
Figure 2.
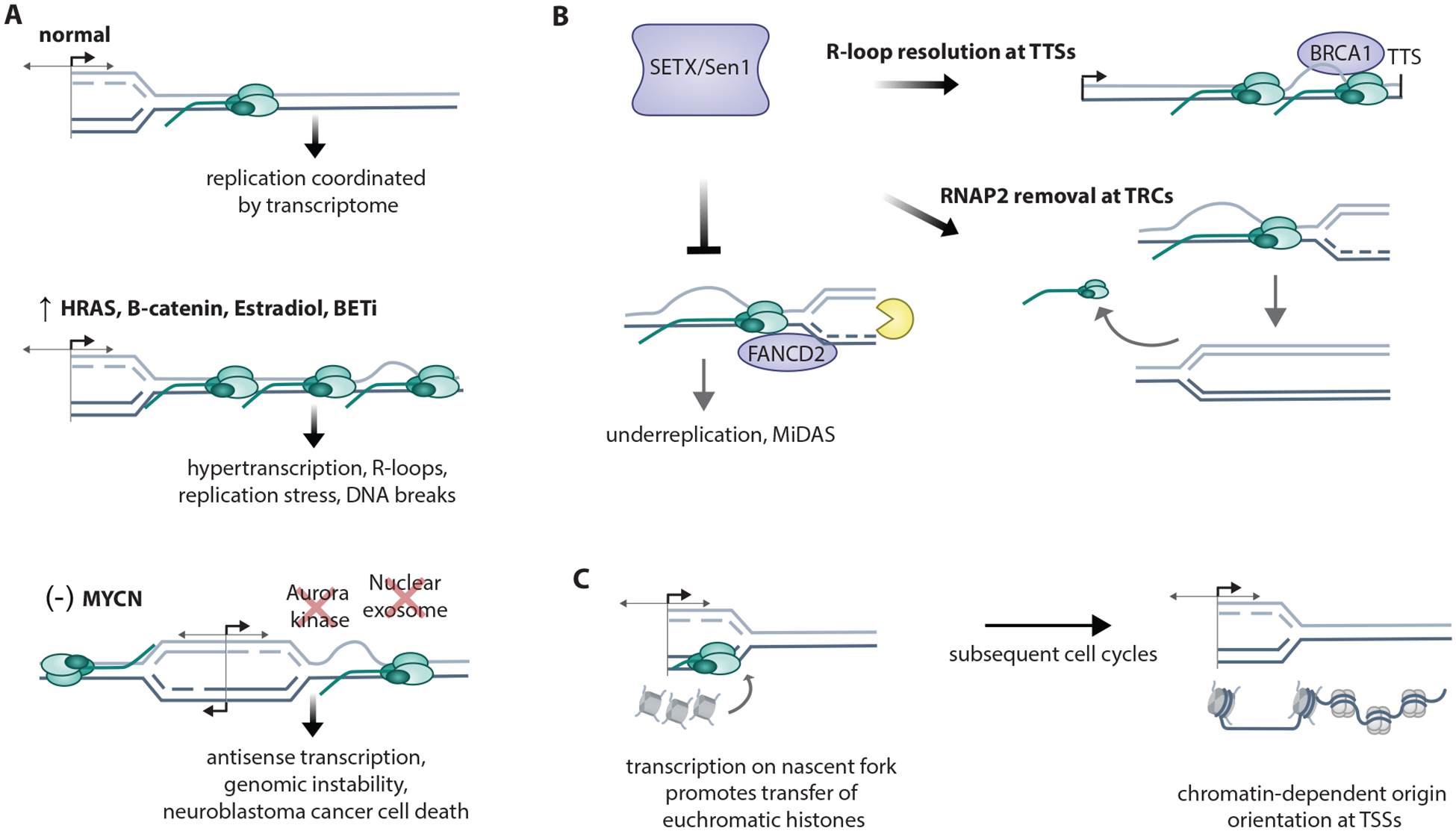
Replication dysregulation causes TRCs
A. Short G1 and premature S-phase entry causes increased usage of canonical origins and over-replication leading to double strand breaks (DSBs) resulting from head-to-tail-fork collisions from mixed G1- and S-phase forks. (Pfander ref) Cyclin E1 overexpression causes intragenic origin firing and TRCs linked to fork collapse and genomic rearrangements (Halazonetis ref), as the result of incomplete MCM redistribution by transcription machinery. In contrast, low-dose HU causes dormant origin firing at transcription termination sites of highly-transcribed genes due to MCM clearance of gene bodies by transcription.
B. Several pathways of replication fork recovery follow TRCs. Replication fork restart by MUS81 fork cleavage leads to RAD52/LIG4/POLD3-dependent fork religation. Replication fork reversal is triggered by MUS81 cleavage, which activates ATR, which limits excessive MUS81 cleavage. Replication fork repriming past R-loop and G4 secondary structures requires PRIMPOL (primase polymerase). Local RNAP removal by ATAD5, RECQ5, or PNUTS resolves TRCs.