

Abstract
The subcellular locations, ultrastructure, and cytotoxic activity of the GroEL-like protein from Actinobacillus actinomycetemcomitans were investigated. Two-dimensional sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) clearly indicated that synthesis of the GroEL-like protein is substantially increased after a thermal shock. Analysis of the purified native GroEL-like protein by transmission electron microscopy revealed the typical 14-mer cylindrical molecule, which had a diameter of about 12 nm. A. actinomycetemcomitans cells grown at 35°C and heat shocked at 43°C were fractionated, and fractions were separated by SDS-PAGE and analyzed by Western immunoblotting using antibodies to GroEL- and DnaK-like proteins. The GroEL-like protein was found in both the soluble and membrane fractions, whereas the DnaK-like protein was mostly found in the cytoplasm. An increase in specific proteins, including the GroEL- and DnaK-like proteins, was found in heat-shocked cells. The subcellular localization of the GroEL-like protein was examined by immunoelectron microscopy of whole cells. More GroEL-like protein was detected in stressed cells than in unstressed cells, and most of it was found not directly associated with outer membranes but rather in extracellular material. The native GroEL-like protein was assessed for cytotoxic activities. The GroEL-like protein increased the proliferation of periodontal ligament epithelial cells at concentrations between 0.4 and 1.0 μg/ml. The number of cells in the culture decreased significantly at higher concentrations. A cell viability assay using HaCaT epithelial cells indicated that the GroEL-like protein was strongly toxic for the cells. These studies suggest the extracellular nature of the GroEL-like protein and its putative role in disease initiation.
Periodontal disease is a family of soft-hard tissue diseases that involves complex bacterium-host interactions. Some forms of periodontal diseases are chronic, slowly progressive infectious processes (such as adult periodontitis), while others progress more rapidly (such as localized juvenile periodontitis [LJP]). Despite the fact that periodontal diseases are mixed infections, considerable evidence suggests that Actinobacillus actinomycetemcomitans is the major causative agent of LJP, a disease involving inflammation of the gingiva and destruction of the periodontal ligament and alveolar bone, resulting in tooth loss (42). Patients with LJP exhibit elevated levels of antibody to the bacterium in serum, saliva, and gingival crevicular fluid (5, 29). Although this bacterium exhibits many putative virulence factors (reviewed in references 26 and 42), the actual in vivo mechanisms by which A. actinomycetemcomitans maintains itself in the oral cavity, evades or interferes with host defenses, destroys host tissues, and inhibits tissue repair remain poorly understood.
In the course of an infection, A. actinomycetemcomitans is likely exposed to a number of environmental changes which may induce the bacterium to synthesize heat shock proteins (Hsp’s), or stress proteins. Hsp’s are highly conserved proteins that play important roles in the physiology of any cell (40). Hsp’s have been grouped into families according to molecular mass, and members of a family have common features (6). Some Hsp’s function as chaperones and may help in transporting proteins across cell membranes or assist in protein folding (6, 10, 37), while others may play a key role in the assembly of cell surface components such as fimbriae (39). Hsp’s may also play a role in microbial pathogenicity. Several important antigenic components observed in a variety of bacterial infections and involved in pathogenesis of the disease have been identified as members of stress protein families (13, 16). They are often identified as dominant antigens in microorganisms and are therefore capable of inducing strong humoral and cellular responses (22). Recent data on the cell surface expression of Hsp’s in eukaryotic cells and the immunological consequences of these proteins have been reviewed by Multhoff and Hightower (28), but very little is known about cell surface expression of Hsp’s in prokaryotes.
The heat shock response in A. actinomycetemcomitans was originally studied by Koga et al. (20) and by Løkensgard et al. (23). Several proteins were identified as Hsp’s, and at least one of them (a 60-kDa protein) reacted strongly with antibodies raised against prokaryotic GroEL or eukaryotic GroEL-like protein. In bacteria, GroEL-like proteins have been shown to bind to nascent proteins and help maintain their secondary structure during stressful conditions (10, 34). Cloning and molecular characterization of the gene for the GroEL-like protein of A. actinomycetemcomitans were reported by Koga et al. (20) and Nakano et al. (30).
Several studies have shown that the GroEL-like protein, a molecular chaperone, can be an immunodominant antigen (1, 15, 20), but its major localization seems to vary with the organism. Recent studies using immunocytochemical procedures indicated that GroEL-like proteins of Mycobacterium species, Borrelia burgdorferi, and Haemophilus ducreyi are located in compartments other than the cytoplasm (8, 11, 34). Surface-associated Hsp60 has been reported in Helicobacter pylori (33), Mycobacterium leprae (14), and Salmonella typhimurium (7). We describe the localization and ultrastructure of the GroEL-like protein of A. actinomycetemcomitans, determined with specific antibodies directed against the purified GroEL-like protein and by immunoelectron microscopy. The cytotoxic effect of the purified native GroEL-like protein on epithelial cells is also reported.
MATERIALS AND METHODS
Bacteria and growth conditions.
A. actinomycetemcomitans ATCC 29522 serotype b and ATCC 33384 serotype c were grown in Todd-Hewitt broth (BBL Microbiology Systems, Cockeysville, Md.) supplemented with 1% yeast extract and incubated at 35°C in an anaerobic chamber (N2/H2/CO2 ratio, 80:10:10).
GroEL-like protein isolation and antibody production.
Purification of the native GroEL-like protein (Hsp64) from A. actinomycetemcomitans ATCC 29522 following a heat shock treatment at 43°C for 1 h was carried out as follows. Harvested cells were washed twice in ice-cold 50 mM phosphate-buffered saline (PBS; pH 7.2), suspended in 75 ml of cell lysis buffer (50 mM Tris-HCl, 1 mM EDTA, 0.2 mM N-tosyl-l-lysine chloromethyl ketone [TLCK], and 0.3 mg of lysozyme/ml [pH 8.0]), and kept on ice for 30 min. Cells were broken by sonication, cell debris were removed, and the material was prepared for affinity chromatography on ATP-agarose according to the method of Hinode et al. (17). The fractions containing the GroEL-like and DnaK-like proteins were collected, pooled, dialyzed against PBS (pH 7.0), and freeze-dried. GroEL-like and DnaK-like proteins in this fraction were separated by size exclusion chromatography. The sample (1 ml) was applied onto a Sepharose CL-4B column (1.5 by 30 cm) that had been equilibrated with PBS, and gel filtration chromatography was performed with a flow rate of 0.1 ml/min. The eluted fractions (2 ml/tube) were analyzed for the GroEL-like protein by both dot immunoblotting using commercial antibodies (rabbit anti-Hsp70 raised against DnaK from Escherichia coli and used at a 1:1,000 dilution [Dako, Mississauga, Ontario, Canada] and rabbit anti-Hsp60 raised against Hsp60 from Synechococcus sp. and used at a 1:3,000 dilution [StressGen, Victoria, British Columbia, Canada]) and sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) followed by silver nitrate staining. The fractions containing either the GroEL- or DnaK-like proteins were pooled. The denatured (obtained after SDS-PAGE and electroelution) GroEL-like protein (10 μg) was injected intracutaneously into a New Zealand White rabbit in the presence of complete Freund adjuvant. Subsequent intramuscular injections in the presence of incomplete Freund adjuvant were carried out at days 8, 19, and 40. The rabbit was bled via the marginal ear vein at days 25 and 33 and via the heart directly at day 48. The antisera were pooled and stored at −20°C until used.
Estimation of the molecular mass of the GroEL-like protein.
Surface-associated material (SAM) and purified native and denatured GroEL-like proteins, in 3-[(3-cholamidopropyl)-dimethylammonio]-1-propanesulfonate (CHAPS; 0.1% [wt/vol]; ICN Biochemicals Inc., Cleveland, Ohio), were submitted to ultrafiltration through 300- and 100-kDa molecular size cutoff filters (Ultrafree-MC; Millipore, Bedford, Mass.) by using a centrifuge (at 5,000 × g for 10 min at 4°C). Resulting filtrates and supernatants for the SAM and native and denatured GroEL-like proteins were then analyzed by Western immunoblotting as described below.
Heat shock and cell fractionation.
Two cultures (400 ml; optical density at 660 nm [OD660] = 0.6) of A. actinomycetemcomitans ATCC 29522 were either maintained at 35°C (control) or elevated to 43°C (heat shocked) for 1 h. Cells were centrifuged and washed twice in 50 mM Tris buffer (pH 7.8) containing 0.03 M NaCl. The culture supernatants were kept for vesicle preparation (see below). The cells were then resuspended in 12 ml of 50 mM Tris (pH 7.8) containing 30% sucrose and 1 mM EDTA. The suspension was left for 15 min at room temperature and then centrifuged at 8,000 × g for 30 min. The supernatant was discarded, and the cell pellet was suspended in 40 ml of ice-cold distilled water (osmotic shock). After 5 min, the suspension was centrifuged at 8,000 × g for 30 min. The supernatant containing the periplasmic material was lyophilized. The pellet was suspended in 4 ml of 50 mM Tris (pH 7.8) containing 10% glycerol, 2 mM MgCl2, 0.2 mg of DNase/ml, and 0.2 mg of RNase/ml, followed by a mild ultrasonic treatment to break cells and disperse the material. The material was centrifuged at 6,000 × g for 15 min, and the supernatant was centrifuged at 200,000 × g for 2 h. The resulting supernatant (∼4 ml) was dialyzed extensively against distilled water and contained the cytoplasmic material. The pellet was resuspended in 2 ml of 2% Triton X-100 containing 10 mM MgCl2. The latter suspension was centrifuged at 200,000 × g for 1 h. The supernatant (∼2 ml) contained the cytoplasmic-membrane-rich cell envelope, and the pellet resuspended in 2 ml of distilled water contained the outer-membrane-rich cell envelope. The vesicles were collected by addition of ammonium sulfate to the culture supernatants to a final saturation of 40% at 4°C. The treated medium was then centrifuged at 20,000 × g for 20 min, and the pellet was suspended in PBS. Vesicles were washed twice by centrifugation (at 27,000 × g for 40 min) and resuspended in 400 μl of PBS. Alternatively, the vesicles were collected by ultracentrifugation (at 100,000 × g for 2 h) of the culture supernatants and were resuspended in 20 ml of 50 mM Tris buffer (pH 7.2) containing 0.5 mM dithiothreitol. A final centrifugation was carried out (at 100,000 × g for 2 h), and vesicles were resuspended in 200 μl of Tris buffer (pH 7.2) and stored at −20°C. All the fractions were kept at −20°C prior to analysis for the presence of GroEL- and DnaK-like proteins.
The SAM from cells of A. actinomycetemcomitans was obtained by the method described by Kirby et al. (19), modified as follows. A. actinomycetemcomitans was grown to mid-log phase. Aliquots of 500 ml were heat shocked at 43°C for 30 min, while a control sample remained at 35°C. Unstressed and heat-stressed cells were harvested by centrifugation at 20,000 × g for 30 min at 4°C and were washed once with 50 mM phosphate buffer (pH 7.2). The SAM was extracted by gentle stirring in saline (PBS) for 1 h at 4°C. Bacterial cells were removed by centrifugation at 30,000 × g for 45 min at 4°C. Supernatants were dialyzed overnight against fresh deionized water, lyophilized, and stored in CHAPS (0.1%, wt/vol) at −20°C.
Determination of cell lysis.
Cell lysis before or during a heat stress was evaluated by spotting washed mid-log cells (OD660 = 0.12 to 1.0) on an agarose gel (1%, wt/vol) and checking for the presence of DNA after the addition of ethidium bromide (0.05 μg/ml). The sensitivity of the procedure was evaluated by spotting native commercial DNA (Pharmacia Biotech, Uppsala, Sweden), and the lower limit of sensitivity was found to be 2.5 ng.
TEM.
A. actinomycetemcomitans ATCC 33384 and ATCC 29522 were grown to an OD660 of 0.25 (or as otherwise indicated) in supplemented Todd-Hewitt broth at 35°C. A tube was kept at 35°C, while another was placed at 43°C for 30 min. Aliquots of heat-shocked and unstressed cells were centrifuged (at 6,000 × g for 2 min) and washed twice in cold 50 mM phosphate buffer (pH 7.2). Stressed and unstressed cells were fixed overnight in PBS–0.1% glutaraldehyde (vol/vol)–3% paraformaldehyde (vol/vol). Pellets were dehydrated and embedded in LR-White. Ultrathin sections were prepared (Reichert ultracut E) and deposited on Formvar-coated nickel grids (JBEM, Dorval, Quebec, Canada). After an incubation of 1 h in 0.5% (wt/vol) bovine serum albumin (BSA) in PBS (50 mM; pH 7.2), the grids were incubated for 2 h at 37°C with either a rabbit anti-A. actinomycetemcomitans GroEL-like protein diluted 1/100 in BSA-PBS, a rabbit anti-pig immunoglobulin G (IgG)-alkaline phosphatase conjugate (Sigma Chemical Co., St. Louis, Mo.) diluted 1/50, or a rabbit anti-E. coli DnaK (Dako Co., Carpinteria, Calif.) diluted 1/50. Then they were washed in PBS and subsequently in fresh deionized water and were reacted with a gold (10 nm)-IgG conjugate (Sigma Chemical Co.) diluted 1/5 in BSA-PBS (0.5%, wt/vol) for 1 h and then stained with phosphotungstic acid (1%, wt/vol; Electron Microscopy Sciences, Fort Washington, Pa.). Grids were examined in a JEOL (model 1200 EX) transmission electron microscope (TEM) operating at 80 kV.
Ultrastructure of the GroEL-like protein determined by TEM.
Ten microliters of purified native GroEL-like protein resuspended in CHAPS (0.1%, wt/vol) was fixed on a Formvar-coated nickel grid for 10 min at room temperature and stained with uranyl acetate (1%, wt/vol). The grids were examined as described above.
SDS-PAGE and Western immunoblot analysis.
The protein concentration in the samples was determined by the method of Lowry et al. (24) by using BSA as a standard. SDS-PAGE was performed according to the procedure of Laemmli (21) by using 10% slab gels (Mini Protean II; Bio-Rad Laboratories, Richmond, Calif.). Electrophoresis was carried out at 200 V for 45 min. Proteins that migrated were stained with Coomassie brilliant blue R-250. Two-dimensional electrophoresis was performed according to the method of O’Farrell (31) by using a Mini Protean II 2D cell (Bio-Rad). The isoelectric focusing of the first dimension was performed at 500 V for 10 min, then at 700 V for 3.5 h over a pI range of 3 to 10. In the second dimension, proteins were separated by molecular mass as described above. Western immunoblotting was performed as described previously (17) by using rabbit antisera against the purified A. actinomycetemcomitans GroEL-like protein (1:5,000 dilution). Following a 1-h incubation with this first antibody, the nitrocellulose membrane was reacted with goat anti-rabbit antibody coupled to alkaline phosphatase (1:3,000 dilution; Bio-Rad Laboratories) for 1 h and was then developed with the alkaline phosphatase color development reagent.
Epithelial-cell growth and viability assays.
Porcine periodontal ligament epithelial cells were isolated from Malassez’ epithelial rests as described previously (3). The cells were cultured in minimal Eagle medium containing 0.01% (wt/vol) penicillin G, 0.1% (vol/vol) gentamicin sulfate, 1.2% (wt/vol) amphotericin B (Fungizone; final concentration, 30 μg of amphotericin B and 24.6 μg of sodium deoxycholate per ml; Gibco BRL), and 15% fetal bovine serum (Gibco BRL) as described previously (32). After cells were cultured for 2 days, the native GroEL-like protein was added and the cultures were continued for 7 days. To measure cell growth, the cultures were fixed with 4% formaldehyde–5% sucrose in 50 mM PBS and were stained with 0.1% crystal violet in 200 mM boric acid (pH 6.0). After the wells were washed with excess distilled water, the stain was dissolved with 10% acetic acid and its intensity was measured with a spectrophotometer at 570 nm. The stain intensity is directly proportional to the number of cells in the culture.
HaCaT cells, a line of spontaneously transformed nontumorigenic skin keratinocytes with characteristics similar to normal keratinocytes (2), were cultured in Dulbecco’s minimal Eagle medium supplemented with antibiotics as described above and 10% fetal calf serum in a humidified atmosphere of 5% CO2 and 95% air at 37°C. Appropriate dilutions of the cell suspension were plated on 96-well culture plates to yield about 8,000 cells per well. After 24 h of culturing, the native GroEL-like protein was added and the cultures were continued for 4 days. To study the viability of the cells treated with the GroEL-like protein, the conversion of tetrazolium salt into blue formazan was measured by using a Cell titer 96 kit (Promega, Madison, Wis.). To test if the effects of the GroEL-like protein are dependent on serum, the cells were also grown in special serum-free keratinocyte medium (KGM; Clonetics, San Diego, Calif.).
RESULTS
Heat shock response of A. actinomycetemcomitans.
Cells of A. actinomycetemcomitans grown at 35°C and transferred to 43°C for 1 h overexpressed several stress proteins that were visualized by SDS-PAGE, Western immunoblot analysis, and two-dimensional electrophoresis (Fig. 1). Among the Hsp’s, three with molecular masses of 64, 74, and 89 kDa were prominent after the stress, while the synthesis of other proteins was diminished.
FIG. 1.

Two-dimensional gel electrophoresis of proteins from unstressed (A) and stressed (B) cells. The isoelectric focusing gel is oriented with the acidic side to the left and high-molecular-mass proteins at the top. The GroEL-like protein (64 kDa) and the DnaK-like protein (74 kDa) are indicated by arrows in panel B. Circled dots in panel A are examples of proteins whose synthesis was lowered by the heat shock.
Ultrastructure and estimation of the molecular mass of native GroEL-like protein.
By use of ultrafilters with different pore sizes, the purified native GroEL-like protein of A. actinomycetemcomitans showed a molecular mass estimated to be higher than 300 kDa, as expected. However, the fraction contained molecules reacting with the anti-GroEL-like protein and having a molecular mass between 100 and 300 kDa (possibly smaller polymers of 2 to 4 copies of the 64-kDa polypeptide). Denatured GroEL-like protein exhibited a molecular mass below 100 kDa. The ultrastructure of the purified native GroEL-like protein was examined by TEM, and the typical 14-mer cylindrical molecule was observed and showed a diameter of about 12 nm (Fig. 2).
FIG. 2.
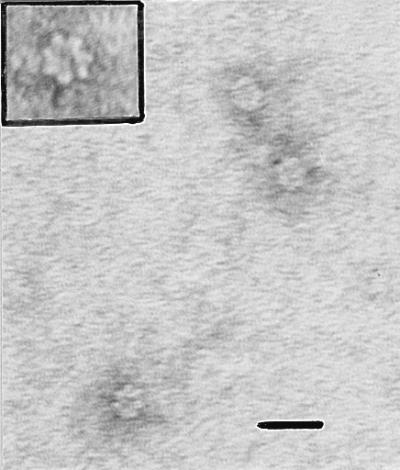
Ultrastructure of the purified native GroEL-like protein stained with uranyl acetate. The sevenfold symmetry can be seen and easily recognized in the enlargement (upper left corner). Bar = 20 nm.
Localization of the GroEL-like protein.
A. actinomycetemcomitans ATCC 29522 grown at 35°C and heat shocked at 43°C was fractionated. The proteins in each fraction were separated by SDS-PAGE and analyzed for GroEL- and DnaK-like proteins by Western immunoblotting (Fig. 3). GroEL-like protein was found in both the soluble (cytoplasmic and periplasmic) and membrane fractions (Fig. 3B). Of the GroEL-like protein found in the soluble fraction, most was detected in the cytoplasm, as expected. In the membrane fractions, most of the protein was found in the cytoplasmic membranes, although some stress proteins were also found in outer-membrane and vesicle fractions. Additional bands seen in Fig. 3B (lane 6) may be degradation products. When we probed with an anti-DnaK antibody, all of the reactivity was found in the cytoplasm and, to a much lower degree, in the periplasm (Fig. 3C). No reactivity was found in the other fractions.
FIG. 3.

Presence of the GroEL-like protein in various cell fractions. Cell fractionation was performed according to the method described in the text. Each lane contains 10 μg of proteins. Lanes 1, unstressed A. actinomycetemcomitans cells; lanes 2, stressed A. actinomycetemcomitans cells; lanes 3, cytoplasm; lanes 4, periplasm; lanes 5, outer membrane; lanes 6, cytoplasmic membrane; lanes 7, extracellular vesicles. The protein profile of each fraction was analyzed by SDS-PAGE and staining with Coomassie brilliant blue R-250 (A) and by Western immunoblot analysis with an anti-GroEL-like antibody (B) or with an anti-DnaK antibody (C).
The subcellular localization of GroEL-like protein in A. actinomycetemcomitans ATCC 33384 and ATCC 29522 was examined by immunoelectron microscopy using unstressed and heat-stressed cells. Results indicated that more reactivity (both inside and outside) was obtained for stressed cells than for unstressed cells (Fig. 4A and B). However, most of the reactivity that was detected outside the cells was not found on the external portion of the outer membranes, as was suggested by the cell fractionation results. The antibody also recognized diffused material in the surroundings of the bacteria. This was confirmed by the analysis of SAM from heat-stressed cells, which exhibited a higher level of GroEL-like protein than SAM from unstressed cells. Because of the well-known ability of A. actinomycetemcomitans to bind Fc components of IgG (38), the background level of immunogold labeling was evaluated with a rabbit anti-pig IgG or without any first antibody. These controls revealed very little labeling (Fig. 4C and D). Finally, we observed that A. actinomycetemcomitans cells stressed by heat also overexpressed DnaK-like proteins both inside and outside the cell (Fig. 4E and F). However, this protein was present in a much lower proportion outside the cells than GroEL-like protein, as previously suggested by the cell fractionation results; this supports the relevance of the localization study of the GroEL-like protein of A. actinomycetemcomitans. Finally, evaluation of cell lysis by DNA contamination outside the cells indicated that strain ATCC 33384 seems to be highly fragile compared to strain ATCC 29522 (data not shown).
FIG. 4.

Electron micrographs showing immunogold detection of GroEL-like protein in ultrathin sections of A. actinomycetemcomitans at 35°C (A) or 43°C (B). Sections were probed with an anti-GroEL-like protein followed by a 10-nm gold–anti-rabbit IgG conjugate. Stressed cells (B) exhibited many more gold particles both inside and outside the cells. The background level of immunogold labeling was evaluated with a rabbit anti-pig IgG (C) or without any first antibody (D), and little or no labeling was found. Immunogold detection of DnaK-like protein in ultrathin sections of A. actinomycetemcomitans at 35°C (E) or 43°C (F) indicated that stressed cells (F) showed more internal labeling than unstressed cells, but little or no labeling was seen outside the cells. In this case, sections were probed with an anti-DnaK followed by a 10-nm gold–anti-rabbit IgG conjugate. Bar = 100 nm.
SAMs of strains ATCC 33384 and ATCC 29522 were separated through different-pore-size filters, and both denatured and native GroEL-like proteins were detected by Western immunoblot analysis in the SAMs of both strains. Moreover, typical tetradecameric forms of native GroEL-like protein were also observed by TEM in the SAMs of both strains.
Effects of the native GroEL-like protein on epithelial cells.
Effects of Hsp on epithelial growth were examined in cultures of periodontal ligament epithelial cells, which resemble junctional epithelial cells in morphology, cytokeratin profile, and adhesion molecules (32). The GroEL-like protein increased the proliferation of periodontal ligament epithelial cells at concentrations ranging from 0.4 to 1 μg/ml. At higher concentrations, the number of cells in the cultures decreased significantly (Fig. 5). On the other hand, the cell viability assay using HaCaT epithelial cells indicated that the GroEL-like protein was strongly toxic for the cells. Even at low concentrations, where no change in cell numbers was observed, cell viability was clearly reduced. After treatment with 4 μg of the GroEL-like protein/ml, 90% of the cells were dead (Table 1). Experiments on the kinetics of the cytotoxic effect indicated that at this concentration of the protein, the viability of HaCaT cells decreased at a linear rate up to 4 days of culture. Essentially identical results were obtained when the HaCaT cells were cultured in serum-free KGM medium, indicating that the effect is not mediated by serum factors (data not shown).
FIG. 5.

Effects of native GroEL-like protein on numbers of cells in cultures of periodontal ligament epithelial cells. Cells were first cultured for 48 h, and then culture was continued in the presence of different concentrations of the GroEL-like protein for 7 days. The numbers of cells in the cultures were measured by staining with crystal violet and measuring the OD570 of the dissolved stain. Values are means plus standard deviations for four samples. Differences between the control and the GroEL-like protein at 0.4 to 4.0 μg/ml were statistically significant (P < 0.05 by Sheffe’s F test).
TABLE 1.
Effects of native GroEL-like protein on cell numbers and viability in cultures of HaCaT epithelial cellsa
| Concn of GroEL-like protein (μg/ml) | No. of cells (% of control)b | Viability (% of control)b |
|---|---|---|
| 0 | 100 ± 9.4 | 100 ± 3.2 |
| 0.40 | 62.1 ± 10.8* | |
| 0.50 | 101 ± 4.9 | 57.7 ± 7.5* |
| 0.67 | 99.0 ± 1.9 | 55.6 ± 8.6* |
| 1.00 | 95.8 ± 6.9 | 56.0 ± 6.1* |
| 2.00 | 74.3 ± 1.4* | 42.9 ± 6.3* |
| 4.00 | 59.5 ± 3.2* | 10.3 ± 3.0* |
Cells were first cultured for 24 h, and then culturing was continued in the presence of different concentrations of Hsp for 4 days. The numbers of cells in the cultures were measured by staining with crystal violet and measuring the OD570 of the dissolved stain. Cell viability was measured in a separate set of cultures by using a tetrazolium-formazan assay.
Values are means ± standard deviations for four samples. Values marked with asterisks are statistically significantly different from control values (P < 0.01 by Sheffe’s F test).
DISCUSSION
While the GroEL-like protein is generally considered a cytoplasmic protein in most prokaryotes, there have been reports suggesting that this protein may also be membrane associated in some organisms (1, 34, 36). Cellular pathways used by Hsp’s when secreted from the bacterial cytoplasm are not well understood, and we should not exclude the possibility of leakage of the GroEL-like protein during experimental manipulation, with subsequent binding to the bacterial cell surface. We tried to detect the GroEL-like protein in different cellular compartments of A. actinomycetemcomitans ATCC 29522 by cell fractionation and Western immunoblot analysis using antibodies specific for the GroEL-like protein. Although cell fractionation probably yields a fair amount of cross-contamination between fractions, the Western blot results indicated that the GroEL-like protein from A. actinomycetemcomitans may be bound to the external membranes of the microorganism in both the outer-membrane fraction and vesicles. In contrast, DnaK antibody detected reactive material only in the cytoplasmic fraction.
Electron microscopy analysis indicated that the GroEL-like protein of A. actinomycetemcomitans is present both inside (in cytoplasm, cytoplasmic membrane, and periplasm) and outside the cells and is more likely to be within the extracellular material surrounding the bacteria than on the outer membrane. This contrasts with results obtained with other bacteria, where GroEL-like proteins were mostly found in the cytoplasm or sometimes associated with membranes (8, 34, 36). Our electron microscopy data also indicate that, even in the absence of stress, the bacterial cells produce a detectable amount of the GroEL-like protein. This was also shown in mycobacterial species (8). The external localization of GroEL-like protein in A. actinomycetemcomitans ATCC 29522 appears not to involve bacterial autolysis as in the case of H. pylori (33) because (i) the examined cells were harvested and heat stressed in mid-log phase, (ii) the DnaK-like protein was localized mostly inside the cells (both by Western immunoblot analysis of cell fractions and by electron microscopy) and can thus be used as an internal marker, and (iii) DNA was not found outside the bacterial cells (as opposed to strain ATCC 33384). Studies of major cell envelope proteins of oral strains of A. actinomycetemcomitans indicated that four or five proteins could be identified, but none of them resembled the GroEL-like protein (4, 27). Recent experimental evidence has indicated that in Legionella pneumophila, Hsp60 was predominantly associated with the cell envelope (12). It was also shown that heat-shocked cells exhibited decreased levels of cell-associated Hsp60 and increased levels of surface epitopes, suggesting that the stress protein was released by stressed bacteria.
The molecular structure of the GroEL-like protein was also investigated in the SAMs of A. actinomycetemcomitans ATCC 29522 and ATCC 33384 by using specific molecular weight filtration fractions and immunoelectron microscopy preparations. The SAM of A. actinomycetemcomitans ATCC 33384 contained the typical tetradecameric forms of the protein. It should be noted that these results were obtained when CHAPS was included in the samples. When the zwitterionic detergent was not included, no GroEL-like particles could be seen. Very little is known about how the GroEL-like protein is exported outside the cells and about the mechanism by which the protein becomes associated with the external material of A. actinomycetemcomitans. Since cytoplasmic, periplasmic, and extracellular forms of the GroEL-like protein are found, either the putative export mechanism must have the ability to export a large protein (at least 896 kDa, i.e., 14 times 64 kDa) which may be active or A. actinomycetemcomitans has the ability to assemble the protein in all cellular compartments. Bacterial extracytoplasmic Hsp’s may play different roles in pathogenicity. Ubiquitin-like epitopes of Candida albicans modulate the interaction between cells and host structures (35), fimbrial chaperones cooperate to assemble fimbrial structures (39), and the GroEL-like protein from H. ducreyi is involved in binding to eukaryotic cells (11).
The present observations indicate that the GroEL-like protein is not an intrinsic part of the surface of A. actinomycetemcomitans, since very little protein can be visualized on the cell surface by electron microscopy. The stress protein seems to be secreted but is not bound to cell envelopes. A recent study showed that a SAM from A. actinomycetemcomitans ATCC 33384 contained a 62-kDa protein (GroEL homologue) capable of bone-resorbing activity (19). Because we found that the GroEL-like protein of A. actinomycetemcomitans is localized partially on the surface of the bacteria and has the potential to be released extracellularly, we tested its effects on epithelial cells, which are the primary targets of the subgingival plaque bacteria. Because both unkeratinized and keratinized epithelial cells are present in the gingiva, we selected two cell types that have properties similar to those of epithelial cells found in the gingiva. The protein was found to have two interesting properties. First, it increases cell proliferation at low concentrations. Because increased proliferation of gingival pocket epithelium is a major mechanism of pocket formation and possibly an initial inflammatory response to pathogenic bacteria, this may be a significant finding. Secondly, the GroEL-like protein has toxic effects on the epithelial cells. Therefore, depending on the A. actinomycetemcomitans concentration in different areas of the pocket, both cell death and increased proliferation may be the consequences of this stress protein. Our study supports the toxic activity of this Hsp, as we demonstrated that native preparations of the protein have a significant cytotoxic effect on epithelial cells grown in vitro. In a previous study, Meghji et al. (25) showed the highly cytotoxic activity of SAMs from A. actinomycetemcomitans ATCC 33384 and Y4 at concentrations as low as 1 ng/ml. The active component was a protein- and carbohydrate-containing material, but not the leukotoxin or the lipopolysaccharides. The same group found that a small protein found in the SAM produced a dose-dependent inhibition of labeled thymidine incorporation by an osteoblast-like cell line (41). They also found that 9 of 16 sera from LJP patients significantly neutralized the antiproliferative activity, while sera from normal subjects could not.
Preliminary data that we have indicate that the GroEL-like protein is recognized by sera of LJP patients (data not shown). These conclusions have also been reached by Koga et al. (20) and Kirby et al. (19), studying small sets of samples. The external localization of the GroEL-like protein may explain its immunogenicity and support its pathogenic role. The concept that the GroEL-like protein may be secreted by adhering and intracellular A. actinomycetemcomitans is interesting. Kakeda and Ishikawa (18) showed that bacterial endosymbionts overproduced and released a GroEL-like protein known as symbionin, which is believed to play an important role in maintaining the endosymbiosis state. Garduño et al. (12) have recently proposed that, along with its putative stress-alleviating functions, Hsp60 may play a key role in supporting the intracellular lifestyle of L. pneumophila. In fact, the same group suggested that host cell contact and internalization are requirements for synthesis of Hsp60 by L. pneumophila (9). A similar mechanism could take place when cells of A. actinomycetemcomitans invade host cells. In summary, the GroEL-like protein may be a major virulence factor of A. actinomycetemcomitans and could play an important role in the pathophysiological mechanism of LJP.
ACKNOWLEDGMENTS
This work was supported in part by grants from the Medical Research Council of Canada, the Réseau de Recherche en Santé Bucco-Dentaire du Fonds de Recherche en Santé du Québec, the Fonds Emile-Beaulieu, and the Laboratoire de Contrôle Microbiologique de l’Université Laval, and by a Grant-in-aid for Scientific Research from the Ministry of Education, Science, and Culture of Japan.
REFERENCES
- 1.Blander S J, Horowitz M A. Major cytoplasmic membrane protein of Legionella pneumophila, a genus common antigen and member of the hsp 60 family of heat shock proteins, induces protective immunity in a guinea pig model of Legionnaires’ disease. J Clin Investig. 1993;91:717–723. doi: 10.1172/JCI116253. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Boukamp P, Petrussevska R T, Breitkleutz D, Hornung J, Markham A, Fusenig N E. Normal keratinization in a spontaneously immortalized aneuploid human keratinocyte cell line. J Cell Biol. 1988;106:761–771. doi: 10.1083/jcb.106.3.761. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Brunette D M, Melcher A H, Moe H K. Culture and origin of epithelium-like and fibroblast-like cells from porcine periodontal ligament explants and cell suspensions. Arch Oral Biol. 1976;21:393–400. doi: 10.1016/0003-9969(76)90001-7. [DOI] [PubMed] [Google Scholar]
- 4.Di Rienzo J M, Spieler E L. Identification and characterization of the major cell envelope proteins of oral strains of Actinobacillus actinomycetemcomitans. Infect Immun. 1983;39:253–261. doi: 10.1128/iai.39.1.253-261.1983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Ebersole J L, Cappelli D, Steffen M J. Longitudinal dynamics of infection and serum antibody in A. actinomycetemcomitans periodontitis. Oral Dis. 1995;1:129–138. doi: 10.1111/j.1601-0825.1995.tb00175.x. [DOI] [PubMed] [Google Scholar]
- 6.Ellis R J. Stress proteins as molecular chaperones. In: van Eden W, Young D B, editors. Stress proteins in medicine. New York, N.Y: Marcel Dekker, Inc.; 1996. pp. 1–26. [Google Scholar]
- 7.Ensgraber M, Loos M. A 66-kilodalton heat shock protein of Salmonella typhimurium is responsible for binding of the bacterium to intestinal mucus. Infect Immun. 1992;60:3072–3078. doi: 10.1128/iai.60.8.3072-3078.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Esaguy N, Aguas A P. Subcellular localization of the 65-kDa heat shock protein in mycobacteria by immunoblotting and immunogold ultracytochemistry. J Submicrosc Cytol Pathol. 1997;29:85–90. [PubMed] [Google Scholar]
- 9.Fernandez R C, Logan S M, Lee S H S, Hoffman P S. Elevated levels of Legionella pneumophila stress protein Hsp60 early in infection of human monocytes and L929 cells correlate with virulence. Infect Immun. 1996;64:1968–1976. doi: 10.1128/iai.64.6.1968-1976.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Flynn G C, Campbell T G, Rothman J E. Peptide binding and release by proteins implicated as catalysts of protein assembly. Science. 1989;245:385–390. doi: 10.1126/science.2756425. [DOI] [PubMed] [Google Scholar]
- 11.Frisk A, Ison C A, Lagergard T. GroEL heat shock protein of Haemophilus ducreyi: association with cell surface and capacity to bind to eukaryotic cells. Infect Immun. 1998;66:1252–1257. doi: 10.1128/iai.66.3.1252-1257.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Garduño R A, Faulkner G, Trevors M A, Vats N, Hoffman P S. Immunolocalization of Hsp60 in Legionella pneumophila. J Bacteriol. 1998;180:505–513. doi: 10.1128/jb.180.3.505-513.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Gaston J S H, Life P F, Bailey L C, Bacon P A. In vitro responses to a 65-kilodalton mycobacterial protein by synovial T cells from inflammatory arthritis patients. J Immunol. 1989;143:1289–1295. [PubMed] [Google Scholar]
- 14.Gillis T P, Miller R A, Young D B, Khanolkar S R, Buchanan T M. Immunochemical characterization of a protein associated with Mycobacterium leprae cell wall. Infect Immun. 1985;49:371–377. doi: 10.1128/iai.49.2.371-377.1985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Gomez F J, Allendoerfer R, Deepe G S., Jr Vaccination with recombinant heat shock protein 60 from Histoplasma capsulatum protects mice against pulmonary histoplasmosis. Infect Immun. 1995;63:2587–2595. doi: 10.1128/iai.63.7.2587-2595.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Hansen K, Bangsborg J M, Fjordvang H, Pederson N S, Hindersson P. Immunochemical characterization of and isolation of the gene for Borrelia burgdorferi immunodominant 60-kilodalton antigen common to a wide range of bacteria. Infect Immun. 1988;56:2047–2053. doi: 10.1128/iai.56.8.2047-2053.1988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Hinode D, Grenier D, Mayrand D. A general procedure for the isolation of heat-shock proteins from periodontopathogenic bacteria. J Microbiol Methods. 1996;25:349–355. [Google Scholar]
- 18.Kakeda K, Ishikawa H. Molecular chaperon produced by an intracellular symbiont. J Biochem. 1991;110:583–587. doi: 10.1093/oxfordjournals.jbchem.a123623. [DOI] [PubMed] [Google Scholar]
- 19.Kirby A C, Meghji S, Nair S P, White P, Reddi K, Nishihara T, Nakashima K, Willis A C, Sim R, Wilson M, Henderson B. The potent bone-resorbing mediator of Actinobacillus actinomycetemcomitans is homologous to the molecular chaperone GroEL. J Clin Investig. 1995;96:1185–1194. doi: 10.1172/JCI118150. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Koga T, Kusuzaki T, Asakawa H, Senpuku H, Nishihara T, Noguchi T. The 64-kilodalton GroEL-like protein of Actinobacillus actinomycetemcomitans. J Periodontal Res. 1993;28:475–477. doi: 10.1111/j.1600-0765.1993.tb02107.x. [DOI] [PubMed] [Google Scholar]
- 21.Laemmli U K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature. 1970;227:680–685. doi: 10.1038/227680a0. [DOI] [PubMed] [Google Scholar]
- 22.Lathigra R B, Butcher P D, Garbe T R, Young D B. Heat-shock proteins as virulence factors of pathogens. Curr Top Microbiol Immunol. 1991;167:125–143. doi: 10.1007/978-3-642-75875-1_8. [DOI] [PubMed] [Google Scholar]
- 23.Løkensgard I, Bakken V, Schenck K. Heat shock response in Actinobacillus actinomycetemcomitans. FEMS Immunol Med Microbiol. 1994;8:321–328. doi: 10.1111/j.1574-695X.1994.tb00459.x. [DOI] [PubMed] [Google Scholar]
- 24.Lowry O H, Rosebrough N Y, Farr A L, Randall R J. Protein measurement with the Folin phenol reagent. J Biol Chem. 1951;193:265–275. [PubMed] [Google Scholar]
- 25.Meghji S, Wilson M, Henderson B, Kinane D. Antiproliferative and cytotoxic activity of surface-associated material from periodontopathogenic bacteria. Arch Oral Biol. 1992;37:637–644. doi: 10.1016/0003-9969(92)90126-s. [DOI] [PubMed] [Google Scholar]
- 26.Meyer D H, Fives-Taylor P M. The role of Actinobacillus actinomycetemcomitans in the pathogenesis of periodontal disease. Trends Microbiol. 1997;5:224–228. doi: 10.1016/S0966-842X(97)01055-X. [DOI] [PubMed] [Google Scholar]
- 27.Miller D, Poolman J T, Bernadina W E, van Kol P J, Ruitenberg E J. Characterization of outer membrane proteins from Actinobacillus actinomycetemcomitans. Microb Pathog. 1990;9:227–233. doi: 10.1016/0882-4010(90)90011-e. [DOI] [PubMed] [Google Scholar]
- 28.Multhoff G, Hightower L E. Cell surface expression of heat shock proteins and the immune response. Cell Stress Chaperones. 1996;1:167–176. doi: 10.1379/1466-1268(1996)001<0167:cseohs>2.3.co;2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Nakagawa S, Machida Y, Nakagawa T, Fujii H, Yamada S, Takazoe I, Okuda K. Infection by Porphyromonas gingivalis and Actinobacillus actinomycetemcomitans, and antibody responses at different ages in humans. J Periodontal Res. 1994;29:9–16. doi: 10.1111/j.1600-0765.1994.tb01085.x. [DOI] [PubMed] [Google Scholar]
- 30.Nakano Y, Inai Y, Yamashita Y, Nagaoka S, Kusuzaki-Nagira T, Nishihara T, Okahashi N, Koga T. Molecular and immunological characterization of a 64-kDa protein of Actinobacillus actinomycetemcomitans. Oral Microbiol Immunol. 1995;10:151–159. doi: 10.1111/j.1399-302x.1995.tb00136.x. [DOI] [PubMed] [Google Scholar]
- 31.O’Farrell P H. High-resolution two-dimensional electrophoresis of proteins. J Biol Chem. 1975;250:4007–4021. [PMC free article] [PubMed] [Google Scholar]
- 32.Pan Y-M, Filth J D, Salonen J, Uitto V-J. Multilayer culture of periodontal ligament epithelial cells: a model for junctional epithelium. J Periodontal Res. 1994;30:97–107. doi: 10.1111/j.1600-0765.1995.tb01258.x. [DOI] [PubMed] [Google Scholar]
- 33.Phadnis S H, Parlow M H, Levy M, Ilver D, Caulkins C M, Connors J B, Dunn B E. Surface localization of Helicobacter pylori urease and a heat shock protein homolog requires bacterial autolysis. Infect Immun. 1996;64:905–912. doi: 10.1128/iai.64.3.905-912.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Scorpio A, Johnson P, Laquerre A, Nelson D R. Subcellular localization and chaperone activities of Borrelia burgdorferi Hsp60 and Hsp70. J Bacteriol. 1994;176:6449–6456. doi: 10.1128/jb.176.21.6449-6456.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Sepulveda P, Lopez-Ribot J L, Gozalbo D, Cervera A, Martinez J P, LaJean Chaffin W. Ubiquitin-like epitopes associated with Candida albicans cell surface receptors. Infect Immun. 1996;64:4406–4408. doi: 10.1128/iai.64.10.4406-4408.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Taguchi H, Yamaguchi H, Yamamoto T, Kamiya S. Immunocytochemical localization of 60-kDa heat shock protein in Vibrio cholerae. Zentbl Bakteriol. 1996;284:496–500. doi: 10.1016/s0934-8840(96)80002-8. [DOI] [PubMed] [Google Scholar]
- 37.Tilly K, Georgopoulos C. Evidence that the two Escherichia coli groE morphogenetic gene products interact in vivo. J Bacteriol. 1982;149:1082–1088. doi: 10.1128/jb.149.3.1082-1088.1982. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Tolo K, Helgeland K. Fc-binding components: a virulence factor in Actinobacillus actinomycetemcomitans? Oral Microbiol Immunol. 1991;6:373–377. doi: 10.1111/j.1399-302x.1991.tb00509.x. [DOI] [PubMed] [Google Scholar]
- 39.van Rosmalen M, Saier M H., Jr Structural and evolutionary relationships between two families of bacterial extracytoplasmic chaperone proteins which function cooperatively in fimbrial assembly. Res Microbiol. 1993;144:507–527. doi: 10.1016/0923-2508(93)90001-i. [DOI] [PubMed] [Google Scholar]
- 40.Watson K. Microbial stress proteins. Adv Microbiol Physiol. 1990;31:183–223. doi: 10.1016/s0065-2911(08)60122-8. [DOI] [PubMed] [Google Scholar]
- 41.White P A, Wilson M, Nair S P, Kirby A C, Reddi K, Henderson B. Characterization of an antiproliferative surface-associated protein from Actinobacillus actinomycetemcomitans which can be neutralized by sera from a proportion of patients with localized juvenile periodontitis. Infect Immun. 1995;63:2612–2618. doi: 10.1128/iai.63.7.2612-2618.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Wilson M, Henderson B. Virulence factors of Actinobacillus actinomycetemcomitans relevant to the pathogenesis of inflammatory periodontal diseases. FEMS Microbiol Rev. 1995;17:365–379. doi: 10.1111/j.1574-6976.1995.tb00220.x. [DOI] [PubMed] [Google Scholar]