

Summary
The oocyte-to-embryo transition (OET) is regulated by maternal products stored in the oocyte cytoplasm, independent of transcription. How maternal products are precisely remodeled to dictate the OET remains largely unclear. In this work, we discover the dynamic solubility phase transition of maternal RNAs during Xenopus OET. We have identified 863 maternal transcripts that transition from a soluble state to a detergent-insoluble one after oocyte maturation. These RNAs are enriched in the animal hemisphere and many of them encode key cell cycle regulators. In contrast, 165 transcripts, including nearly all Xenopus germline RNAs and some vegetally localized somatic RNAs, undergo an insoluble-to-soluble phase transition. This phenomenon is conserved in zebrafish. Our results demonstrate that the phase transition of germline RNAs influences their susceptibility to RNA degradation machinery and is mediated by the remodeling of germ plasm. This work thus identifies important remodeling mechanisms that act on RNAs to control vertebrate OET.
Keywords: RNA phase transition, germ plasm, Xvelo1, Bucky ball, oocyte-to-embryo transition, Xenopus, zebrafish
Graphical Abstract

eTOC blurb:
Hwang et al. report that many maternal RNAs can transition between the soluble and insoluble phases during vertebrate oocyte-to-embryo transition (OET). In the case of germline RNAs, they undergo an insoluble-to-soluble phase transition during the OET as a result of the remodeling of germ plasm.
Introduction
The oocyte-to-embryo transition (OET) is one of the most dramatic developmental transitions during which the oocyte and sperm fuse to produce an embryo capable of giving rise to progeny. Prior to the OET, the oocyte accumulates large amounts of maternal products during oogenesis. During the OET, a series of events, including meiotic oocyte maturation, ovulation, fertilization, and zygotic genome activation, occur sequentially, allowing the oocyte to transition into a rapidly growing embryo. Strikingly, these events are precisely regulated by maternal gene products stored in fully-grown oocytes, independent of transcription 1. After decades of extensive investigation, it is still largely unclear how maternal gene products are remodeled during the OET to direct the initiation of embryonic development.
It is well known that numerous remodeling events happen during the OET. These include the remodeling of cellular organelles, as well as macromolecules in the oocyte. One of the best-studied remodeling events during the OET is the remodeling of the endoplasmic reticulum (ER) during oocyte maturation. In most vertebrate species, the ER is closely associated with the germinal vesicle (GV) with some tubular ER being distributed in the cytoplasm of fully-grown oocytes. After germinal vesicle breakdown (GVBD), the morphology and distribution of the ER are changed massively. As a result, a substantial amount of the ER is placed under the plasma membrane, around the future sperm entry site. This unique organization allows rapid release of calcium from the ER upon sperm entry, facilitating egg activation 2-10. In some animal species, ER is pivotal for the asymmetric localization of maternal RNAs, especially some RNAs essential for early embryonic patterning 11-15. In Xenopus, interfering with ER remodeling during the oocyte maturation severely impairs the proper localization of maternal RNAs 16. Interestingly, the proteasome system is remodeled during the OET as well. In mice, proteasomes become highly enriched in the nucleus after the OET 17. In Xenopus, proteasomes are translocated to the animal hemisphere during the OET. This increases the half-life of vegetally localized Dnd1, allowing the accumulation of Dnd1 protein in the embryo to facilitate germline development 18.
A few recent studies demonstrate that at the global level, maternal mRNAs are remodeled during the OET. In humans, many maternal mRNAs are deadenylated first and then re-polyadenylated. During re-adenylation, massive incorporation of non-A residues occurs. Interfering with this remodeling event arrests human embryos at the 1-cell stage 19. In Xenopus, a large amount of maternal RNAs is associated with the ER in fully-grown oocytes. During oocyte maturation, the mRNA-ER association is decreased, leading to the relocation of many maternal RNAs in mature eggs. The decreased mRNA-ER association was observed during mouse oocyte maturation as well 16. In a study to analyze the dynamic structural changes in the 3’UTR of zebrafish maternal mRNAs, Shi et al. detected an opening in the 3’UTR structure immediately after fertilization and a gradual close during cleavage, followed by a reopening after the zygotic genome activation. These structural changes alter the accessibility to specific RNA-binding proteins, influencing the stability of maternal RNAs 20. These findings are exciting and highlight the importance of the remodeling of maternal mRNAs during the OET.
In this study, we report that a subset of maternal mRNA undergoes solubility phase transition during Xenopus and zebrafish OET. Our results reveal that some mRNAs encoding cell cycle regulators become more insoluble during the OET, whereas germline RNAs, which are highly insoluble in the oocyte, are solubilized during the OET. We provide evidence that the solubility of germline RNAs is regulated by Xvelo1/Buc proteins, which form the matrix of the germ plasm 21-24. Moreover, we show that the solubility of germline RNAs influences their susceptibility to RNA degradation machinery. Our work thus uncovers important RNA remodeling mechanisms that occur during vertebrate OET.
Results
Solubility phase transition of maternal RNA during oocyte maturation
We recently investigated RNA-ER association during Xenopus oocyte maturation by fractionation RNA-seq. Our results reveal that the majority of maternal RNAs are distributed in the cytosolic and ER fractions in the oocyte and mature egg. Interestingly, about 10% of RNAs are present in the insoluble pellet fraction, which is resistant to Triton X-100 extraction and can be precipitated by centrifugation at 800 x g 16. While the total amount of RNA in the pellet fraction remains largely unchanged during oocyte maturation, the composition of transcripts in the pellet fraction is dynamically regulated. To simplify our analysis, we combined the cytosolic and ER fractions as the soluble fraction and calculated the percentage of each transcript distributed in the soluble and insoluble (the pellet fraction) fractions. As shown in Fig 1A and S-table 1, while the majority of RNAs are soluble, some RNAs exist in an insoluble form. We detected 233 insoluble transcripts in the oocyte. After oocyte maturation, 29.2% of these transcripts remain insoluble. The remaining 70.8% of transcripts show a decrease in the pellet fraction. Among all transcripts mainly distributed in the soluble fraction in the oocyte, 863 transcripts show a significant increase (above 2-fold) in the insoluble fraction after oocyte maturation. These observations indicate that the solubility of maternal RNA is dynamically regulated during oocyte maturation. We categorized RNAs in the insoluble fraction into three groups, RNAs undergoing an insoluble-to-soluble phase transition (I-S) during oocyte maturation, RNAs going through a soluble-to-insoluble phase transition (S-I), and RNAs that are highly insoluble in both oocytes and mature eggs (I-I).
Fig 1. Solubility phase transition of RNA during oocyte maturation.
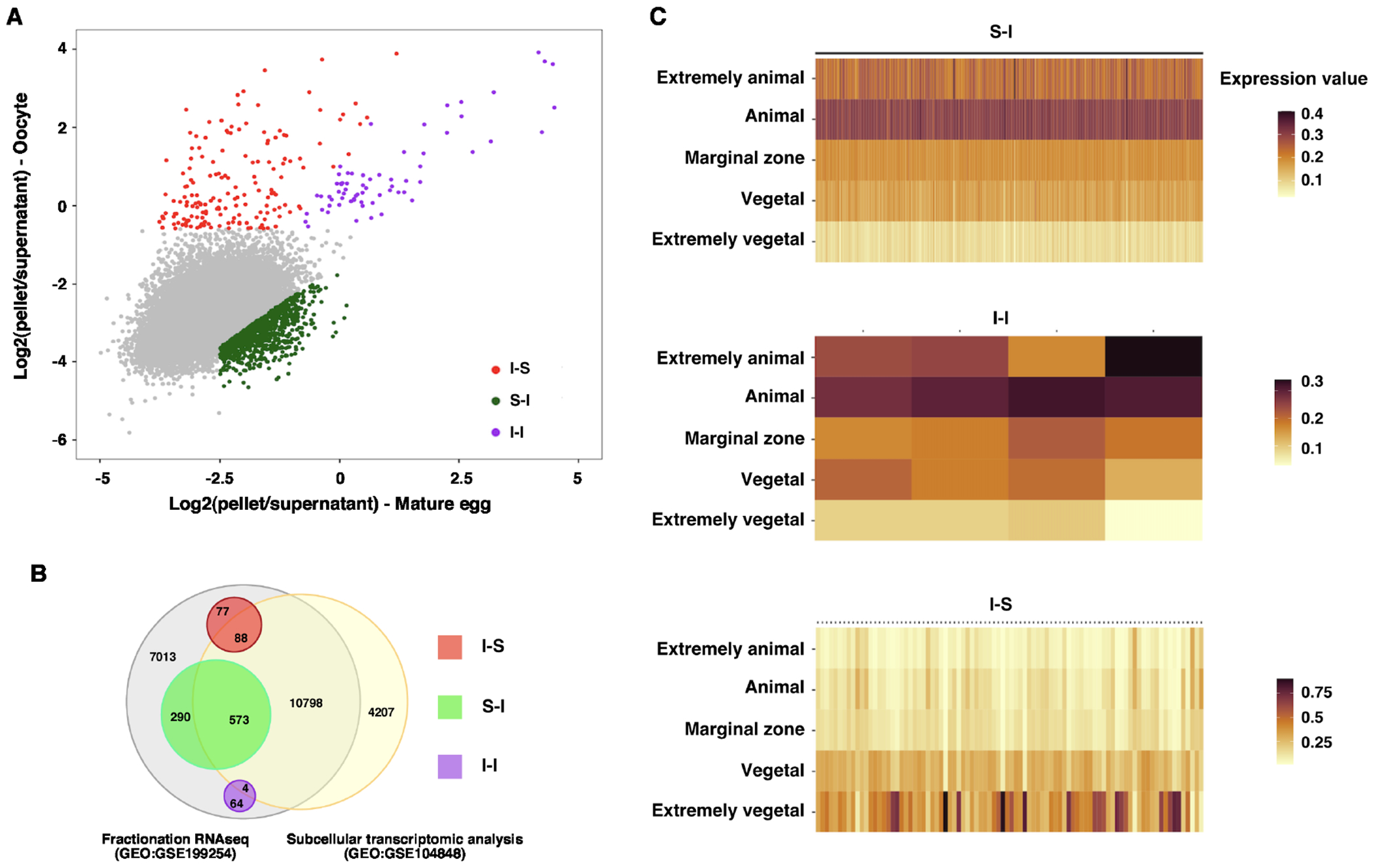
A. Scatter plot shows the pellet to the supernatant ratio in the oocyte (Y-axis) and mature egg (X-axis). I-S transcripts (red) were defined as transcripts enriched by more than 40% in the oocyte pellet fraction and reduced by more than 25% in the pellet fraction after oocyte maturation. I-I transcripts (magenta) were defined as those transcripts enriched by more than 40% in the oocyte pellet fraction, excluding those belonging to the I-S group. S-I transcripts (green) were defined as a more than 2-fold increase in the pellet fraction of mature eggs after oocyte maturation and more than 15% pellet enrichment. B. Venn diagram shows the overlapping relationship between the fractionation RNA-seq (GEO:GSE199254) and subcellular transcriptomic analysis (GEO:GSE104848). C. Heat maps show the distribution of S-I, I-I, and I-S transcripts along the animal-vegetal axis.
In Xenopus, many maternal RNAs are asymmetrically distributed along the animal-vegetal axis 25. To begin understanding the solubility phase transition of maternal RNAs during oocyte maturation, we first determined if RNAs undergoing phase transitions are asymmetrically located along the A/V axis. To this end, we compared our fractionation RNA-seq data to the subcellular transcriptomic analysis by Sindelka et al. 26, in which the distribution of maternal transcripts was analyzed by dissecting the egg along the animal-vegetal axis, followed by RNA-seq. Among 863 S-I RNAs, 573 were detected by Sindelka et al 26 (Fig 1B). Intriguingly, all these RNAs are enriched in the animal hemisphere (Fig 1C and S-table 2). In contrast, I-S RNAs show the opposite pattern. Among 88 I-S RNAs detected by Sindelka et al 26 (Fig 1B and S-table 2), the majority of them are vegetally localized (Fig 1C and S-table 2). These findings suggest that distinct mechanisms operate along the animal-vegetal axis to regulate the solubility phase transition of maternal RNAs during oocyte maturation.
Soluble-to-insoluble phase transition of maternal RNA during oocyte maturation
To better understand the RNA solubility phase transition during oocyte maturation, we first studied S-I RNAs. In the fractionation RNA-seq analysis, S-I RNAs show at least a 2-fold increase in the pellet fraction after oocyte maturation (Fig 2A). We performed a Gene Ontology (GO) analysis and found that S-I RNAs are involved in the mRNA metabolic process, cell division, DNA metabolic process, and other cellular processes important for cell proliferation (Fig 2B). To validate the RNA-seq result, fully-grown oocytes and mature eggs were fractionated into soluble and insoluble fractions, followed by RT-qPCR for the expression of ccna1, wee2.S, hmmr.L, parpbp.L, cep152.L, lig4.L, larp1b.S, exd3.L, pif1.L, sox3.S, sema3d.S, ssx2ip.L, espl1.L, wee2.L, ncapd2.S, fbxo43.L, cdc6.L, ccdc18.L, zbtb12.L, kank1.L, ccnb1.2.L, ncbp1.S, eif2ak3.S, dock7.S, ccnb2.L, dbr1.L, and rad21.L. Indeed, all these S-I RNAs show a significant increase in the insoluble fraction after oocyte maturation (Fig 2C and D), confirming these maternal transcripts indeed go through soluble-to-insoluble phase transition during oocyte maturation.
Fig 2. Soluble-to-insoluble phase transition of RNA during oocyte maturation.
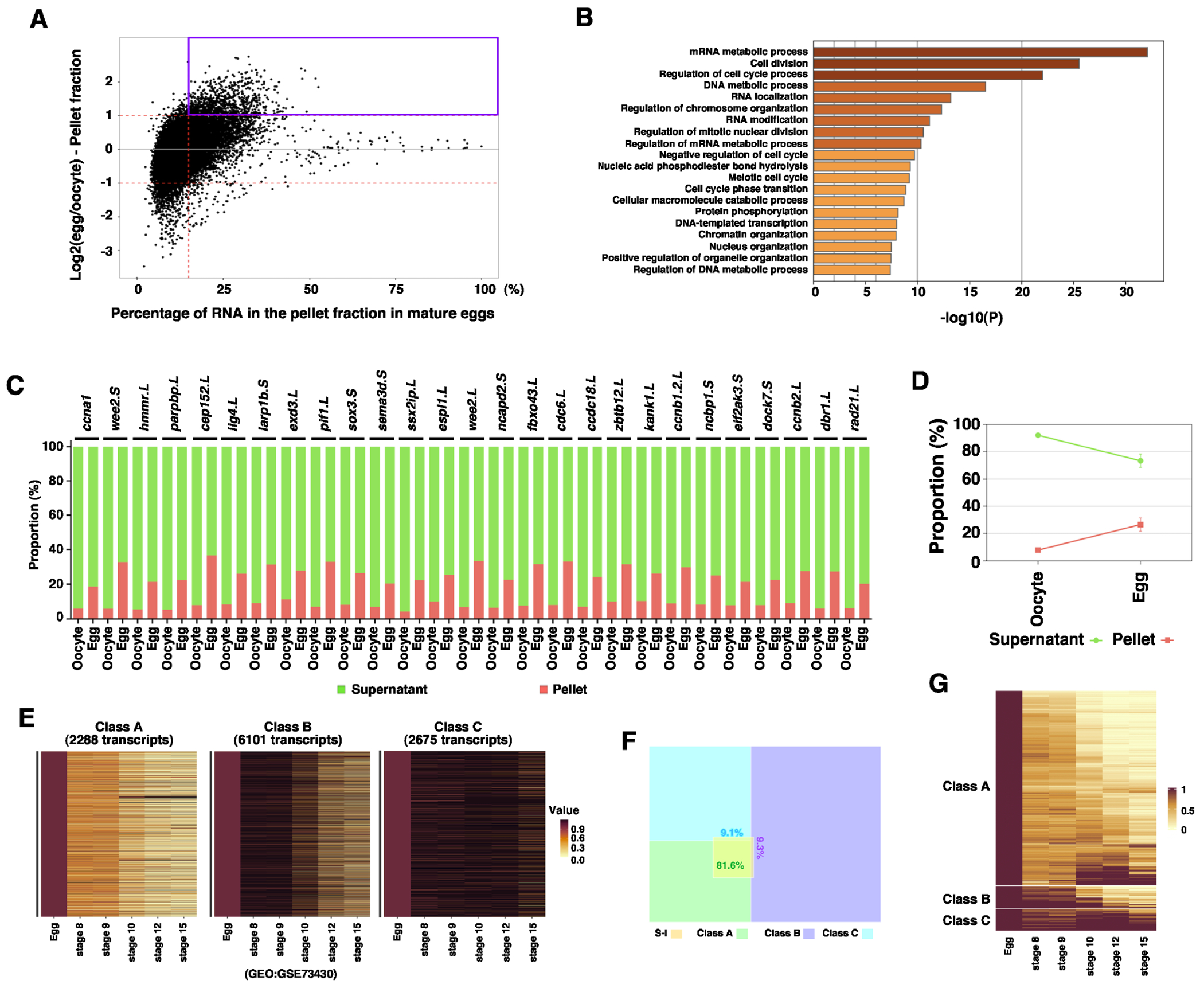
A. MA plot shows the percentage of RNA in the pellet fraction in the mature egg (X-axis) and the ratio between the RNA in the pellet fraction of the egg and that of the oocyte (Y-axis). S-I transcripts are highlighted in the magenta box. B. Gene ontology analysis demonstrates the top biological processes that are enriched among S-I transcripts. C. Fractionation RT-qPCR was performed to validate fractionation RNA-seq results. The percentage distribution of ccna1, wee2.S, hmmr.L, parpbp.L, cep152.L, lig4.L, larp1b.S, exd3.L, pif1.L, sox3.S, sema3d.S, ssx2ip.L, espl1.L, wee2.L, ncapd2.S, fbxo43.L, cdc6.L, ccdc18.L, zbtb12.L, kank1.L, ccnb1.2.L, ncbp1.S, eif2ak3.S, dock7.S, ccnb2.L, dbr1.L, and rad21.L in the supernatant and pellet fractions were calculated. D. The percentage distribution of all markers analyzed in C was combined and plotted into the graph. E. Heat maps show the classification of maternal transcripts based on their degradation during the MZT. Class A transcripts are most rapidly degraded. Class B transcripts are degraded relatively slowly. Class C transcripts are relatively stable during early development. F. Venn diagram shows the majority of S-I transcripts belong to Class A. G. Heatmap shows the expression of S-I RNAs during early embryonic development.
It is well known that after fertilization, Xenopus embryos go through 12 rapid synchronous cell divisions. At the mid-blastula transition (MBT), large-scale transcription happens, followed by slower asynchronous cell divisions 27,28. The lengthening of the cell cycle after the MBT is a consequence of the rapid degradation of maternal cell cycle regulators such as Cyclins during the maternal-to-zygotic transition (MZT) 29,30. Since several cyclin RNAs, including ccna1, ccnb1, ccne1, and ccno undergo soluble-to-insoluble phase transition during oocyte maturation, we set out to determine if the soluble-to-insoluble phase transition correlates with the degradation of maternal RNAs during the MZT. Gene expression profiles of unfertilized Xenopus eggs and embryos at various developmental stages have been analyzed by RNA-seq 31. Using these datasets, we assessed the turnover of maternal mRNAs during the MZT. We divided maternal RNA into three classes. Class A RNAs are rapidly degraded after the MBT. Degradation of Class B RNAs occurs relatively slowly. Class C RNAs are relatively stable (Fig 2 E, S-table 3). Intriguingly, the majority (81%) of S-I RNAs fall into Class A (Fig 2F and G), indicating that the soluble-to-insoluble phase transition of RNAs during oocyte maturation correlates with the turnover of these RNAs during the MZT.
Insoluble-to-soluble phase transition of maternal RNA during oocyte maturation
Next, we investigated RNAs that are enriched in the insoluble fraction in the oocyte. As shown in Fig 3A and S-table 1, 233 I-S transcripts are highly enriched in the pellet fraction in the oocyte. Among these, 165 transcripts undergo an insoluble-to-soluble phase transition during oocyte maturation. We performed GO (Fig 3B) and protein-protein interaction (PPI) analysis (Fig 3C). The results reveal that nearly all Xenopus germline RNAs are I-S transcripts. These include ddx25 32, nanos1 33, and dnd1 34, and genes recently found to be important for PGC development 35. Many germline regulators form a protein-protein interaction network (Fig 3C).
Fig 3. Insoluble-to-soluble phase transition of RNA during oocyte maturation.
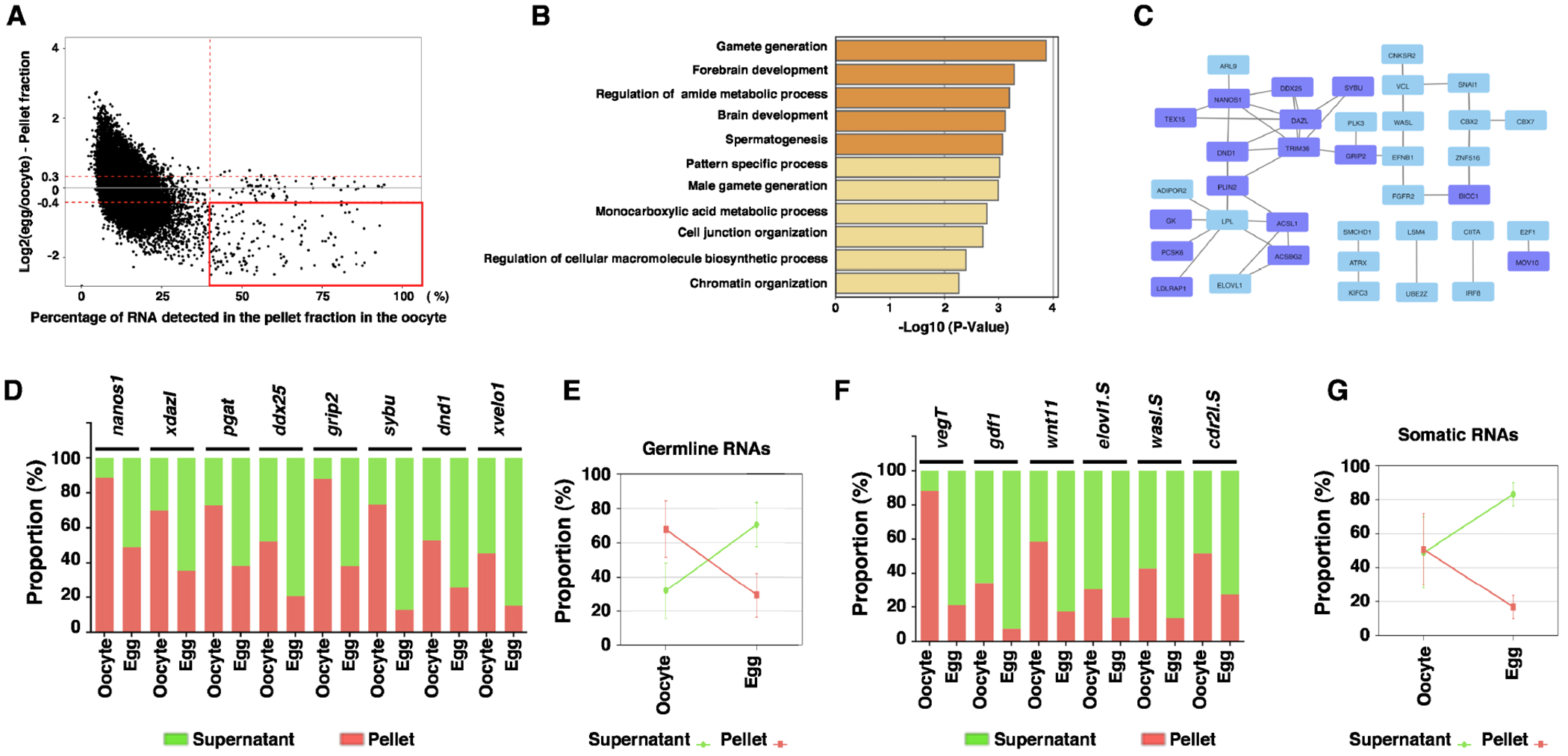
A. MA plot shows the percentage of RNA in the pellet fraction in the oocyte (X-axis) and the ratio between the RNA in the pellet fraction of the egg and that of the oocyte (Y-axis). I-S transcripts are highlighted in the red box. B. Gene ontology analysis demonstrates the top biological processes that are enriched among I-S transcripts. C. Protein-Protein Interaction (PPI) map shows the transcripts selected from a red box in panel A. This PPI shows only proteins interacting with at least one or more other proteins. Purple boxes indicate germline transcripts. D-G. Fractionation RT-qPCR was performed to validate fractionation RNA-seq results. D. The percentage distribution of germline I-S RNAs, including nanos1, xdazl, pgat, ddx25, grip2, sybu, dnd1, and xvelo1 in the supernatant and pellet fractions, were calculated. E. The percentage distribution of all germline RNAs analyzed in D was combined and plotted into the graph. F. The percentage distribution of somatic I-S RNAs, including vegT, gdf1, wnt11, elov11.S, wasl.S, and cdr2l.S in the supernatant and pellet fractions, were calculated. G. The percentage distribution of all somatic I-S RNAs analyzed in F was combined and plotted into the graph.
To validate RNA-seq data experimentally, we fractionated Xenopus oocytes and mature eggs into the soluble and insoluble fractions and analyzed 8 germline I-S RNAs (nanos133, xdazl36, pgat37, ddx2532, grip238, sybu39, dnd134, and xvelo140) and 6 somatic I-S RNAs (vegT, gdf1, wnt11, elovl1.S, wasl.S, and cdr2l.S) by RT-qPCR. Indeed, all the germline transcripts analyzed are enriched in the insoluble fraction in the oocyte. Among these, nanos1 and grip2 RNAs represent the most extreme cases, with as high as 80% of RNAs distributed in the insoluble fraction (Fig 3D and E). After oocyte maturation, the percentage of nanos1, xdazl, pgat, ddx25, grip2, sybu, dnd1, and xvelo1 mRNAs in the insoluble fraction decreases more than two-fold (Fig 3D and E). Similar to germline I-S RNAs, vegT, gdf1, wnt11, elovl1.S, wasl.S, and cdr2l.S are enriched in the insoluble fraction in the oocyte, and are released into the soluble fraction after oocyte maturation (Fig 3F and G). In parallel, we assessed the expression of I-I RNAs by fractionation RT-qPCR. We found rgs2.L, rnu2, thbs1.S, gata6, and bcam.S are enriched in the insoluble fraction in both oocyte and mature egg (S-Fig 1).
Our bioinformatic analysis suggests that the soluble-to-insoluble and insoluble-to-soluble phase transition occurs in the animal and vegetal hemispheres, respectively (Fig 1C). To validate this experimentally, we dissected oocytes and mature eggs into animal and vegetal halves for fractionation RT-qPCR. Indeed, we found the soluble-to-insoluble phase transition occurs in the animal hemisphere (S-Fig 2A), whereas the insoluble-to-soluble phase transition occurs in the vegetal hemisphere (S-Fig 2B). Since ER remodeling during the OET is important for RNA localization 16, we went on to determine if ER remodeling has an impact on RNA solubility phase transition during the OET. To this end, we disrupted the tubular ER network formation during the OET by treating oocytes with cytochalasin B (CB) 16, and performed a fractionation RT-qPCR using control and CB-treated samples. We found that the CB treatment had no effect on RNA solubility phase transition (S-Fig 3), demonstrating that ER remodeling and RNA phase transition are regulated by distinct mechanisms.
We further extended our analysis by determining when germline mRNAs become insolubilized during oogenesis, we collected stage II, III, IV, and VI oocytes, and performed fractionation RT-qPCR. Our results reveal that nanos1, pgat, xdazl, ddx25, sybu, and grip2 are significantly enriched in the insoluble phase as early as in stage II oocytes (S-Fig 4A). In contrast, dnd1 and xvelo1 are initially soluble and gradually move into the insoluble fraction as oogenesis proceeds (S-Fig 4A). The timing of germline RNAs insolubilization correlates with the assembly of these RNAs into the Balbiani body (Bb) or germ plasm (S-Fig 4B and C). As a control, we also assessed I-I RNAs. Our results reveal that thbs1.S, gata6, and rgs2.L are highly enriched in the insoluble fraction throughout the oogenesis. Interestingly, the majority of rnu2 transcript was detected in the insoluble fraction during early oogenesis. In stage IV and VI oocytes, the percentage of rnu2 in the insoluble fraction decreases. By contrast, bcam.S is soluble during early oogenesis, but becomes increasingly insoluble in stage IV and VI oocytes (S-Fig 4D). These results suggest that different RNAs are recruited into insoluble compartments in the oocyte through distinct mechanisms during oogenesis.
Insoluble-to-soluble phase transition of germline RNAs is a consequence of the degradation of Xvelo1
To understand the biological significance of RNA solubility phase transition during oocyte maturation, we investigated the mechanism governing the insoluble-to-soluble phase transition of germline RNAs. As germline RNAs are sequestered in the germ plasm in the oocyte, we hypothesized that the germ plasm is remodeled during oocyte maturation, leading to the phase transition of germline RNAs.
Since Xvelo1 and its zebrafish homolog Buc are essential for the formation of Balbiani body (Bb) and germ plasm, we examined the expression of Xvelo1 during oocyte maturation. Our results reveal that the Xvelo1 protein is highly enriched in the insoluble fraction in the oocyte. Even after immunoprecipitation (IP) enrichment, we still could not detect Xvelo1 protein in the soluble fraction (Fig 4A). After oocyte maturation, the expression level of Xvelo1 is decreased sharply. After fertilization, the level of Xvelo1 is further reduced. We could no longer detect the Xvelo1 protein after the mid-blastula transition (MBT) (Fig 4B). To confirm this finding, we performed immunostaining. In fully grown oocytes, we were able to detect numerous Xvelo1 puncta in the cortical region and deep cytoplasm in the vegetal hemisphere. After germinal vesicle breakdown (GVBD), the number of Xvelo1 puncta is reduced gradually. In mature eggs, we could detect Xvelo1 puncta only in the cortex of the vegetal pole (Fig 4C).
Fig 4. Turnover of Xvelo1 during oocyte maturation results in the solubilization of germline RNAs.
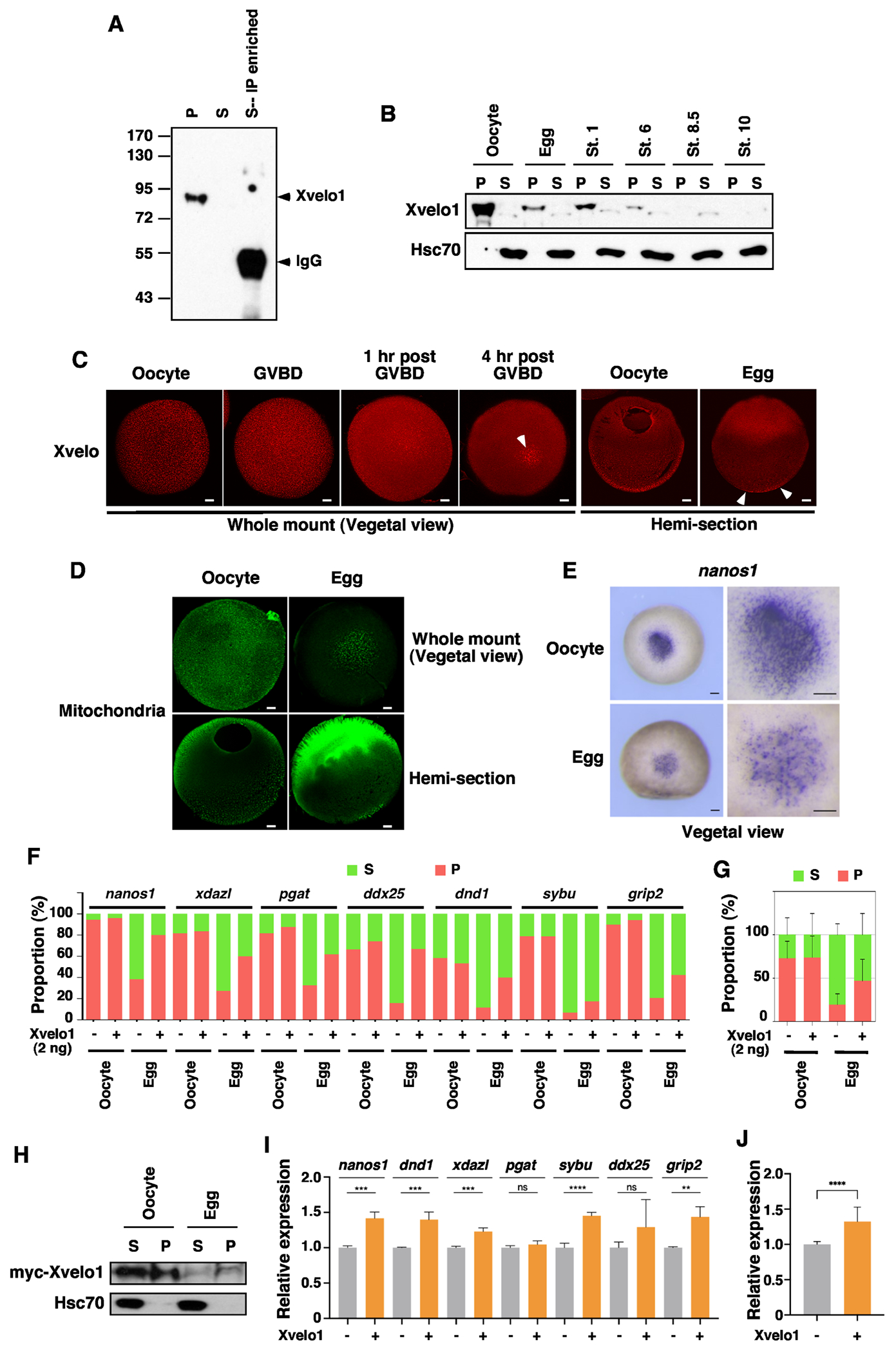
A. Xvelo1 protein is enriched in the insoluble fraction in the oocyte. Oocytes were lysed in NP-40 lysis buffer. After centrifugation, the lysate was separated into the supernatant (S) and pellet (P). Supernatant prepared from 10 oocytes was incubated with an anti-Xvelo1 antibody to enrich Xvelo1 in the soluble fraction. The supernatant, pellet, and IP samples were mixed with SDS sample buffer and subjected to western blotting. B. The expression of Xvelo1 in the oocytes, ovulated eggs, and embryos at stages 1, 6, 8.5, and 10 was analyzed by western blot. C. The expression of Xvelo1 during oocyte maturation was analyzed by IF. White arrowheads point to Xvelo1 remaining in the vegetal pole in mature eggs. The scale bars indicate 100 μm. D. Oocytes and mature eggs of the Dria transgenic frogs, which carry a mitochondria-specific GFP transgene, were stained with an anti-GFP antibody. The scale bars indicate 100 μm. E. Whole mount in situ results show nanosl transcripts are located in punctate aggregates in the vegetal of the oocyte. After oocyte maturation, nanosl transcripts show a diffuse appearance, with only a small number of puncta remaining in the vegetal pole. The scale bars indicate 200 μm. F and G. Overexpression of Xvelo1 prevents solubilization of germline RNAs after oocyte maturation. Oocytes were injected with 2 ng of myc-Xvelo1 RNA, and cultured for 2 days, followed by progesterone treatment. Fractionation RT-qPCR was performed to assess the phase transition of nanos1, xdazl, pgat, ddx25, dnd1, sybu, and grip2 (F). G is the combination of all germline RNAs analyzed in F. H. myc-Xvelo1 RNA injected oocytes were cultured normally or treated with progesterone. Oocytes and mature eggs were fractionated and analyzed for the expression of myc-Xvelo1 and endogenous Hsc70 by western blotting. I and J. Control and Xvelo1 injected oocytes were used to generate embryos via the host-transfer technique. At stage 11.5, embryos derived from control and Xvelo1 injected oocytes were harvested to assess the expression of nanos1, xdazl, pgat, ddx25, dnd1, sybu, and grip2. J is the combination of all markers analyzed in I. Student’s t-tests were performed. ** p<0.01, *** p<0.001, ****p<0.0001.
As Xvelo1 is the key component of the germ plasm matrix 21-24, we went on to determine if the germ plasm is indeed remodeled during oocyte maturation. We assessed mitochondria and nanos1 RNA, which are sequestered in the germ plasm in the oocyte. Consistent with the dynamic changes in Xvelo1 expression, we detected a large amount of mitochondria aggregates in the entire vegetal hemisphere. After oocyte maturation, mitochondria aggregates were detected only in the cortex at the vegetal pole (Fig 4D). Similarly, nanos1 RNAs form puncta in the vegetal pole of fully-grown oocytes. After oocyte maturation, the number of nanos1 puncta is reduced. Diffused nanos1 in situ signals become obvious in the vegetal pole of mature eggs (Fig 4E).
The above results support the idea that degradation of Xvelo1 during oocyte maturation results in the insoluble-to-soluble phase transition of germline RNAs. To test this hypothesis directly, we asked if overexpression of Xvelo1 could prevent the solubilization of germline RNAs during oocyte maturation. Indeed, we found overexpression of Xvelo1 significantly decreased the solubility of germline RNAs in mature eggs (Fig 4F and G). Western blot analysis reveals that a large amount of overexpressed Xvelo1 was present in the pellet fraction in the oocyte. Like endogenous Xvelo1, myc-Xvelo1 was degraded during oocyte maturation. Nonetheless, insoluble myc-Xvelo1 remains detectable in mature eggs (Fig 4H). Using Xvelo1 overexpressed oocytes, we generated embryos via the host-transfer technique 41. Embryos derived from control and Xvelo1 overexpressed oocytes were harvested in the middle of the MZT (stage 11.5) and analyzed for the expression of germline markers. We found that overexpression of Xvelo1 significantly increases the stability of nanos1, dnd1, xdazl, sybu, and grip2 (Fig 4I and J). Taken together, we conclude that Xvelo1 prevents the solubilization of germline RNAs and protects them from degradation during gastrulation. The insoluble-to-soluble phase transition of germline RNAs during oocyte maturation is a consequence of the degradation of Xvelo1 protein.
Insoluble germline RNAs are resistant to RNase A treatment in vitro
In Xenopus oocytes, germline determinants are sequestered in numerous small germ plasm “islands” that are finely dispersed in the vegetal hemisphere. After fertilization, they coalesce into a few large aggregates that are inherited later by primordial germ cells (PGCs). Germline components remaining in the somatic tissue are degraded during the MZT. The above results demonstrate that germline RNAs are more stable during the MZT in embryos derived from Xvelo1 overexpressed Xenopus oocytes (Fig 4I and J). Consistently, when overexpressed in early zebrafish embryos, Buc, which is the homolog of Xvelo1, forms aggregates and prevents clearance of germline RNAs in somatic tissue, converting somatic cells into PGCs 22,42. We speculated that germline RNAs sequestered in the germ plasm may be protected from RNA degradation machinery. Solubilization of germline RNAs during oocyte maturation increases their chance for degradation, facilitating clearance of germline RNAs in the soma during embryonic development.
To test this hypothesis, we carried out an in vitro RNase A treatment assay. We crushed oocytes in lysis buffer and treated the crude lysates with various amounts of RNase A at 37 C° for 5 min. RT-qPCR was performed subsequently to measure the sensitivity of nanos1, ddx25, grip2, actin, hsc70, and h2a RNAs to RNase A. We found that compared to somatic RNAs (actin, hsc70, and h2a), germline RNAs (nanos1, ddx25, and grip2), which are highly insoluble in the oocyte, are less sensitive to RNase A treatment (Fig 5A). We carefully assessed the changes in the sensitivity of RNAs to RNase A during oocyte maturation. Crude lysates from oocytes and mature eggs were treated with 12.5 pg/μl of RNase A for various amounts of time, followed by RT-qPCR for a larger panel of germline RNAs (nanos1, xdazl, pgat, sybu, dnd1, xvelo1, ddx25, and grip2) and somatic RNAs (actin, psma1, odc, gapdh, psme3, ccna1, hsc70, and h2a) (Fig 5B). As expected, somatic mRNAs in oocytes and mature eggs were rapidly degraded by RNase A. The sensitivity of somatic RNAs to RNase A remains unchanged during oocyte maturation. Compared to somatic mRNAs, germline mRNAs were degraded more slowly and to a lesser extent. Interestingly, we found that germline RNAs became more sensitive to the RNase A treatment after oocyte maturation (Fig 5C). To determine if the change in the sensitivity of germline RNAs to RNase A is a result of the solubilization of germline RNAs during oocyte maturation, we fractionated RNase A-treated lysates and measured the sensitivity of soluble and insoluble germline RNAs to RNase A (Fig 5D). As shown in Fig 5E, soluble germline RNAs are highly sensitive to RNase A treatment. We could not detect any difference between oocytes and mature eggs. Insoluble germline RNAs are more resistant to RNase A treatment. Among 8 germline RNAs analyzed, the sensitivity of insoluble nanos1, xdazl, pgat, sybu, xvelo1, and ddx25 to RNase A remains unchanged after oocyte maturation. The sensitivity of insoluble grip2 and dnd1 to RNase A increases slightly. Collectively, the above results suggest that when sequestered in the germ plasm, germline RNAs are less prone to degradation. Solubilization of germline RNAs increases their susceptibility to RNA degradation machinery.
Fig 5. Insoluble germline RNAs are resistant to RNase A in vitro.
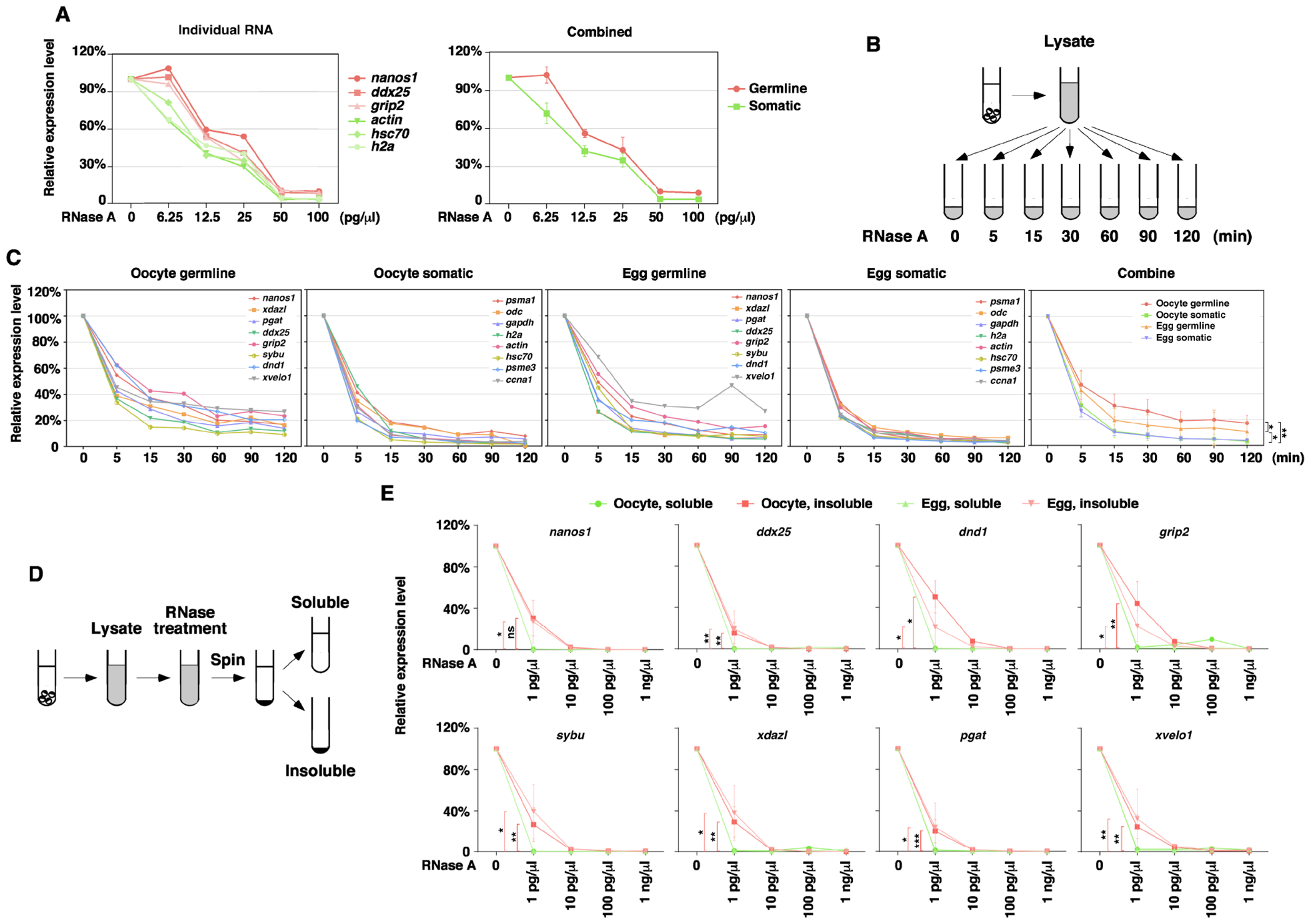
A. Crude oocyte lysate was treated with various doses of RNase A. After RNase A-treatment, RNA was extracted for RT-qPCR. The level of nanos1, ddx25, grip2, actin, hsc70, and h2a was measured. RNA from untreated lysate was set as 100%. B. Schematic diagram shows the procedure for the experiments in panel C. C. Crude oocyte lysate was treated with 12.5 pg/μl RNase A for various amounts of time. Degradation kinetics of germline (nanos1, xdazl, pgat, ddx25, grip2, sybu, dnd1, and xvelo1) and somatic (psma1, odc, gapdh, h2a, actin, hsc70, psme3, and ccna1) RNAs were measured by RT-qPCR. The expression of each germline and somatic RNAs in the oocyte and the mature egg was shown individually. The panel on the right side is the combination of all germline and somatic RNAs in the oocyte and egg. Two-way RM ANOVA tests were performed (oocyte germline vs. oocyte somatic: F (1, 7)=26.8, p=0.0013; egg germline vs. egg somatic: F (1, 7)=6.24, p=0.0411; oocyte germline vs. egg germline: F (1, 7)=8.85, p=0.0207; oocyte somatic vs. egg somatic: F (1, 7)=0.62, p=0.4544), *p < 0.05, **p < 0.01. D. Schematic diagram shows the procedure of experiments in panel E. E. Crude lysate was treated with RNase A, separated into the soluble and insoluble fractions, followed by RT-qPCR for nanos1, xdazl, pgat, ddx25, grip2, sybu, dnd1, and xvelo1. Ratio-paired t-tests at 1 pg/ul were performed. *p < 0.05, **p < 0.01, ***p < 0.001.
Solubility and stability of germline RNAs in zebrafish bucky ball mutant
To definitively determine if the germ plasm, by maintaining germline RNAs in an insoluble state, protects them from the RNA degradation machinery, we took advantage of the zebrafish bucky ball mutant, which lacks Buc protein and is deficient in the formation of Bb and germ plasm. As reported previously 21,22,43, the ER and mitochondria are accumulated in the Bb in wild-type zebrafish oocytes, but are randomly distributed in the entire cytoplasm in bucky ball mutant oocytes (Fig 6A). We performed fractionation and assessed the solubility of several germline RNAs, including nanos3 44, dnd 45, dazl 46, ca15b 47, gra 48, and vasa 49. We found these germline RNAs are enriched in the insoluble fraction in fully-grown wild-type zebrafish oocytes. In contrast, the amount of germline RNAs detected in the insoluble fraction is markedly reduced in bucky ball mutant oocytes (Fig 6B), demonstrating that Buc plays an important role in maintaining zebrafish germline RNAs in an insoluble state. We further compared the expression of germline RNAs in control and bucky ball mutant embryos at the 1-cell stage. We detected a statistically significant decrease in the expression of vasa, gra, and dazl (Fig 6C). Since bucky ball mutant embryos cannot form blastoderm properly, we could not analyze the degradation of germline RNAs during the MZT. Nevertheless, the observation that the expression of a subset of germline RNAs is decreased in bucky ball mutant embryos at the 1-cell stage supports the idea that soluble germline RNAs are more susceptible to RNA degradation machinery.
Fig 6. Buc regulates the solubility and stability of germline RNAs in zebrafish.
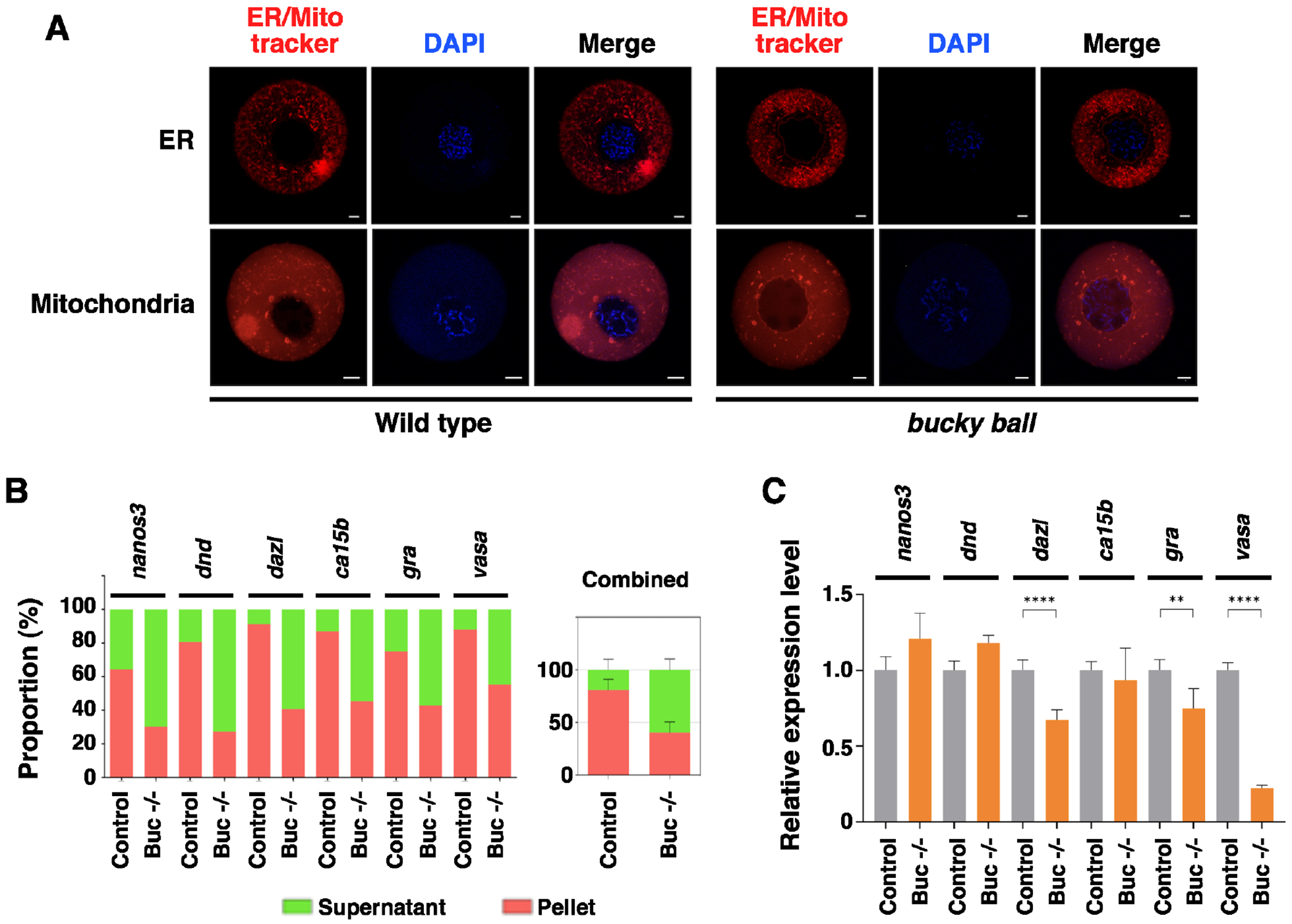
A. Wild type and bucky ball oocytes were stained with ER-tracker and Mito-tracker. Scale bars on the images for ER trackers indicate 20 μm, and those for Mito trackers indicate 10 μm. B. Fractionation RT-qPCR was performed to measure the solubility of nanos3, dnd, dazl, ca15b, gra, and vasa in fully-grown oocytes from the wild-type fish and bucky ball mutants. The right panel is the combination of all these germline RNAs. C. The expression of nanos3, dnd, dazl, ca15b, gra, and vasa in 1-cell stage embryos derived from the wild-type and bucky ball mutant females were assessed by RT-qPCR. Student’s t-tests were performed. ** p<0.01, ****p<0.0001.
Inspired by the above findings, we went on to investigate if the solubility phase transition of mRNA occurs during zebrafish OET. We performed fractionation on oocytes at various stages, ovulated eggs, and pre-MBT embryos, and assessed the solubility of several germline RNAs, together with ccna1, ccnb1, and wee2, which are S-I RNAs in Xenopus (Fig 2). Our results reveal that the solubility of ccna1 and ccnb1 decreases after oocyte maturation. In the case of wee2, although its solubility is not significantly changed during oocyte maturation, it is more enriched in the insoluble fraction after fertilization (Fig 7). Germline RNAs show a rather complex pattern. Compared to somatic RNAs, germline RNAs are more enriched in the insoluble fraction in fully-grown oocytes. Among the six germline RNAs analyzed, dazl, ca15b, gra, and vasa are already in the insoluble fraction in stage I oocytes. Recruitment of nanos3 and dnd to the insoluble fraction occurs gradually during oogenesis. We did not detect any significant changes in the solubility of germline RNAs during oocyte maturation. Instead, we found that the solubility of dazl, ca15b, nanos3, and dnd increases gradually during pre-MBT stages (Fig 7). Thus, similar to what happens in Xenopus, some cell cycle regulators become increasingly insoluble during zebrafish oocyte maturation and early development. The insoluble-to-soluble phase transition does occur in some zebrafish germline RNAs, albeit during cleavage and blastula stages. While some species-specific differences clearly exist, the above results demonstrate that the phase transition of maternal mRNA during the OET is evolutionarily conserved.
Fig 7. RNA phase transition during zebrafish OET.
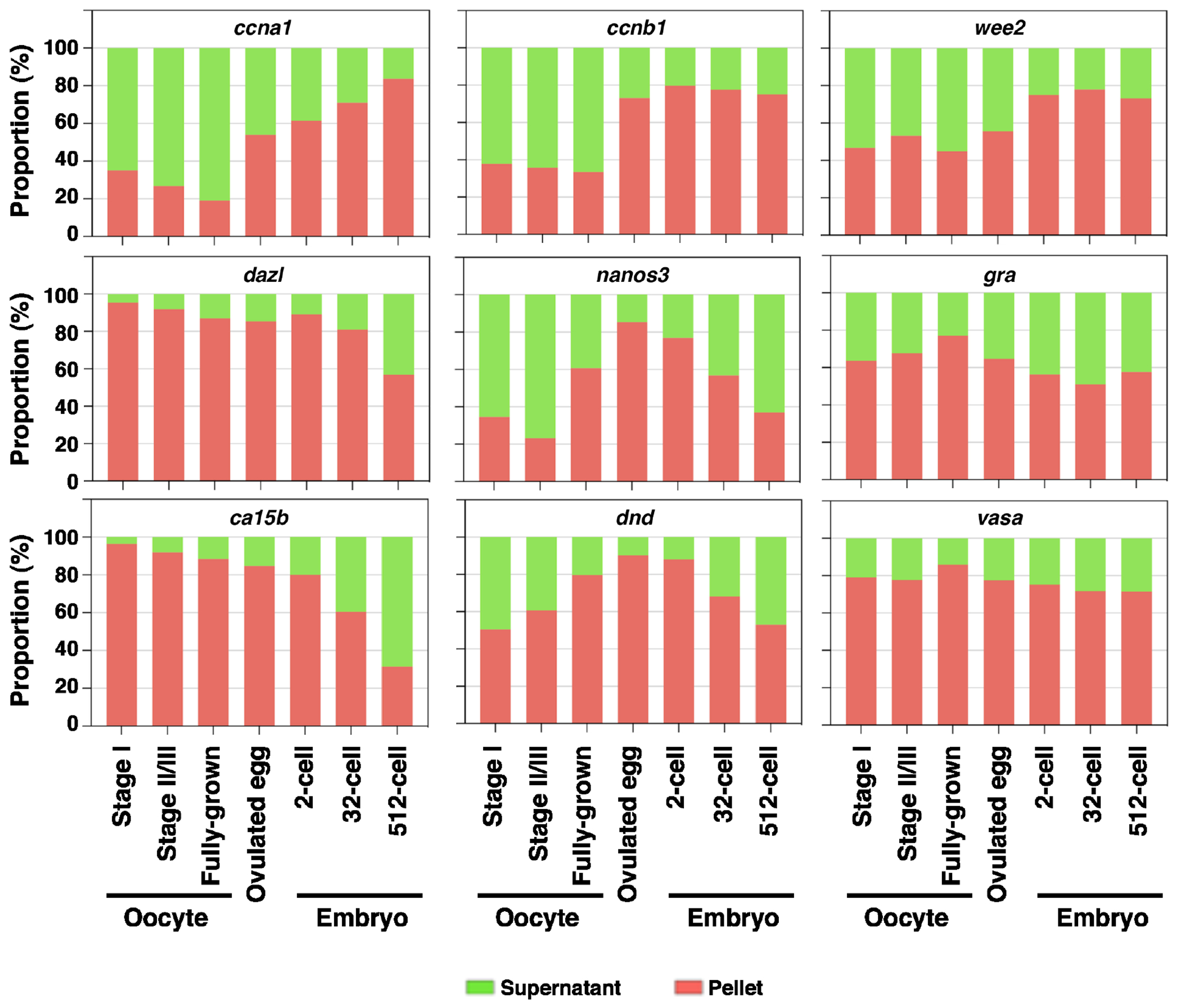
Fractionation RT-qPCR was performed to measure the percentage distribution of ccna1, ccnb1, wee2, dazl, nanos3, gra, ca15b, dnd, and vasa in stage I, stage I/II, and fully-grown oocytes, ovulated eggs, and embryos at 2-, 32-, and 512-cell stages.
Discussion
The OET is one of the most dramatic developmental transitions during which the oocyte undergoes meiotic maturation, fuses with the sperm to form the zygote, and ultimately activates the zygotic genome to direct embryonic development. These events occur in the absence of transcription and are regulated precisely by maternal products that are synthesized and stored in the oocyte during oogenesis. Currently, it remains largely unclear how maternal products are remodeled during the OET to orchestrate the beginning of embryogenesis.
Here we report the solubility phase transition of maternal RNA during the OET, an important remodeling event that appears to be evolutionarily conserved. We found that during Xenopus oocyte maturation, a group of maternal RNAs become increasingly resistant to detergent extraction. The majority of these RNAs are asymmetrically localized in the animal hemisphere. Many of them encode cell cycle regulators that are rapidly degraded during the MZT. We tested some of these maternal genes in zebrafish. Indeed, we found that ccna1 and ccnb1, which encode cyclin proteins, undergo a soluble-to-insoluble phase transition. Like in the Xenopus oocyte, zebrafish ccnb1 is localized in the animal pole 46, suggesting that the soluble-to-insoluble phase transition of maternal mRNA is regulated by an evolutionarily conserved mechanism. In contrast to these S-I RNAs, many vegetally localized RNAs are highly insoluble in Xenopus oocytes, but become solubilized after oocyte maturation. Intriguingly, a subset of I-S RNAs encode proteins essential for Xenopus germline development. Our results reveal that some zebrafish germline RNAs are I-S transcripts as well. Interestingly, we noticed two differences between Xenopus and zebrafish. First of all, the solubility phase transition of zebrafish germline RNAs occurs after fertilization, not during oocyte maturation. Secondly, some zebrafish germline RNAs, including vasa and gra, remain insoluble even at the MBT. Thus, while the solubility phase transition of maternal RNA happens in both Xenopus and zebrafish, some species-specific differences clearly exist.
Our results reveal that the transition of Xenopus germline RNAs into the insoluble phase during early oogenesis is caused by the recruitment of germline RNAs into the germ plasm. Likewise, the solubilization of germline RNAs is a consequence of the remodeling of the germ plasm during the OET. In support of this idea, we found that Xvelo1/Buc, the matrix protein of the germ plasm 21-24, is essential for maintaining germline RNAs in an insoluble state. In zebrafish bucky ball mutant oocytes, which cannot assemble Bb/germ plasm, germline RNAs become much more soluble. During Xenopus oocyte maturation, Xvelo1 is markedly down-regulated, leading to the solubilization of germline RNAs. We found the solubilization of germline RNAs can be inhibited by overexpression of Xvelo1 during Xenopus oocyte maturation. Interestingly, the solubilization of zebrafish germline RNAs occurs after fertilization. This observation is consistent with the finding that down-regulation of Buc occurs during zebrafish early embryonic development. In a study by Riemer et al., abundant Buc was detected in the entire blastodisc after fertilization. Buc declines rapidly during cleavage. By the 128-cell and dome stage, only four Buc protein aggregates are maintained in the embryo 50. This would explain why the timing of the solubilization of germline RNAs is different in Xenopus and zebrafish.
Dynamic regulation of RNA solubility plays an important role in translational regulation in many contexts. It has been reported that amyloid-like aggregation of the RNA-binding protein Rim4 during yeast gametogenesis is important for it to repress the translation of its targets 51. In zebrafish, maternal pou5f3 RNAs are assembled into RNA granules. These granules are solid-like in the oocyte, but become more liquid-like during early embryonic development, leading to translational activation of pou5f3 RNAs in early embryos 52. In neurons, many mRNAs are assembled into neuronal granules and maintained in a translationally repressed state during their transportation. Upon arrival at their final destination, disassembly of these mRNAs from granules results in translational activation of these mRNAs 53. To determine if RNA solubility phase transition during the OET plays a general role in translational regulation, we examined the expression level of proteins encoded by S-I, I-S, and I-I RNAs during Xenopus OET, using the proteomic data published recently by Peuchen et al 54. We failed to detect any obvious correlation between RNA solubility phase transition and changes in the expression of corresponding proteins on a global level (S-Fig 5). It seems unlikely that RNA solubility phase transition serves as a general mechanism to regulate the global translation of maternal RNAs during the OET.
In the case of germline RNAs, a large body of studies has demonstrated that germ plasm functions as the cargo for the vegetal transportation of germline RNAs. Hence insolubilization of germline RNAs during early oogenesis, i.e., recruitment of germline RNAs into the Bb or germ plasm, is essential for the asymmetric localization of germline RNAs 55-59. What is the function of the solubility phase transition of germline RNAs during the OET? Our results presented here demonstrate that the phase transition of germline RNAs influences their susceptibility to RNA degradation machinery. We show that insoluble germline RNAs are more resistant to RNase A treatment in vitro. Germline RNAs are more stable in Xenopus embryos that are derived from Xvelo1 overexpressed oocytes. In zebrafish bucky ball mutants, where the solubility of germline RNAs is increased, a subset of germline RNAs is down-regulated. Thus, maintaining germline RNAs in an insoluble form ensures that germline RNAs are stored in a protective environment. It is well-known that after fertilization, only a fraction of germline RNAs coalesce into large aggregates and are inherited by PGCs. The remaining ones are degraded in somatic cells. Interfering with somatic clearance of germline RNAs impairs the segregation of the germline from the soma. It has been reported that overexpression of Buc prevents the degradation of germline RNAs in somatic tissue, converting somatic cells into PGCs 22,42. Based on the literature and results presented here, we thus propose that the solubilization of germline RNAs during the OET, by increasing their accessibility to RNA degradation machinery, facilitates the clearance of germline RNAs in the soma.
It is worth mentioning that the solubility phase transition during the OET influences RNAs differently. Insolubilization of germline RNAs by sequestering them in the germ plasm can facilitate their vegetal transportation during early oogenesis 55-59 and protect them from RNA degradation machinery (Fig 4, 5, and 6). Different from germline RNAs, the soluble-to-insoluble phase transition of S-I RNAs correlates with their degradation during the MZT. The majority of S-I RNAs are rapidly degraded after the zygotic genome activation (Fig 2). It will be of great interest to determine if insolubilization of S-I RNAs during the OET potentiates their degradation during the MZT.
In summary, this work demonstrates that many RNAs can transition between the soluble and insoluble phases under physiological conditions. It raises the striking possibility that as an imporant post-transcriptional regulatory mechanism, precisely regulated RNA solubility phase transition may play fundamentally important roles in a wide variety of biological processes.
Limitations of the study
Our finding that a fairly large number of maternal RNAs are insolubilized during oogenesis (I-S and I-I transcripts) and the OET (S-I transcripts) is fundamentally important. It argues for the existence of remodeling mechanisms that act on RNAs to regulate oogenesis and the OET. RNAs become insoluble likely because they are incorporated into some insoluble protein condensates. The identification of the I-I, I-S, and S-I RNAs suggests that there are several types of protein condensates, which are presumably involved in distinct post-transcriptional regulatory mechanisms important for the oogenesis and OET. In this paper, we have provided a detailed mechanistic explanation for the insoluble-to-soluble phase transition of germline RNAs, representing the first step toward understanding these important regulatory mechanisms. Apart from germline RNAs, which represent only a small portion of I-S transcripts, we do not know how other RNA phase transition events are regulated during the OET. Our results reveal that the soluble-to-insoluble and insoluble-to-soluble phase transition occurs in distinct compartments during Xenopus OET. Vegetally localized Xvelo1 cannot regulate all these RNA phase transition events. Clearly, further work is needed to identify other protein condensates responsible for the RNA phase transition during the OET.
STAR Methods
Resource Availability
Lead contact
Further information and requests for resources and reagents should be directed to and will be fulfilled by the lead contact, Jing Yang (yangj@illinois.edu).
Materials availability
This study did not generate any new unique reagents.
Data and code availability
RNA sequencing data analyzed in this study have been deposited to the Gene Expression Omnibus (GEO) with the dataset identifier GSE199254.
This paper does not report the original code.
Any additional information required to reanalyze the data reported in this paper is available from the lead contact upon request.
Experimental Model and Study Participant Details
Xenopus laevis
Xenopus procedures were approved by the University of Illinois at Urbana-Champaign Institutional Animal Care and Use Committee (IACUC) under animal protocol #20125 and performed in accordance with the recommendations of the Guide for the Care and Use of Laboratory Animals of the National Institutes of Health. All experimental animals were housed in recirculating water systems with adjusted multiple parameters, including 18 °C temperature and 7.2 pH. The Xenopus facility was controlled by an automatic 12-hour light and dark cycle environment. Oocytes were collected from ovarian tissues by manual defolliculation or collagenase treatment and cultured in the oocyte culture medium (OCM) 60. In order to induce oocyte meiotic maturation, stage VI oocytes were cultured in the OCM containing 2 μM progesterone and incubated at 18 °C overnight. Different stages of oocytes and mature eggs were collected for further analysis. For microinjection, xvelo1 mRNAs were injected into the vegetal hemisphere of the oocyte using a Narishige IM300 microinjector, and then the oocyte was either cultured in the OCM or treated with progesterone. To analyze RNA phase transition along the animal-vegetal axis, we dissected oocytes/mature eggs into the animal and vegetal halves using Gastromaster. Animal and vegetal halves were used for cell fractionation and RT-qPCR.
Danio rerio
Zebrafish procedures were approved by the University of Illinois at Urbana-Champaign Institutional Animal Care and Use Committee (IACUC) under animal protocol #21179 and performed in accordance with the recommendations of the Guide for the Care and Use of Laboratory Animals of the National Institutes of Health. All experimental animals were housed in recirculating water systems with adjusted multiple parameters, including 28 °C temperature and 7.4 pH. The zebrafish facility was controlled by an automatic 14-hour light and 10-hour dark cycle environment. The Tübingen strain was purchased from ZIRC (Zebrafish International Resource Center), and Buckyball mutant zebrafish were received from Dr. Mullins’ lab at the University of Pennsylvania. Zebrafish oocytes were obtained as described 61. Briefly, ovarian tissues were dissected and dissociated by 15 mg/ml collagenase treatment for 30 min at room temperature (RT). Oocytes at each stage were sorted and collected manually for further experiments.
Method Details
Cell Fractionation
Fractionation was performed as described 16 with slight modification. Briefly, Xenopus oocytes and mature eggs were rinsed with ice-cold washing buffer (150 mM KOAc, 2.5 mM MgOAc2, and 20 mM K-HEPES pH 7.4). To extract soluble fraction, four oocytes or eggs were homogenized in 160 μl of ice-cold extraction buffer (150 mM KOAc, 2.5 mM MgOAc2, 20 mM K-HEPES pH 7.4, 2 mM DTT, 1 mM PSMF, 50 μg/ml cycloheximide, and 200 units/ml RNase inhibitor) containing 0.5 % Triton X-100 and centrifuged at 800 x g for 5 min. After centrifugation, supernatants were transferred to new tubes and centrifuged again at 10,000 x g for 10 min to remove contaminating organelles and cell debris. Pellets were resuspended in 160 μl of ice-cold extraction buffer with 0.5 % Triton X-100. The resulting supernatants (soluble fractions) and resuspended pellet solution (insoluble fractions) were used for further analysis. For the fractionation of zebrafish samples, five zebrafish oocytes, ovulated eggs, and embryos were pooled and homogenized in 100 μl of ice-cold extraction buffer containing 0.5 % Triton X-100 and centrifuged at 10,000 x g for 10 min. After centrifugation, supernatants were transferred to new tubes. Pellets were resuspended with 100 μl of ice-cold extraction buffer containing 0.5 % Triton X-100. All centrifugations were performed in a refrigerated centrifuge at 4 °C.
Whole-mount immunofluorescence
Xenopus stage VI and mature eggs were fixed in Dent’s fixative (80 % of methanol and 20 % DMSO) at −20 °C overnight. After fixation, samples were washed with 100 % methanol three times for 5 min each time and stored in 100 % methanol at −20 °C until the staining procedure was initiated. For whole-mount immunofluorescence, samples were rehydrated in serial dilution of methanol with TBST (TBS with 0.1% Triton X-100) for 5 min each at room temperature. Oocytes and eggs were bisected by a razor blade to obtain hemi-sectioned or vegetal hemisphere samples. After bisection, samples were washed in TBST once and incubated in a blocking buffer (0.15 % Triton X-100, 2 % BSA in TBS) containing 10 % normal serum from the same host as the secondary antibody for an hour at RT. subsequently, samples were incubated in a blocking buffer with the primary antibodies (1:100) overnight at 4 °C. Samples were washed with TBST six times for 30 min each, incubated in a blocking buffer with the secondary antibodies (1:500) overnight at 4 °C, and washed with TBST six times for 30 min each. Stained samples were washed with 100% methanol twice for 10 min each and then added in BABB (1:2 ratio of benzyl alcohol: benzyl benzoate) to be cleared. Images were acquired by Nikon A1Rsi confocal microscope.
In vitro RNase A assay
For time-dependent RNase A assay, 40 oocytes and 40 mature eggs were harvested and homogenized in 800 μl of lysis buffer. 100 μl of the lysates was saved as ‘no RNase A′. The remaining lysates were transferred into six tubes, each containing 100 μl, and treated with RNase A (12.5 pg/μl; final concentration) at 37 °C for 5, 15, 30, 60, 90, and 120 min. After the RNase A treatment, RNAs were extracted for RT-qPCR. For fractionation RNase A assay, 20 oocytes and 20 mature eggs were harvested and homogenized in 800 μl of extraction buffer (150 mM KOAc, 2.5 mM MgOAc2, 20 mM K-HEPES pH 7.4, 2 mM DTT, 1 mM PSMF, 50 μg/ml cycloheximide, and 200 units/ml RNase inhibitor) containing 0.5 % Triton X-100. 160 μl of the lysates was saved as ‘no RNase A’. The remaining lysates were transferred into 4 tubes (160 μl each) and incubated in different concentrations (1 pg/μl, 10 pg/μl, 100 pg/μl, and 1 ng/μl) of RNase A at 37 °C for 2 hr. After incubation with RNase A, samples were fractionated into soluble and insoluble fractions by the above protocol. RNAs were extracted using TRIzol reagent according to the manufacturer’s instructions.
In vitro transcription
Xvelo1 mRNAs were synthesized from 2 μg plasmid templates using the mMESSAGE mMACHINE Kit for SP6. All probes used in this study were synthesized from 2 μg plasmid templates using T3 RNA polymerase. To be incorporated Digoxigenin (DIG) UTPs into the probes, 1 μl of DIG RNA labeling mix was added to the in vitro transcription reaction solution. Dig-labeled probes were detected by anti-Digoxigenin-AP Fab fragments/BM-purple staining.
Xenopus in situ hybridization
Oocytes at various stages were fixed with MEMFA (0.1 M MOPS pH 7.4, 2 mM EGTA, 1 mM MgSO4, and 3.7% formaldehyde solution) for an hour at RT, washed with PBS twice, and dehydrated in methanol. Dehydrated samples were stored in 100 % methanol at −20°C. For in situ hybridization, all samples were rehydrated in serial dilution of methanol with PBSW (PBS with 0.1 % Tween-20) for 5 min each at RT. Stage III, IV, V, and VI oocytes were hemi-sectioned by a razor blade. Once all stage II oocytes and bisected mid and late-stage oocytes were prepared, in situ hybridization was performed as described 62.
RNA Extraction and Quantitative RT-PCR
Total RNAs were extracted from the soluble and insoluble fraction of oocytes, eggs, or embryos using TRIzol reagent in accordance with the manufacturer’s instructions. Reverse transcription and real-time PCR were performed according to standard protocols to analyze the expression level of mRNAs. Ct values were acquired by Applied Biosystems QuantStudio 3 Real-Time PCR System. All primers used are listed in the supplementary table.
Immunoprecipitation and Western Blots
For immunoprecipitation, fully-grown oocytes were homogenized in 0.5 % NP-40 lysis buffer (50 mM Tris pH 7.6, 125mM NaCl, 1 mM EDTA, 0.5 % NP-40) with protease inhibitor cocktails and centrifuged at 20,000 x g for 10 min. Supernatants were transferred to new tubes. Xvelo1 antibodies were added to the supernatant and incubated overnight at 4 °C. Subsequently, protein G-coupled agarose beads were added to precipitate Xvelo1 proteins. After washing the beads with lysis buffer three times (5 min each), proteins were eluted in SDS sample buffer by boiling for 5 min at 100°C. For western blots, fractionated samples and whole oocyte/egg lysates were mixed with 2 x SDS sample buffer, boiled for 5 min at 100°C, separated on SDS-PAGE, and transferred to PVDF membranes for western blotting according to the standard protocol for western blots.
Host transfer
Host transfer was performed as described 41. Briefly, manually defolliculated oocytes were vegetally injected with 1ng of xvelo1 mRNAs and cultured at 18 °C for 24 hr. Then, control and injected oocytes were treated with 2 μM of progesterone to induce oocyte meiotic maturation. In parallel, host female frogs were prepared by administrating hCG (human chorionic gonadotropin). After oocyte maturation, matured eggs were stained with vital dyes to distinguish between control and Xvelo1 overexpression samples and transferred into host female frogs. Finally, transferred eggs were recovered and fertilized with sperm. Embryos derived from transferred eggs were harvested at stage 11.5 for gene expression analysis.
Immuno-fluorescence in situ hybridization (Immuno-FISH)
Stage I and II Xenopus oocytes were used. Sample preparation is mentioned above in Xenopus in situ hybridization section. After hybridization with fluorescence ddx25-546 probe, oocytes were washed with 2xSSC (20xSSC pH 7.0; 175.3 g NaCl, 88.2 g sodium citrate) containing 0.1% CHAPS twice for 30 min each at RT, 0.2xSSC containing 0.1% CHAPS twice for 30 min each at 60 °C, and TBST (TBS with 0.1% Triton X-100) for 5 min at RT. For whole-mount immunofluorescence, the samples were incubated in a blocking buffer (0.15 % Triton X-100, 2 % BSA in TBS) containing 10 % normal donkey serum for an hour at RT. Then, the oocytes were incubated in a blocking buffer with the primary antibodies (1:200) overnight at 4 °C and washed with TBST three times for 10 min each. After that, the samples were incubated in a TBST with the secondary antibodies (1:500) for an hour at RT and washed with TBST three times for 10 min each. All stained samples were dehydrated with 100 % methanol and mounted with BABB. All images were acquired using Nikon A1Rsi confocal microscope.
Treatment of cytochalasin B
Defolliculated Xenopus oocytes were pretreated with 25 μg/ml of cytochalasin B to inhibit F-actin polymerization at RT for 1 hr. Then pretreated oocytes were cultured in the OCM containing 2 μM progesterone and incubated at 18 °C overnight. After that, oocytes and eggs were harvested for cell fractionation. DMSO was used as a control.
Mitochondria and endoplasmic reticulum (ER) staining of zebrafish oocytes
Mitochondria and ER staining were performed as described 61 with slight modification. Zebrafish stage I oocytes were isolated from the ovarian tissues of the wild-type and Buc mutant zebrafish according to the protocol mentioned above. Stage I oocytes were stained with 0.5 μM of MitoTracker and 0.5 μM of ER tracker each in staining buffer (PBS with 0.1 % BSA) at RT for 30 min. Then, samples were washed with staining buffer three times for 10 min each at RT. After washing, samples were stained with 0.5 μg/ml of Hoechst 33342 in the staining buffer for 10 min at RT and then mounted with staining buffer in glass concavity slides. All images were acquired using Nikon A1Rsi confocal microscope.
Analysis of RNA-Sequencing
All RNA-seq datasets used in this study were listed in the Key resources table. The fractionation RNA-seq was analyzed as described 16 with slight modification. Briefly, all analyses were performed using abundant transcripts (17,811) and a proportion scale according to the previous research analysis 16. The soluble fraction is the sum of the cytosolic and ER fractions, and the pellet was considered insoluble. These 17,811 transcripts were classified into three groups below.
Key resources table
| REAGENT or RESOURCE | SOURCE | IDENTIFIER |
|---|---|---|
| Antibodies | ||
| Rabbit anti-Xvelo | (Boke et al., 2016) 24 | N/A |
| Goat anti-GFP | Rockland | Cat. 600-101-215; RRID: AB_218182 |
| Mouse anti-HSC70 | Santa Cruz | Cat. sc-7298; RRID: AB_627761 |
| Mouse anti-c-Myc | Thermo Fisher | Cat. 13-2500; RRID: AB_2533008 |
| Secondary anti-rabbit-HRP | GE Healthcare | Cat. NA934V |
| Secondary anti-mouse-HRP | GE Healthcare | Cat. NA931V |
| Alexa Fluor™ donkey anti-rabbit-594 | Thermo Fisher | Cat. A-21207; RRID: AB_141637 |
| Alexa Fluor™ donkey anti-rabbit-488 | Thermo Fisher | Cat. A-21206; RRID: AB_2535792 |
| Alexa Fluor™ donkey anti-goat-488 | Thermo Fisher | Cat. A-11055; RRID: AB_2534102 |
| Chemicals, peptides, and recombinant proteins | ||
| RiboLock RNase Inhibitor | Thermo Fisher | Cat. EO0381 |
| TRIzol reagent | Ambion | Cat. 15596018 |
| 2x SYBR Green qPCR Master Mix | Bimake | Cat. B21203 |
| MitoTracker™ Red CMXRos | Thermo Fisher | Cat. M7512 |
| ER tracker™ Red | Thermo Fisher | Cat. E34250 |
| BM Purple AP substrate | Roche | Cat. 11442074001 |
| M-MLV Reverse Transcriptase | Promega | Cat. M1701 |
| Pierce protein G agarose | Thermo Fisher | Cat. 20399 |
| T3 RNA polymerase | Promega | Cat. P2083 |
| DIG RNA Labeling Mix | Roche | Cat. 11277073910 |
| Anti-Digoxigenin-AP, Fab fragments | Roche | Cat. 11093274910 |
| Cytochalasin B | Sigma-Aldrich | Cat. C6762 |
| ChromaTide Alexa fluor 546-14-UTP | Life Technologies | Cat. C11404 |
| Hoechst 33342 | Invitrogen | Cat. H1399 |
| Critical commercial assays | ||
| ECL™ Prime Western Blotting Detection Reagent | Amersham | Cat. RPN2236 |
| PureLink™ RNA Mini Kit | Ambion | Cat. 12183025 |
| mMESSAGE mMACHINE Kit for SP6 | Ambion | Cat. AM1340 |
| Deposited data | ||
| RNA-sequencing raw and analyzed data | (Hwang et al., 2022) 16 | GEO: GSE199254 |
| RNA-seq raw and analyzed data for A-V axis | (Sindelka et al., 2018) 26 | GEO: GSE104848 |
| RNA-seq analyzed data for developmental gene expression of Xenopus laevis | (Session et al., 2016) 31 | GEO: GSE73430 |
| Mass spectrometry analyzed data | (Peuchen et al., 2017) 54 | MassIVE: MSV000081416 |
| Experimental models: Organisms/strains | ||
| Xenopus laevis: Wild type | Nasco | LM00715 |
| Xenopus laevis: Dria transgenic line | (Taguchi et al., 2012) 64 | NXR |
| Danio rerio: TÜ (Tübingen) lines | ZIRC (Zebrafish International Resource Center) | ZL57 |
| Danio rerio: bucky ball (buc) -/- mutant | (Dosch et al., 2004) 65 | N/A |
| Oligonucleotides | ||
| See S-table 4 for a list of oligonucleotides | ||
| Recombinant DNA | ||
| pCS2-Xvelo1 | This paper | N/A |
| pCS2-dnd1 | (Aguero et al., 2017) 66 | N/A |
| pCS2-nanos1 | (Aguero et al., 2017) 66 | N/A |
| pCS2-pgat | (Aguero et al., 2017) 66 | N/A |
| pCS2-mCherry-Myc-Xvelo1 | This paper | N/A |
| Ddx25 | Francis Crick Institute | TEgg065i20 |
| Software and algorithms | ||
| GraphPad Prism | GraphPad | https://www.graphpad.com/ |
| Metascape | (Zhou et al., 2019) 67 | https://metascape.org |
| ImageJ | (Schneider et al., 2012) 63 | https://imagej.net/ij/index.html |
| STRING | (Von Mering et al., 2005) 68 | https://string-db.org |
| BioRender | BioRender | https://www.biorender.com |
The Soluble-to-Insoluble Phase Transition (S-I): Transcripts that were increased more than two times and were enriched more than 15% in the pellet fraction of mature eggs after oocyte maturation (863).
The Insoluble-to-Soluble Phase Transition (I-S): Transcripts that were enriched more than 40% in the pellet fraction of oocytes and those pellet fraction is reduced by more than 25% after oocyte maturation (165).
The Insoluble-Insoluble State (I-I): Among the transcripts enriched more than 40% in the oocyte pellet fraction, the rest of the transcripts excluding those belonging to the I-S (68).
All GO and Protein-Protein interaction (PPI) results were generated by the criteria above.
Comparison to RNA localization data along the A-V axis in Xenopus eggs
The distribution of 15,005 transcripts along the animal-vegetal axis in the Xenopus laevis eggs has been reported previously 26. Among them, 10,798 transcripts were detected in our fractionation RNA-seq. Among these 10,798 transcripts, 573, 88, and 4 transcripts were detected in S-I, I-S, and I-I groups, respectively. The heat maps were generated by the expression level of each section along the animal-vegetal axis.
Comparison to RNA expression level during the early Xenopus embryonic development
RNA-seq dataset analyzing the expression level of Xenopus early embryos was reported previously 31. 11,169 transcripts were detected in both our fractionation RNA-seq and the Session study. Among these, 105 transcripts that are not expressed in the egg were filtered out. We analyzed the expression of RNAs in mature eggs, stages 8, 9, 10, 12, and 15 embryos. The results were normalized by the expression level in the mature egg. Since this analysis aimed to show the degradation of maternal RNAs in early embryonic development, early zygotic genes were not included in the expression analysis. We defined early zygotic genes as those showing a normalized value higher than 1. In our analysis, when the expression level of a specific gene was decreased by more than 25 % in the following stage, that gene was deemed to occurring degradation. Based on those criteria, all transcripts were classified into three groups below.
Class A (Rapid degraded genes, 2288 transcripts): Transcripts that started to degrade between the mature egg to stage 8 embryos and between stages 8 to stage 9 embryos.
Class B (Slowly degraded genes, 6101 transcripts): Transcripts that started to degrade between stages 9 to 10 embryos and between stages 10 to 11 embryos.
Class C (Relatively stable genes, 2675 transcripts): Transcripts that excluded those belonging to classes A and B.
All heat map results and diagrams were generated by the criteria above.
Comparison to proteomic data in Xenopus oocytes and embryos
The expression levels of proteins from stage VI oocyte to stage 2 embryo have been reported previously 54. Among all proteins detected by mass spec, 166, 21, and 2 were encoded by S-I, I-S, and I-I transcripts, respectively. The expression of these proteins was converted to the log scale and normalized by the expression levels in mature eggs. The heatmap was generated based on the protein expression levels in each stage to visualize the changes in protein levels.
Quantification and Statistical Analysis
All information about the statistical details is provided in the figure legends. Visualization for RNA-seq and proteomic profile-related results was performed by R studio. All image analysis and statistical tests were performed by ImageJ 63 and GraphPad Prism 10, respectively.
Supplementary Material
Table S1. List of I-S, S-I, and I-I transcripts, related to Figure 1.
Table S2. Distribution of I-S, S-I, and I-I transcripts along the animal-vegetal axis, related to Figure 1.
Table S3. Degradation of maternal transcripts during early embryonic development, related to Figure 2.
Table S4. Sequences of primers used for RT-qPCR analysis, related to STAR Methods section.
Highlights:
Many maternal RNAs transition between soluble and insoluble states during the OET
Germline RNAs undergo soluble-to-insoluble phase transition during the OET
Insoluble germline RNAs are resistant to RNA degradation machinery
Degradation of Velo/Buc during the OET allows solubilization of germline RNAs
Acknowledgment
We thank Dr. Mary Mullins for providing the bucky ball mutant zebrafish, Jia Fu for technical support. JY is supported by a grant from NIH (R35 GM131810). WM is supported by NIH grants (R03AI146900 and R01GM140306).
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
Declaration of interests
The authors declare no competing interests.
References
- 1.Schultz RM, Stein P, and Svoboda P (2018). The oocyte-to-embryo transition in mouse: past, present, and future. Biol Reprod 99, 160–174. 10.1093/biolre/ioy013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Jaffe LA, and Terasaki M (1994). Structural changes in the endoplasmic reticulum of starfish oocytes during meiotic maturation and fertilization. Dev Biol 164, 579–587. 10.1006/dbio.1994.1225. [DOI] [PubMed] [Google Scholar]
- 3.Terasaki M, and Jaffe LA (1991). Organization of the sea urchin egg endoplasmic reticulum and its reorganization at fertilization. J Cell Biol 114, 929–940. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Mehlmann LM, Terasaki M, Jaffe LA, and Kline D (1995). Reorganization of the endoplasmic reticulum during meiotic maturation of the mouse oocyte. Dev Biol 170, 607–615. 10.1006/dbio.1995.1240. [DOI] [PubMed] [Google Scholar]
- 5.Shiraishi K, Okada A, Shirakawa H, Nakanishi S, Mikoshiba K, and Miyazaki S (1995). Developmental changes in the distribution of the endoplasmic reticulum and inositol 1,4,5-trisphosphate receptors and the spatial pattern of Ca2+ release during maturation of hamster oocytes. Dev Biol 170, 594–606. 10.1006/dbio.1995.1239. [DOI] [PubMed] [Google Scholar]
- 6.Kume S, Yamamoto A, Inoue T, Muto A, Okano H, and Mikoshiba K (1997). Developmental expression of the inositol 1,4,5-trisphosphate receptor and structural changes in the endoplasmic reticulum during oogenesis and meiotic maturation of Xenopus laevis. Dev Biol 182, 228–239. 10.1006/dbio.1996.8479. [DOI] [PubMed] [Google Scholar]
- 7.Kline D. (2000). Attributes and dynamics of the endoplasmic reticulum in mammalian eggs. Current topics in developmental biology 50, 125–154. [DOI] [PubMed] [Google Scholar]
- 8.Terasaki M, Runft LL, and Hand AR (2001). Changes in organization of the endoplasmic reticulum during Xenopus oocyte maturation and activation. Mol Biol Cell 12, 1103–1116. 10.1091/mbc.12.4.1103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.FitzHarris G, Marangos P, and Carroll J (2007). Changes in endoplasmic reticulum structure during mouse oocyte maturation are controlled by the cytoskeleton and cytoplasmic dynein. Dev Biol 305, 133–144. 10.1016/j.ydbio.2007.02.006. [DOI] [PubMed] [Google Scholar]
- 10.Stitzel ML, and Seydoux G (2007). Regulation of the oocyte-to-zygote transition. Science 316, 407–408. 10.1126/science.1138236. [DOI] [PubMed] [Google Scholar]
- 11.Deshler JO, Highett MI, and Schnapp BJ (1997). Localization of Xenopus Vg1 mRNA by Vera protein and the endoplasmic reticulum. Science 276, 1128–1131. 10.1126/science.276.5315.1128. [DOI] [PubMed] [Google Scholar]
- 12.Alarcon VB, and Elinson RP (2001). RNA anchoring in the vegetal cortex of the Xenopus oocyte. J Cell Sci 114, 1731–1741. [DOI] [PubMed] [Google Scholar]
- 13.Chang P, Torres J, Lewis RA, Mowry KL, Houliston E, and King ML (2004). Localization of RNAs to the mitochondrial cloud in Xenopus oocytes through entrapment and association with endoplasmic reticulum. Mol Biol Cell 15, 4669–4681. 10.1091/mbc.e04-03-0265. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Prodon F, Dru P, Roegiers F, and Sardet C (2005). Polarity of the ascidian egg cortex and relocalization of cER and mRNAs in the early embryo. J Cell Sci 118, 2393–2404. 10.1242/jcs.02366. [DOI] [PubMed] [Google Scholar]
- 15.Sardet C, Nishida H, Prodon F, and Sawada K (2003). Maternal mRNAs of PEM and macho 1, the ascidian muscle determinant, associate and move with a rough endoplasmic reticulum network in the egg cortex. Development 130, 5839–5849. 10.1242/dev.00805. [DOI] [PubMed] [Google Scholar]
- 16.Hwang H, Yun S, Arcanjo RB, Divyanshi, Chen S, Mei W, Nowak RA, Kwon T, and Yang J (2022). Regulation of RNA localization during oocyte maturation by dynamic RNA-ER association and remodeling of the ER. Cell Rep 41, 111802. 10.1016/j.celrep.2022.111802. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Solter D, Hiiragi T, Evsikov AV, Moyer J, De Vries WN, Peaston AE, and Knowles BB (2004). Epigenetic mechanisms in early mammalian development. Cold Spring Harb Symp Quant Biol 69, 11–17. 10.1101/sqb.2004.69.11. [DOI] [PubMed] [Google Scholar]
- 18.Hwang H, Jin Z, Krishnamurthy VV, Saha A, Klein PS, Garcia B, Mei W, King ML, Zhang K, and Yang J (2019). Novel functions of the ubiquitin-independent proteasome system in regulating Xenopus germline development. Development 146. 10.1242/dev.172700. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Liu Y, Zhao H, Shao F, Zhang Y, Nie H, Zhang J, Li C, Hou Z, Chen ZJ, Wang J, et al. (2023). Remodeling of maternal mRNA through poly(A) tail orchestrates human oocyte-to-embryo transition. Nat Struct Mol Biol. 10.1038/s41594-022-00908-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Shi B, Zhang J, Heng J, Gong J, Zhang T, Li P, Sun BF, Yang Y, Zhang N, Zhao YL, et al. (2020). RNA structural dynamics regulate early embryogenesis through controlling transcriptome fate and function. Genome Biol 21, 120. 10.1186/s13059-020-02022-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Marlow FL, and Mullins MC (2008). Bucky ball functions in Balbiani body assembly and animal-vegetal polarity in the oocyte and follicle cell layer in zebrafish. Dev Biol 321, 40–50. 10.1016/j.ydbio.2008.05.557. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Bontems F, Stein A, Marlow F, Lyautey J, Gupta T, Mullins MC, and Dosch R (2009). Bucky ball organizes germ plasm assembly in zebrafish. Curr Biol 19, 414–422. 10.1016/j.cub.2009.01.038. [DOI] [PubMed] [Google Scholar]
- 23.Nijjar S, and Woodland HR (2013). Protein interactions in Xenopus germ plasm RNP particles. PLoS ONE 8, e80077. 10.1371/journal.pone.0080077. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Boke E, Ruer M, Wuhr M, Coughlin M, Lemaitre R, Gygi SP, Alberti S, Drechsel D, Hyman AA, and Mitchison TJ (2016). Amyloid-like Self-Assembly of a Cellular Compartment. Cell 166, 637–650. 10.1016/j.cell.2016.06.051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Houston DW (2013). Regulation of cell polarity and RNA localization in vertebrate oocytes. Int Rev Cell Mol Biol 306, 127–185. 10.1016/B978-0-12-407694-5.00004-3. [DOI] [PubMed] [Google Scholar]
- 26.Sindelka R, Abaffy P, Qu Y, Tomankova S, Sidova M, Naraine R, Kolar M, Peuchen E, Sun L, Dovichi N, and Kubista M (2018). Asymmetric distribution of biomolecules of maternal origin in the Xenopus laevis egg and their impact on the developmental plan. Sci Rep 8, 8315. 10.1038/s41598-018-26592-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Newport J, and Kirschner M (1982). A major developmental transition in early Xenopus embryos: II. Control of the onset of transcription. Cell 30, 687–696. 0092-8674(82)90273-2 [pii]. [DOI] [PubMed] [Google Scholar]
- 28.Newport J, and Kirschner M (1982). A major developmental transition in early Xenopus embryos: I. characterization and timing of cellular changes at the midblastula stage. Cell 30, 675–686. 0092-8674(82)90272-0 [pii]. [DOI] [PubMed] [Google Scholar]
- 29.Howe JA, Howell M, Hunt T, and Newport JW (1995). Identification of a developmental timer regulating the stability of embryonic cyclin A and a new somatic A-type cyclin at gastrulation. Genes Dev 9, 1164–1176. [DOI] [PubMed] [Google Scholar]
- 30.Howe JA, and Newport JW (1996). A developmental timer regulates degradation of cyclin E1 at the midblastula transition during Xenopus embryogenesis. Proc Natl Acad Sci U S A 93, 2060–2064. 10.1073/pnas.93.5.2060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Session AM, Uno Y, Kwon T, Chapman JA, Toyoda A, Takahashi S, Fukui A, Hikosaka A, Suzuki A, Kondo M, et al. (2016). Genome evolution in the allotetraploid frog Xenopus laevis. Nature 538, 336–343. 10.1038/nature19840. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.MacArthur H, Houston DW, Bubunenko M, Mosquera L, and King ML (2000). DEADSouth is a germ plasm specific DEAD-box RNA helicase in Xenopus related to eIF4A. Mechanisms of development 95, 291–295. [DOI] [PubMed] [Google Scholar]
- 33.Zhou Y, and King ML (1996). Localization of Xcat-2 RNA, a putative germ plasm component, to the mitochondrial cloud in Xenopus stage I oocytes. Development 122, 2947–2953. [DOI] [PubMed] [Google Scholar]
- 34.Horvay K, Claussen M, Katzer M, Landgrebe J, and Pieler T (2006). Xenopus Dead end mRNA is a localized maternal determinant that serves a conserved function in germ cell development. Dev Biol 291, 1–11. 10.1016/j.ydbio.2005.06.013. [DOI] [PubMed] [Google Scholar]
- 35.Owens DA, Butler AM, Aguero TH, Newman KM, Van Booven D, and King ML (2017). High-throughput analysis reveals novel maternal germline RNAs crucial for primordial germ cell preservation and proper migration. Development 144, 292–304. 10.1242/dev.139220. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Houston DW, Zhang J, Maines JZ, Wasserman SA, and King ML (1998). A Xenopus DAZ-like gene encodes an RNA component of germ plasm and is a functional homologue of Drosophila boule. Development 125, 171–180. [DOI] [PubMed] [Google Scholar]
- 37.Hudson C, and Woodland HR (1998). Xpat, a gene expressed specifically in germ plasm and primordial germ cells of Xenopus laevis. Mechanisms of development 73, 159–168. [DOI] [PubMed] [Google Scholar]
- 38.Tarbashevich K, Koebernick K, and Pieler T (2007). XGRIP2.1 is encoded by a vegetally localizing, maternal mRNA and functions in germ cell development and anteroposterior PGC positioning in Xenopus laevis. Dev Biol 311, 554–565. 10.1016/j.ydbio.2007.09.012. [DOI] [PubMed] [Google Scholar]
- 39.Oh D, and Houston DW (2017). Role of maternal Xenopus syntabulin in germ plasm aggregation and primordial germ cell specification. Dev Biol 432, 237–247. 10.1016/j.ydbio.2017.10.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Claussen M, and Pieler T (2004). Xvelo1 uses a novel 75-nucleotide signal sequence that drives vegetal localization along the late pathway in Xenopus oocytes. Dev Biol 266, 270–284. 10.1016/j.ydbio.2003.09.043. [DOI] [PubMed] [Google Scholar]
- 41.Heasman J, Holwill S, and Wylie CC (1991). Fertilization of cultured Xenopus oocytes and use in studies of maternally inherited molecules. Methods in cell biology 36, 213–230. [DOI] [PubMed] [Google Scholar]
- 42.Ye D, Zhu L, Zhang Q, Xiong F, Wang H, Wang X, He M, Zhu Z, and Sun Y (2019). Abundance of Early Embryonic Primordial Germ Cells Promotes Zebrafish Female Differentiation as Revealed by Lifetime Labeling of Germline. Mar Biotechnol (NY) 21, 217–228. 10.1007/s10126-019-09874-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Elkouby YM, Jamieson-Lucy A, and Mullins MC (2016). Oocyte Polarization Is Coupled to the Chromosomal Bouquet, a Conserved Polarized Nuclear Configuration in Meiosis. PLoS Biol 14, e1002335. 10.1371/journal.pbio.1002335. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Beer RL, and Draper BW (2013). nanos3 maintains germline stem cells and expression of the conserved germline stem cell gene nanos2 in the zebrafish ovary. Dev Biol 374, 308–318. 10.1016/j.ydbio.2012.12.003. [DOI] [PubMed] [Google Scholar]
- 45.Weidinger G, Stebler J, Slanchev K, Dumstrei K, Wise C, Lovell-Badge R, Thisse C, Thisse B, and Raz E (2003). dead end, a novel vertebrate germ plasm component, is required for zebrafish primordial germ cell migration and survival. Curr Biol 13, 1429–1434. [DOI] [PubMed] [Google Scholar]
- 46.Howley C, and Ho RK (2000). mRNA localization patterns in zebrafish oocytes. Mechanisms of development 92, 305–309. 10.1016/s0925-4773(00)00247-1. [DOI] [PubMed] [Google Scholar]
- 47.Wang H, Teng Y, Xie Y, Wang B, Leng Y, Shu H, and Deng F (2013). Characterization of the carbonic anhydrases 15b expressed in PGCs during early zebrafish development. Theriogenology 79, 443–452. 10.1016/j.theriogenology.2012.10.016. [DOI] [PubMed] [Google Scholar]
- 48.Strasser MJ, Mackenzie NC, Dumstrei K, Nakkrasae LI, Stebler J, and Raz E (2008). Control over the morphology and segregation of Zebrafish germ cell granules during embryonic development. BMC Dev Biol 8, 58. 10.1186/1471-213X-8-58. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Yoon C, Kawakami K, and Hopkins N (1997). Zebrafish vasa homologue RNA is localized to the cleavage planes of 2- and 4-cell-stage embryos and is expressed in the primordial germ cells. Development 124, 3157–3165. 10.1242/dev.124.16.3157. [DOI] [PubMed] [Google Scholar]
- 50.Riemer S, Bontems F, Krishnakumar P, Gomann J, and Dosch R (2015). A functional Bucky ball-GFP transgene visualizes germ plasm in living zebrafish. Gene Expr Patterns 18, 44–52. 10.1016/j.gep.2015.05.003. [DOI] [PubMed] [Google Scholar]
- 51.Berchowitz LE, Kabachinski G, Walker MR, Carlile TM, Gilbert WV, Schwartz TU, and Amon A (2015). Regulated Formation of an Amyloid-like Translational Repressor Governs Gametogenesis. Cell 163, 406–418. 10.1016/j.cell.2015.08.060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Sato K, Sakai M, Ishii A, Maehata K, Takada Y, Yasuda K, and Kotani T (2022). Identification of embryonic RNA granules that act as sites of mRNA translation after changing their physical properties. iScience 25, 104344. 10.1016/j.isci.2022.104344. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Thomas MG, Pascual ML, Maschi D, Luchelli L, and Boccaccio GL (2014). Synaptic control of local translation: the plot thickens with new characters. Cell Mol Life Sci 71, 2219–2239. 10.1007/s00018-013-1506-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Peuchen EH, Cox OF, Sun L, Hebert AS, Coon JJ, Champion MM, Dovichi NJ, and Huber PW (2017). Phosphorylation Dynamics Dominate the Regulated Proteome during Early Xenopus Development. Sci Rep 7, 15647. 10.1038/s41598-017-15936-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Kloc M, Bilinski S, Chan AP, Allen LH, Zearfoss NR, and Etkin LD (2001). RNA localization and germ cell determination in Xenopus. Int Rev Cytol 203, 63–91. [DOI] [PubMed] [Google Scholar]
- 56.King ML, Messitt TJ, and Mowry KL (2005). Putting RNAs in the right place at the right time: RNA localization in the frog oocyte. Biol Cell 97, 19–33. [DOI] [PubMed] [Google Scholar]
- 57.Oh D, and Houston DW (2017. RNA Localization in the Vertebrate Oocyte: Establishment of Oocyte Polarity and Localized mRNA Assemblages. Results Probl Cell Differ 63, 189–208. 10.1007/978-3-319-60855-6_9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Jamieson-Lucy A, and Mullins MC (2019). The vertebrate Balbiani body, germ plasm, and oocyte polarity. Current topics in developmental biology 135, 1–34. 10.1016/bs.ctdb.2019.04.003. [DOI] [PubMed] [Google Scholar]
- 59.O’Connell LC, and Mowry KL (2021). Regulation of spatially restricted gene expression: linking RNA localization and phase separation. Biochem Soc Trans 49, 2591–2600. 10.1042/BST20210320. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Houston DW (2018). Oocyte Host-Transfer and Maternal mRNA Depletion Experiments in Xenopus. Cold Spring Harb Protoc 2018. 10.1101/pdb.prot096982. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Lee KL, and Marlow FL (2019). Visualizing the Balbiani Body in Zebrafish Oocytes. Methods Mol Biol 1920, 277–293. 10.1007/978-1-4939-9009-2_16. [DOI] [PubMed] [Google Scholar]
- 62.Rorick AM, Mei W, Liette NL, Phiel C, El-Hodiri HM, and Yang J (2007). PP2A:B56epsilon is required for eye induction and eye field separation. Dev Biol 302, 477–493. [DOI] [PubMed] [Google Scholar]
- 63.Schneider CA, Rasband WS, and Eliceiri KW (2012). NIH Image to ImageJ: 25 years of image analysis. Nat Methods 9, 671–675. 10.1038/nmeth.2089. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Taguchi A, Takii M, Motoishi M, Orii H, Mochii M, and Watanabe K (2012). Analysis of localization and reorganization of germ plasm in Xenopus transgenic line with fluorescence-labeled mitochondria. Dev Growth Differ 54, 767–776. 10.1111/dgd.12005. [DOI] [PubMed] [Google Scholar]
- 65.Dosch R, Wagner DS, Mintzer KA, Runke G, Wiemelt AP, and Mullins MC (2004). Maternal control of vertebrate development before the midblastula transition: mutants from the zebrafish I. Dev Cell 6, 771–780. 10.1016/j.devcel.2004.05.002. [DOI] [PubMed] [Google Scholar]
- 66.Aguero T, Jin Z, Chorghade S, Kalsotra A, King ML, and Yang J (2017). Maternal Dead-end 1 promotes translation of nanos1 by binding the eIF3 complex. Development 144, 3755–3765. 10.1242/dev.152611. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Zhou Y, Zhou B, Pache L, Chang M, Khodabakhshi AH, Tanaseichuk O, Benner C, and Chanda SK (2019). Metascape provides a biologist-oriented resource for the analysis of systems-level datasets. Nature communications 10, 1523. 10.1038/s41467-019-09234-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.von Mering C, Jensen LJ, Snel B, Hooper SD, Krupp M, Foglierini M, Jouffre N, Huynen MA, and Bork P (2005). STRING: known and predicted protein-protein associations, integrated and transferred across organisms. Nucleic acids research 33, D433–437. 10.1093/nar/gki005. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Table S1. List of I-S, S-I, and I-I transcripts, related to Figure 1.
Table S2. Distribution of I-S, S-I, and I-I transcripts along the animal-vegetal axis, related to Figure 1.
Table S3. Degradation of maternal transcripts during early embryonic development, related to Figure 2.
Table S4. Sequences of primers used for RT-qPCR analysis, related to STAR Methods section.
Data Availability Statement
RNA sequencing data analyzed in this study have been deposited to the Gene Expression Omnibus (GEO) with the dataset identifier GSE199254.
This paper does not report the original code.
Any additional information required to reanalyze the data reported in this paper is available from the lead contact upon request.