al. Ghrelin attenuates cAMP-PKA signaling to evoke insulinostatic cascade in islet β-cells. Diabetes. 2011;60:2315–2324. doi: 10.2337/db11-0368. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Kurashina T, Dezaki K, Yoshida M, Sukma Rita R, Ito K, et al. The β-cell GHSR and downstream cAMP/TRPM2 signaling account for insulinostatic and glycemic effects of ghrelin. Sci Rep. 2015;5:14041. doi: 10.1038/srep14041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Damdindorj B, Dezaki K, Kurashina T, Sone H, Rita R, et al. Exogenous and endogenous ghrelin counteracts GLP-1 action to stimulate cAMP signaling and insulin secretion in islet β-cells. FEBS Lett. 2012;586:2555–2562. doi: 10.1016/j.febslet.2012.06.034. [DOI] [PubMed] [Google Scholar]
- 21.Ito K, Dezaki K, Yoshida M, Yamada H, Miura R, et al. Endogenous α2A-adrenoceptor-operated sympathoadrenergic tones attenuate insulin secretion via cAMP/TRPM2 signaling. Diabetes. 2017;66(3):699–709. doi: 10.2337/db16-1166. [DOI] [PubMed] [Google Scholar]
- 22.Cummings DE, Purnell JQ, Frayo RS, Schmidova K, Wisse BE, et al. A preprandial rise in plasma ghrelin levels suggests a role in meal initiation in humans. Diabetes. 2001;50:1714–1719. doi: 10.2337/diabetes.50.8.1714. [DOI] [PubMed] [Google Scholar]
- 23.Wagner TF, Loch S, Lambert S, Straub I, Mannebach S, et al. Transient receptor potential M3 channels are ionotropic steroid receptors in pancreatic β cells. Nat Cell Biol. 2008;10:1421–1430. doi: 10.1038/ncb1801. [DOI] [PubMed] [Google Scholar]
- 24.Klose C, Straub I, Riehle M, Ranta F, Krautwurst D, et al. Fenamates as TRP channel blockers: mefenamic acid selectively blocks TRPM3. Br J Pharmacol. 2011;162:1757–1769. doi: 10.1111/j.1476-5381.2010.01186.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Cheng H, Beck A, Launay P, Gross SA, Stokes AJ, et al. TRPM4 controls insulin secretion in pancreatic β-cells. Cell Calcium. 2007;41:51–61. doi: 10.1016/j.ceca.2006.04.032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Krishnan K, Ma Z, Bjorklund A, Islam MS. Role of transient receptor potential melastatin-like subtype 5 channel in insulin secretion from rat β-cells. Pancreas. 2014;43:597–604. doi: 10.1097/MPA.0000000000000027. [DOI] [PubMed] [Google Scholar]
- 27.Shigeto M, Ramracheya R, Tarasov AI, Cha CY, Chibalina MV, et al. GLP-1 stimulates insulin secretion by PKC-dependent TRPM4 and TRPM5 activation. J Clin Invest. 2015;125:4714–4728. doi: 10.1172/JCI81975. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Cao DS, Zhong L, Hsieh TH, Abooj M, Bishnoi M, et al. Expression of transient receptor potential ankyrin 1 (TRPA1) and its role in insulin release from rat pancreatic β cells. PLoS One. 2012;7:e38005. doi: 10.1371/journal.pone.0038005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Babes A, Fischer MJ, Filipovic M, Engel MA, Flonta ML, et al. The anti-diabetic drug glibenclamide is an agonist of the transient receptor potential Ankyrin 1 (TRPA1) ion channel. Eur J Pharmacol. 2013;704:15–22. doi: 10.1016/j.ejphar.2013.02.018. [DOI] [PubMed] [Google Scholar]
- 30.Caterina MJ, Rosen TA, Tominaga M, Brake AJ, Julius D. A capsaicin-receptor homologue with a high threshold for noxious heat. Nature. 1999;398:436–441. doi: 10.1038/18906. [DOI] [PubMed] [Google Scholar]
- 31.Muraki K, Iwata Y, Katanosaka Y, Ito T, Ohya S, et al. TRPV2 is a component of osmotically sensitive cation channels in murine aortic myocytes. Circ Res. 2003;93:829–838. doi: 10.1161/01.RES.0000097263.10220.0C. [DOI] [PubMed] [Google Scholar]
- 32.Juvin V, Penna A, Chemin J, Lin YL, Rassendren FA. Pharmacological characterization and molecular determinants of the activation of transient receptor potential V2 channel orthologs by 2-aminoethoxydiphenyl borate. Mol Pharmacol. 2007;72:1258–1268. doi: 10.1124/mol.107.037044. [DOI] [PubMed] [Google Scholar]
- 33.Monet M, Gkika D, Lehen’kyi V, Pourtier A, Vanden Abeele F, et al. Lysophospholipids stimulate prostate cancer cell migration via TRPV2 channel activation. Biochim Biophys Acta. 2009;1793:528–539. doi: 10.1016/j.bbamcr.2009.01.003. [DOI] [PubMed] [Google Scholar]
- 34.Shibasaki K, Murayama N, Ono K, Ishizaki Y, Tominaga M. TRPV2 enhances axon outgrowth through its activation by membrane stretch in developing sensory and motor neurons. J Neurosci. 2010;30:4601–4612. doi: 10.1523/JNEUROSCI.5830-09.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Mihara H, Boudaka A, Shibasaki K, Yamanaka A, Sugiyama T, et al. Involvement of TRPV2 activation in intestinal movement through nitric oxide production in mice. J Neurosci. 2010;30:16536–16544. doi: 10.1523/JNEUROSCI.4426-10.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Shibasaki K. Physiological significance of TRPV2 as a mechanosensor, thermosensor and lipid sensor. J Physiol Sci. 2016;66:359–365. doi: 10.1007/s12576-016-0434-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Link TM, Park U, Vonakis BM, Raben DM, Soloski MJ, et al. TRPV2 has a pivotal role in macrophage particle binding and phagocytosis. Nat Immunol. 2010;11:232–239. doi: 10.1038/ni.1842. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Sun W, Uchida K, Suzuki Y, Zhou Y, Kim M, et al. Lack of TRPV2 impairs thermogenesis in mouse brown adipose tissue. EMBO Rep. 2016;17:383–399. doi: 10.15252/embr.201540819. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Iwata Y, Katanosaka Y, Arai Y, Shigekawa M, Wakabayashi S. Dominant-negative inhibition of Ca2+ influx via TRPV2 ameliorates muscular dystrophy in animal models. Hum Mol Genet. 2009;18:824–834. doi: 10.1093/hmg/ddn408. [DOI] [PubMed] [Google Scholar]
- 40.De Petrocellis L, Ligresti A, Moriello AS, Allara M, Bisogno T, et al. Effects of cannabinoids and cannabinoid-enriched Cannabis extracts on TRP channels and endocannabinoid metabolic enzymes. Br J Pharmacol. 2011;163:1479–1494. doi: 10.1111/j.1476-5381.2010.01166.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Kanzaki M, Zhang YQ, Mashima H, Li L, Shibata H, et al. Translocation of a calcium-permeable cation channel induced by insulin-like growth factor-I. Nat Cell Biol. 1999;1:165–170. doi: 10.1038/11086. [DOI] [PubMed] [Google Scholar]
- 42.Stokes AJ, Shimoda LM, Koblan-Huberson M, Adra CN, Turner H. A TRPV2-PKA signaling module for transduction of physical stimuli in mast cells. J Exp Med. 2004;200:137–147. doi: 10.1084/jem.20032082. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Sun W, Uchida K, Takahashi N, Iwata Y, Wakabayashi S, et al. Activation of TRPV2 negatively regulates the differentiation of mouse brown adipocytes. Pflugers Arch. 2016;468:1527–1540. doi: 10.1007/s00424-016-1846-1. [DOI] [PubMed] [Google Scholar]
- 44.Neal JW, Clipstone NA. Calcineurin mediates the calcium-dependent inhibition of adipocyte differentiation in 3T3-L1 cells. J Biol Chem. 2002;277:49776–49781. doi: 10.1074/jbc.M207913200. [DOI] [PubMed] [Google Scholar]
- 45.Kida R, Yoshida H, Murakami M, Shirai M, Hashimoto O, et al. Direct action of capsaicin in brown adipogenesis and activation of brown adipocytes. Cell Biochem Funct. 2016;34:34–41. doi: 10.1002/cbf.3162. [DOI] [PubMed] [Google Scholar]
- 46.Baboota RK, Singh DP, Sarma SM, Kaur J, Sandhir R, et al. Capsaicin induces “brite” phenotype in differentiating 3T3-L1 preadipocytes. PLoS One. 2014;9:e103093. doi: 10.1371/journal.pone.0103093. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Bishnoi M, Kondepudi KK, Gupta A, Karmase A, Boparai RK. Expression of multiple Transient Receptor Potential channel genes in murine 3T3-L1 cell lines and adipose tissue. Pharmacol Rep. 2013;65:751–755. doi: 10.1016/S1734-1140(13)71055-7. [DOI] [PubMed] [Google Scholar]
- 48.Ma S, Yu H, Zhao Z, Luo Z, Chen J, et al. Activation of the cold-sensing TRPM8 channel triggers UCP1-dependent thermogenesis and prevents obesity. J Mol Cell Biol. 2012;4:88–96. doi: 10.1093/jmcb/mjs001. [DOI] [PubMed] [Google Scholar]
- 49.Rossato M, Granzotto M, Macchi V, Porzionato A, Petrelli L, et al. Human white adipocytes express the cold receptor TRPM8 which activation induces UCP1 expression, mitochondrial activation and heat production. Mol Cell Endocrinol. 2014;383:137–146. doi: 10.1016/j.mce.2013.12.005. [DOI] [PubMed] [Google Scholar]
- 50.Ye L, Kleiner S, Wu J, Sah R, Gupta RK, et al. TRPV4 is a regulator of adipose oxidative metabolism, inflammation, and energy homeostasis. Cell. 2012;151:96–110. doi: 10.1016/j.cell.2012.08.034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Kusudo T, Wang Z, Mizuno A, Suzuki M, Yamashita H. TRPV4 deficiency increases skeletal muscle metabolic capacity and resistance against diet-induced obesity. J Appl Physiol. 2012;112:1223–1232. doi: 10.1152/japplphysiol.01070.2011. [DOI] [PubMed] [Google Scholar]
- 52.Henry CJK, Emery B. Effect of spiced food on metabolic rate. Hum Nutr Clin Nutr. 1986;40:165–168. [PubMed] [Google Scholar]
- 53.Kawada T, Hagihara K, Iwai K. Effects of capsaicin on lipid metabolism in rats fed a high fat diet. J Nutr. 1986;116:1272–1278. doi: 10.1093/jn/116.7.1272. [DOI] [PubMed] [Google Scholar]
- 54.Kawada T, Watanabe T, Takaishi T, Tanaka T, Iwai K. Capsaicin-induced β-adrenergic action on energy metabolism in rats: influence of capsaicin on oxygen consumption, the respiratory quotient, and substrate utilization. Proc Soc Exp Biol Med. 1986;183:250–256. doi: 10.3181/00379727-183-42414. [DOI] [PubMed] [Google Scholar]
- 55.Ludy MJ, Moore GE, Mattes RD. The effects of capsaicin and capsiate on energy balance: critical review and meta-analyses of studies in humans. Chem Senses. 2012;37:103–121. doi: 10.1093/chemse/bjr100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Whiting S, Derbyshire E, Tiwari BK. Capsaicinoids and capsinoids. A potential role for weight management? A systematic review of the evidence. Appetite. 2012;59:341–348. doi: 10.1016/j.appet.2012.05.015. [DOI] [PubMed] [Google Scholar]
- 57.Vetter I, Lewis RJ. Natural product ligands of TRP channels. Adv Exp Med Biol. 2011;704:41–85. doi: 10.1007/978-94-007-0265-3_3. [DOI] [PubMed] [Google Scholar]
- 58.McNamara FN, Randall A, Gunthorpe MJ. Effects of piperine, the pungent component of black pepper, at the human vanilloid receptor (TRPV1) Br J Pharmacol. 2005;144:781–790. doi: 10.1038/sj.bjp.0706040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Okumura Y, Narukawa M, Iwasaki Y, Ishikawa A, Matsuda H, et al. Activation of TRPV1 and TRPA1 by black pepper components. Biosci Biotechnol Biochem. 2010;74:1068–1072. doi: 10.1271/bbb.90964. [DOI] [PubMed] [Google Scholar]
- 60.Witte DG, Cassar SC, Masters JN, Esbenshade T, Hancock AA. Use of a fluorescent imaging plate reader-based calcium assay to assess pharmacological differences between the human and rat vanilloid receptor. J Biomol Screen. 2002;7:466–475. doi: 10.1177/108705702237679. [DOI] [PubMed] [Google Scholar]
- 61.Iwasaki Y, Morita A, Iwasawa T, Kobata K, Sekiwa Y, et al. A nonpungent component of steamed ginger—[10]-shogaol- increases adrenaline secretion via the activation of TRPV1. Nutr Neurosci. 2006;9:169–178. doi: 10.1080/110284150600955164. [DOI] [PubMed] [Google Scholar]
- 62.Kobata K, Todo T, Yazawa S, Iwai K, Watanabe T. Novel capsaicinoid-like substances, capsiate and dihydrocapsiate, from the fruits of a nonpungent cultivar, CH-19 Sweet, of pepper (Capsicum annuum L.) J Agric Food Chem. 1998;46:1695–1697. doi: 10.1021/jf980135c. [DOI] [Google Scholar]
- 63.Iida T, Moriyama T, Kobata K, Morita A, Murayama N, et al. TRPV1 activation and induction of nociceptive response by a non-pungent capsaicin-like compound, capsiate. Neuropharmacology. 2003;44:958–967. doi: 10.1016/S0028-3908(03)00100-X. [DOI] [PubMed] [Google Scholar]
- 64.Iwasaki Y, Saito O, Tanabe M, Inayoshi K, Kobata K, et al. Monoacylglycerols activate capsaicin receptor, TRPV1. Lipids. 2008;43:471–483. doi: 10.1007/s11745-008-3182-5. [DOI] [PubMed] [Google Scholar]
- 65.Ohnuki K, Haramizu S, Watanabe T, Yazawa S, Fushiki T. CH-19 sweet, nonpungent cultivar of red pepper, increased body temperature in mice with vanilloid receptors stimulation by capsiate. J Nutr Sci Vitaminol. 2001;47:295–298. doi: 10.3177/jnsv.47.295. [DOI] [PubMed] [Google Scholar]
- 66.Kawabata F, Inoue N, Masamoto Y, Matsumura S, Kimura W, et al. Non-pungent capsaicin analogs (capsinoids) increase metabolic rate and enhance thermogenesis via gastrointestinal TRPV1 in mice. Biosci Biotechnol Biochem. 2009;73:2690–2697. doi: 10.1271/bbb.90555. [DOI] [PubMed] [Google Scholar]
- 67.Helliwell RJ, McLatchie LM, Clarke M, Winter J, Bevan S, et al. Capsaicin sensitivity is associated with the expression of the vanilloid (capsaicin) receptor (VR1) mRNA in adult rat sensory ganglia. Neurosci Lett. 1998;250:177–180. doi: 10.1016/S0304-3940(98)00475-3. [DOI] [PubMed] [Google Scholar]
- 68.Tominaga M, Caterina MJ, Malmberg AB, Rosen TA, Gilbert H, et al. The cloned capsaicin receptor integrates multiple pain-producing stimuli. Neuron. 1998;21:531–543. doi: 10.1016/S0896-6273(00)80564-4. [DOI] [PubMed] [Google Scholar]
- 69.Kobayashi A, Osaka T, Namba Y, Inoue S, Lee TH, et al. Capsaicin activates heat loss and heat production simultaneously and independently in rats. Am J Physiol. 1998;275:R92–R98. doi: 10.1152/ajpregu.1998.275.1.R92. [DOI] [PubMed] [Google Scholar]
- 70.Astrup A, Simonsen L, Bulow J, Madsen J, Christensen NJ. Epinephrine mediates facultative carbohydrate-induced thermogenesis in human skeletal muscle. Am J Physiol. 1989;257:E340–E345. doi: 10.1152/ajpendo.1989.257.3.E340. [DOI] [PubMed] [Google Scholar]
- 71.Watanabe T, Kawada T, Kurosawa M, Sato A, Iwai K. Adrenal sympathetic efferent nerve and catecholamine secretion excitation caused by capsaicin in rats. Am J Physiol. 1988;255:E23–E27. doi: 10.1152/ajpendo.1988.255.1.E23. [DOI] [PubMed] [Google Scholar]
- 72.Watanabe T, Kawada T, Yamamoto M, Iwai K. Capsaicin, a pungent principle of hot red pepper, evokes catecholamine secretion from the adrenal medulla of anesthetized rats. Biochem Biophys Res Commun. 1987;142:259–264. doi: 10.1016/0006-291X(87)90479-7. [DOI] [PubMed] [Google Scholar]
- 73.Watanabe T, Kawada T, Iwai K. Effect of capsaicin pretreatment on capsaicin-induced catecholamine secretion from the adrenal medulla in rats. Proc Soc Exp Biol Med. 1988;187:370–374. doi: 10.3181/00379727-187-3-RC1. [DOI] [PubMed] [Google Scholar]
- 74.Watanabe T, Sakurada N, Kobata K. Capsaicin-, resiniferatoxin-, and olvanil-induced adrenaline secretions in rats via the vanilloid receptor. Biosci Biotechnol Biochem. 2001;65:2443–2447. doi: 10.1271/bbb.65.2443. [DOI] [PubMed] [Google Scholar]
- 75.Alawi KM, Aubdool AA, Liang L, Wilde E, Vepa A, et al. The sympathetic nervous system is controlled by transient receptor potential vanilloid 1 in the regulation of body temperature. FASEB J. 2015;29:4285–4298. doi: 10.1096/fj.15-272526. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Kim M, Goto T, Yu R, Uchida K, Tominaga M, et al. Fish oil intake induces UCP1 upregulation in brown and white adipose tissue via the sympathetic nervous system. Sci Rep. 2015;5:18013. doi: 10.1038/srep18013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Pacher P, Batkai S, Kunos G. Haemodynamic profile and responsiveness to anandamide of TRPV1 receptor knock-out mice. J Physiol. 2004;558:647–657. doi: 10.1113/jphysiol.2004.064824. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Iwai K, Yazawa A, Watanabe T. Roles as metabolic regulators of the non-nutrients, capsaicin and capsiate, supplemented to diets. Proc Japan Acad. 2003;79B:207–212. doi: 10.2183/pjab.79B.207. [DOI] [Google Scholar]
- 79.Osaka T, Lee TH, Kobayashi A, Inoue S, Kimura S. Thermogenesis mediated by a capsaicin-sensitive area in the ventrolateral medulla. NeuroReport. 2000;11:2425–2428. doi: 10.1097/00001756-200008030-00017. [DOI] [PubMed] [Google Scholar]
- 80.Masuda Y, Haramizu S, Oki K, Ohnuki K, Watanabe T, et al. Upregulation of uncoupling proteins by oral administration of capsiate, a nonpungent capsaicin analog. J Appl Physiol. 2003;95:2408–2415. doi: 10.1152/japplphysiol.00828.2002. [DOI] [PubMed] [Google Scholar]
- 81.Ono K, Tsukamoto-Yasui M, Hara-Kimura Y, Inoue N, Nogusa Y, et al. Intragastric administration of capsiate, a transient receptor potential channel agonist, triggers thermogenic sympathetic responses. J Appl Physiol. 2011;110:789–798. doi: 10.1152/japplphysiol.00128.2010. [DOI] [PubMed] [Google Scholar]
- 82.Okamatsu-Ogura Y, Tsubota A, Ohyama K, Nogusa Y, Saito M, et al. Capsinoids suppress diet-induced obesity through uncoupling protein 1-dependent mechanism in mice. J Funct Foods. 2015;19:1–9. doi: 10.1016/j.jff.2015.09.005. [DOI] [Google Scholar]
- 83.Jordt SE, Bautista DM, Chuang HH, McKemy DD, Zygmunt PM, et al. Mustard oils and cannabinoids excite sensory nerve fibres through the TRP channel ANKTM1. Nature. 2004;427:260–265. doi: 10.1038/nature02282. [DOI] [PubMed] [Google Scholar]
- 84.Macpherson LJ, Geierstanger BH, Viswanath V, Bandell M, Eid SR, et al. The pungency of garlic: activation of TRPA1 and TRPV1 in response to allicin. Curr Biol. 2005;15:929–934. doi: 10.1016/j.cub.2005.04.018. [DOI] [PubMed] [Google Scholar]
- 85.Koizumi K, Iwasaki Y, Narukawa M, Iitsuka Y, Fukao T, et al. Diallyl sulfides in garlic activate both TRPA1 and TRPV1. Biochem Biophys Res Commun. 2009;382:545–548. doi: 10.1016/j.bbrc.2009.03.066. [DOI] [PubMed] [Google Scholar]
- 86.Iwasaki Y, Tanabe M, Kayama Y, Abe M, Kashio M, et al. Miogadial and miogatrial with α, β-unsaturated 1,4-dialdehyde moieties–novel and potent TRPA1 agonists. Life Sci. 2009;85:60–69. doi: 10.1016/j.lfs.2009.04.017. [DOI] [PubMed] [Google Scholar]
- 87.Story GM, Peier AM, Reeve AJ, Eid SR, Mosbacher J, et al. ANKTM1, a TRP-like channel expressed in nociceptive neurons, is activated by cold temperatures. Cell. 2003;112:819–829. doi: 10.1016/S0092-8674(03)00158-2. [DOI] [PubMed] [Google Scholar]
- 88.Choi MJ, Jin Z, Park YS, Rhee YK, Jin YH. Transient receptor potential (TRP) A1 activated currents in TRPV1 and cholecystokinin-sensitive cranial visceral afferent neurons. Brain Res. 2011;1383:36–42. doi: 10.1016/j.brainres.2011.02.009. [DOI] [PubMed] [Google Scholar]
- 89.Iwasaki Y, Tanabe M, Kobata K, Watanabe T. TRPA1 agonists -allyl isothiocyanate and cinnamaldehyde- induce adrenaline secretion. Biosci Biotechnol Biochem. 2008;72:2608–2614. doi: 10.1271/bbb.80289. [DOI] [PubMed] [Google Scholar]
- 90.Yoshida T, Yoshioka K, Wakabayashi Y, Nishioka H, Kondo M. Effects of capsaicin and isothiocyanate on thermogenesis of interscapular brown adipose tissue in rats. J Nutr Sci Vitaminol. 1988;34:587–594. doi: 10.3177/jnsv.34.587. [DOI] [PubMed] [Google Scholar]
- 91.Kawada T, Sakabe S, Aoki N, Watanabe T, Higeta K, et al. Intake of sweeteners and pungent ingredients increases the thermogenin content in brown adipose tissue of rat. J Agric Food Chem. 1991;39:651–654. doi: 10.1021/jf00004a004. [DOI] [Google Scholar]
- 92.Tamura Y, Iwasaki Y, Narukawa M, Watanabe T. Ingestion of cinnamaldehyde, a TRPA1 agonist, reduces visceral fats in mice fed a high-fat and high-sucrose diet. J Nutr Sci Vitaminol. 2012;58:9–13. doi: 10.3177/jnsv.58.9. [DOI] [PubMed] [Google Scholar]
- 93.Masamoto Y, Kawabata F, Fushiki T. Intragastric administration of TRPV1, TRPV3, TRPM8, and TRPA1 agonists modulates autonomic thermoregulation in different manners in mice. Biosci Biotechnol Biochem. 2009;73:1021–1027. doi: 10.1271/bbb.80796. [DOI] [PubMed] [Google Scholar]
- 94.McKemy DD, Neuhausser WM, Julius D. Identification of a cold receptor reveals a general role for TRP channels in thermosensation. Nature. 2002;416:52–58. doi: 10.1038/nature719. [DOI] [PubMed] [Google Scholar]
- 95.Peier AM, Moqrich A, Hergarden AC, Reeve AJ, Andersson DA, et al. A TRP channel that senses cold stimuli and menthol. Cell. 2002;108:705–715. doi: 10.1016/S0092-8674(02)00652-9. [DOI] [PubMed] [Google Scholar]
- 96.Kobayashi K, Fukuoka T, Obata K, Yamanaka H, Dai Y, et al. Distinct expression of TRPM8, TRPA1, and TRPV1 mRNAs in rat primary afferent neurons with aδ/c-fibers and colocalization with trk receptors. J Comp Neurol. 2005;493:596–606. doi: 10.1002/cne.20794. [DOI] [PubMed] [Google Scholar]
- 97.Takashima Y, Daniels RL, Knowlton W, Teng J, Liman ER, et al. Diversity in the neural circuitry of cold sensing revealed by genetic axonal labeling of transient receptor potential melastatin 8 neurons. J Neurosci. 2007;27:14147–14157. doi: 10.1523/JNEUROSCI.4578-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Kondo T, Obata K, Miyoshi K, Sakurai J, Tanaka J, et al. Transient receptor potential A1 mediates gastric distention-induced visceral pain in rats. Gut. 2009;58:1342–1352. doi: 10.1136/gut.2008.175901. [DOI] [PubMed] [Google Scholar]
- 99.Bautista DM, Siemens J, Glazer JM, Tsuruda PR, Basbaum AI, et al. The menthol receptor TRPM8 is the principal detector of environmental cold. Nature. 2007;448:204–208. doi: 10.1038/nature05910. [DOI] [PubMed] [Google Scholar]
- 100.Dhaka A, Murray AN, Mathur J, Earley TJ, Petrus MJ, et al. TRPM8 is required for cold sensation in mice. Neuron. 2007;54:371–378. doi: 10.1016/j.neuron.2007.02.024. [DOI] [PubMed] [Google Scholar]
- 101.Colburn RW, Lubin ML, Stone DJ, Jr, Wang Y, Lawrence D, et al. Attenuated cold sensitivity in TRPM8 null mice. Neuron. 2007;54:379–386. doi: 10.1016/j.neuron.2007.04.017. [DOI] [PubMed] [Google Scholar]
- 102.Tajino K, Hosokawa H, Maegawa S, Matsumura K, Dhaka A, et al. Cooling-sensitive TRPM8 is thermostat of skin temperature against cooling. PLoS One. 2011;6:e17504. doi: 10.1371/journal.pone.0017504. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Almeida MC, Hew-Butler T, Soriano RN, Rao S, Wang W, et al. Pharmacological blockade of the cold receptor TRPM8 attenuates autonomic and behavioral cold defenses and decreases deep body temperature. J Neurosci. 2012;32:2086–2099. doi: 10.1523/JNEUROSCI.5606-11.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Nakamura K. Central circuitries for body temperature regulation and fever. Am J Physiol Regul Integr Comp Physiol. 2011;301:R1207–R1228. doi: 10.1152/ajpregu.00109.2011. [DOI] [PubMed] [Google Scholar]
- 105.Morrison SF, Madden CJ, Tupone D. Central neural regulation of brown adipose tissue thermogenesis and energy expenditure. Cell Metab. 2014;19:741–756. doi: 10.1016/j.cmet.2014.02.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Saito M, Okamatsu-Ogura Y, Matsushita M, Watanabe K, Yoneshiro T, et al. High incidence of metabolically active brown adipose tissue in healthy adult humans: effects of cold exposure and adiposity. Diabetes. 2009;58:1526–1531. doi: 10.2337/db09-0530. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.van Marken Lichtenbelt WD, Vanhommerig JW, Smulders NM, Drossaerts JM, Kemerink GJ, et al. Cold-activated brown adipose tissue in healthy men. N Engl J Med. 2009;360:1500–1508. doi: 10.1056/NEJMoa0808718. [DOI] [PubMed] [Google Scholar]
- 108.Inokuma K, Ogura-Okamatsu Y, Toda C, Kimura K, Yamashita H, et al. Uncoupling protein 1 is necessary for norepinephrine-induced glucose utilization in brown adipose tissue. Diabetes. 2005;54:1385–1391. doi: 10.2337/diabetes.54.5.1385. [DOI] [PubMed] [Google Scholar]
- 109.Cannon B, Nedergaard J. Brown adipose tissue: function and physiological significance. Physiol Rev. 2004;84:277–359. doi: 10.1152/physrev.00015.2003. [DOI] [PubMed] [Google Scholar]
- 110.Cypess AM, Weiner LS, Roberts-Toler C, Franquet Elia E, Kessler SH, et al. Activation of human brown adipose tissue by a β3-adrenergic receptor agonist. Cell Metab. 2015;21:33–38. doi: 10.1016/j.cmet.2014.12.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Parysow O, Mollerach AM, Jager V, Racioppi S, San Roman J, et al. Low-dose oral propranolol could reduce brown adipose tissue F-18 FDG uptake in patients undergoing PET scans. Clin Nucl Med. 2007;32:351–357. doi: 10.1097/01.rlu.0000259570.69163.04. [DOI] [PubMed] [Google Scholar]
- 112.Yoneshiro T, Aita S, Matsushita M, Kameya T, Nakada K, et al. Brown adipose tissue, whole-body energy expenditure, and thermogenesis in healthy adult men. Obesity. 2011;19:13–16. doi: 10.1038/oby.2010.105. [DOI] [PubMed] [Google Scholar]
- 113.Yoneshiro T, Aita S, Matsushita M, Okamatsu-Ogura Y, Kameya T, et al. Age-related decrease in cold-activated brown adipose tissue and accumulation of body fat in healthy humans. Obesity. 2011;19:1755–1760. doi: 10.1038/oby.2011.125. [DOI] [PubMed] [Google Scholar]
- 114.Matsushita M, Yoneshiro T, Aita S, Kameya T, Sugie H, et al. Impact of brown adipose tissue on body fatness and glucose metabolism in healthy humans. Int J Obes. 2014;38:812–817. doi: 10.1038/ijo.2013.206. [DOI] [PubMed] [Google Scholar]
- 115.Kajimura S, Saito M. A new era in brown adipose tissue biology: molecular control of brown fat development and energy homeostasis. Annu Rev Physiol. 2014;76:225–249. doi: 10.1146/annurev-physiol-021113-170252. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Yoneshiro T, Aita S, Matsushita M, Kayahara T, Kameya T, et al. Recruited brown adipose tissue as an antiobesity agent in humans. J Clin Invest. 2013;123:3404–3408. doi: 10.1172/JCI67803. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Hanssen MJ, van der Lans AA, Brans B, Hoeks J, Jardon KM, et al. Short-term cold acclimation recruits brown adipose tissue in obese humans. Diabetes. 2016;65:1179–1189. doi: 10.2337/db15-1372. [DOI] [PubMed] [Google Scholar]
- 118.Yoneshiro T, Saito M. Activation and recruitment of brown adipose tissue as anti-obesity regimens in humans. Ann Med. 2015;47:133–141. doi: 10.3109/07853890.2014.911595. [DOI] [PubMed] [Google Scholar]
- 119.Yoneshiro T, Aita S, Kawai Y, Iwanaga T, Saito M. Nonpungent capsaicin analogs (capsinoids) increase energy expenditure through the activation of brown adipose tissue in humans. Am J Clin Nutr. 2012;95:845–850. doi: 10.3945/ajcn.111.018606. [DOI] [PubMed] [Google Scholar]
- 120.Nirengi S, Homma T, Inoue N, Sato H, Yoneshiro T, et al. Assessment of human brown adipose tissue density during daily ingestion of thermogenic capsinoids using near-infrared time-resolved spectroscopy. J Biomed Opt. 2016;21:091305. doi: 10.1117/1.JBO.21.9.091305. [DOI] [PubMed] [Google Scholar]
- 121.Snitker S, Fujishima Y, Shen H, Ott S, Pi-Sunyer X, et al. Effects of novel capsinoid treatment on fatness and energy metabolism in humans: possible pharmacogenetic implications. Am J Clin Nutr. 2009;89:45–50. doi: 10.3945/ajcn.2008.26561. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Zsiboras C, Matics R, Hegyi P, Balasko M, Petervari E, et al. Capsaicin and capsiate could be appropriate agents for treatment of obesity: a meta-analysis of human studies. Crit Rev Food Sci Nutr. 2016 doi: 10.1080/10408398.2016.1262324. [DOI] [PubMed] [Google Scholar]
- 123.Sidossis LS, Porter C, Saraf MK, Borsheim E, Radhakrishnan RS, et al. Browning of subcutaneous white adipose tissue in humans after severe adrenergic stress. Cell Metab. 2015;22:219–227. doi: 10.1016/j.cmet.2015.06.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Oudart H, Groscolas R, Calgari C, Nibbelink M, Leray C, et al. Brown fat thermogenesis in rats fed high-fat diets enriched with n-3 polyunsaturated fatty acids. Int J Obes Relat Metab Disord. 1997;21:955–962. doi: 10.1038/sj.ijo.0800500. [DOI] [PubMed] [Google Scholar]
- 125.Saito M, Yoneshiro T, Matsushita M. Food ingredients as anti-obesity agents. Trends Endocrinol Metab. 2015;26:585–587. doi: 10.1016/j.tem.2015.08.009. [DOI] [PubMed] [Google Scholar]
- 126.Saito M, Yoneshiro T, Matsushita M. Activation and recruitment of brown adipose tissue by cold exposure and food ingredients in humans. Best Pract Res Clin Endocrinol Metab. 2016;30:537–547. doi: 10.1016/j.beem.2016.08.003. [DOI] [PubMed] [Google Scholar]