
Fig. 3.
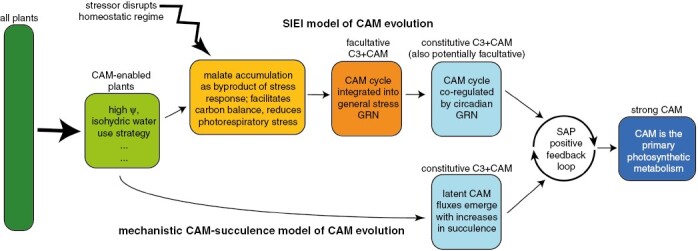
Model of CAM evolution showing two potential trajectories from C3 to strong CAM. Understanding which characteristics of C3 plants facilitate the establishment of a nascent CAM cycle (‘CAM-enabled’ box) remains an open research question; likely candidates include species that maintain high tissue water potentials and operate with a conservative water-use strategy. The upper trajectory follows the SIEI model, hypothesizing that stress induction results in variation in malate accumulation, which is selected for and eventually genetically assimilated, with a full CAM cycle becoming regulated by stress-induced gene regulatory networks (GRNs). Eventually, a CAM cycle also becomes constitutively expressed, potentially via co-regulation with circadian GRNs and/or increases in succulence (see lower trajectory). The lower trajectory follows a ‘mechanistic CAM–succulence model’, whereby all plants have a latent CAM cycle such that increased fluxes may be selected for when expressed in a facilitating anatomical context. Both trajectories result in a plant with a constitutive but weakly expressed CAM cycle with moderate succulence. The evolutionary transition to strong CAM is driven by a ‘synergistic anatomical pleiotropy’, whereby further anatomical changes towards increased succulence positively influence both CAM and water-storage functions, driving convergent evolution of strong-CAM succulent life forms across the tree of life (sensuEdwards, 2019). Abbreviations: Ψ, photosynthetic tissue water potential; GRN, gene regulatory network; SAP, synergistic anatomical pleiotropy.