

Summary
Eastern North American migratory monarch butterflies exhibit migratory behavioral states in fall and spring characterized by sun-dependent oriented flight. However, it is unclear how monarchs transition between these behavioral states at their overwintering site. Using a modified Mouritsen-Frost flight simulator, we confirm individual directionality and compass-based orientation (leading to group orientation) in fall migrants, and also uncover sustained flight propensity and direction-based flight reinforcement as distinctly migratory behavioral traits. By testing monarchs at their Mexican overwintering sites, we show that overwintering monarchs show reduced propensity for sustained flight and lose individual directionality, leading to the loss of group-level orientation. Overwintering fliers orient axially in a time-of-day dependent manner, which may indicate local versus long-distance directional heading. These results support a model of migratory flight behavior in which modular, state-dependent switches for flight propensity and orientation control are highly dynamic and are controlled in season- and location-dependent manners.
Subject areas: Ecology, Entomology, Evolutionary biology
Graphical abstract
Highlights
-
•
First individual flight behavior measurements at the Mexican overwintering sites
-
•
Monarch migratory flight behavior comprises multiple components
-
•
Sustained flight and southward direction temporarily cease at overwintering site
-
•
Flight simulator captures daily movements reported at overwintering clusters
Ecology; Entomology; Evolutionary biology
Introduction
Animal migrations have evolved to increase survival in adverse conditions and maximize fitness in heterogenous environments. Migrations are initiated, maintained, and terminated through carefully regulated processes that are time sensitive and environmentally modulated (reviewed in a study by Ramenofsky and Wingfield).1 A suite of behavioral, physiological, morphological, and metabolic traits differ dramatically between migratory and non-migratory (e.g., breeding) states and are often conflicting, such as highly mobile, social migrants versus sedentary, territorial non-migrants.2 Transitioning between migratory states requires system-wide coordination on different scales—from specific cellular changes to the development of entire organ systems—that are often mediated by differential hormone activity (reviewed in studies by Rankin, Sharma et al., and Cornelius et al.)3,4,5 or epigenetic mechanisms.6,7,8 Migration initiation involves integrating intrinsic (e.g., circadian and circannual timers) and extrinsic (e.g., environmental, social) factors in vertebrate (reviewed in a study by Cornelius et al)5 and invertebrate (reviewed in a study by Chapman et al.)9 migrants. The mechanisms of termination, on the other hand, are much less well understood.2
In contrast to initiation where predictive environmental cues (e.g., photoperiod or temperature) are sufficient for induction, termination uniquely involves spatial specificity that requires spatial recognition and resource evaluation (i.e., individuals will terminate migration at specific locations that ultimately maximize their fitness).5,10 Hormonal control of the transition from migratory to breeding physiology (also called “arrival biology” in migratory birds) has been best examined in Arctic birds (reviewed in a study by Ramenofsky and Wingfield)11 and also has been studied in insects (e.g., a study by Jiang et al).12 Controllers of the spatially relevant behavioral aspects of migration termination, however, are not well known. Some long-distance migrants, such as Pacific salmon or sea turtles, imprint natal habitats using geomagnetic13,14,15,16 or olfactory17 cues, returning to natal sites to terminate migratory movements and transition life history stages. Social environment plays a large role in migration termination in low site fidelity nomadic migrants, e.g., the pine siskin.18 While ultimate evolutionary drivers of migration termination site are hypothesized,19 how migration is terminated at specific locations remains an open problem in many migratory systems. Understanding termination is important in order to identify the ultimate selective forces shaping stopover ecology20 and predict dynamics of migrations that have significant impact on human life.21
The migration of the Eastern North American (ENA) monarch butterfly (Danaus plexippus) presents an interesting case of migration termination because these butterflies have no previous experience with their overwintering sites. Therefore, their behavioral flexibility is not thought to involve learning or currently understood imprinting mechanisms. Each fall, monarch butterflies will fly up to 4,000 km from across the US and southern Canada to overwinter in the oyamel fir forests of the Trans-Mexican Volcanic Belt in central Mexico. Individuals persist in reproductive diapause until the subsequent spring when they mate and remigrate northwards to repopulate their northern range in 2–4 partially overlapping generations. The sites of monarch overwintering clusters are highly predictable, with the vast majority of individuals overwintering in the same broad groups of trees for at least the last half-century.22,23 Because no individual butterfly makes the journey more than once, overwintering site destination must involve a genetically encoded spatiotemporal program. It is hypothesized that a specific microclimate attracts monarchs to particular peaks,24,25,26 but the specific conditions of the microclimate are unclear and how these conditions terminate migratory behavior are unknown. An important first step to answering these questions is describing how migratory behavior changes at the overwintering sites.
Fall migratory monarchs are characterized by two primary adaptations that enable their annual migration: reproductive diapause and migratory flight behavior. While the termination of reproductive diapause is known to involve juvenile hormone (JH) in monarchs,27,28 transitions between migratory behavioral states are not well studied. Monarchs utilize a primary time-compensated sun compass29,30 and a secondary inclination magnetic compass31 to navigate in a south-southwestward direction. Polarized UV light is required for the initiation of migratory flight behavior.32,33 Antennal circadian clocks control the time-compensation component of oriented flight behavior.34,35 In spring, the same migratory monarchs flip their orientation and remigrate northwards due to experiencing extended periods of cold temperature.36 Direction flipping explains how individual butterflies make bidirectional migratory trips. However, the behavioral transitions that occur between the migratory and re-migratory states remain unexplained.
We sought to characterize how monarch behavior changes at the overwintering sites. Specifically, we wanted to determine if migratory behavior changes as an integrated trait or do component behavioral traits vary independently. Answering this question can provide initial insight into the physiological mechanisms that mediate migration termination at these specific sites in this species. In order to understand how different features of the monarch migratory state change along the course of the migration, particularly at their overwintering site, we designed and implemented a novel flight simulator apparatus and computer-vision based analysis to test migratory flight behavior at the Mexican overwintering sites.
Results
A modified flight simulator captures migratory flight behavior
We began our studies by utilizing a modified Mouritsen-Frost (MF) simulator to measure the two characteristic migratory flight behavior phenotypes: individual directionality and compass-directed, group-level orientation (Figure 1A). The simulator differs from the MF simulator in that monarchs are attached reversibly and non-invasively to a flexible tether (thread) and allowed to fly within a barrel with an unobstructed view of the sun37 versus being attached to a rigid tether. Flight was recorded from a camera mounted above the simulator and flight patterns (heading direction and wingbeat frequency) were analyzed using a computer vision approach (Figure 1).
Figure 1.

A modified Mouritsen-Frost flight simulator captures essential features of migratory flight
(A) Monarch butterfly flying in the modified simulator. We determined individual direction by training a model to identify the “origin” of the simulator (yellow box) and the head of the monarch (red box). The white box indicates a monarch identified in the model.
(B) Directionality as measured by weighted r for summer non-migrants, fall migrants, and overwintering migrants. Error bars indicate 1.5 times interquartile range.
(C–E) Orientation plots for all individuals tested in sunny conditions. Individual butterflies are represented by circles colored by membership score based on weighted r. The distance from the origin is given by individual r. For ease of visualization, all r values were normalized to the maximum r value measured in the study (r = 0.900). Circle size is proportional to an individual’s proportion flying time (PFT). Resultant group R∗(weighted r) vector (gray wedge) is only shown for group(s) with significant group directionality (p < 0.05). For ease of visualization, all R∗ vectors are normalized to the highest group R∗ value (R∗ = 1.472 for directional fall fliers tested in sunny conditions).
The key features of fall migratory flight are recapitulated in our apparatus. As expected from previous reports, we observed substantial variation in flight among individuals, from virtually no flight (6 s) to continuous flight over the full 10 min trial (Figure 1; Table S1). We assessed individual directionality through a “weighted r” score that accounts for the fact that flight times (the amount of time an individual was in flight during the 10 min video) differed among individuals. Weighted r is derived from multiplying the individual r value (the resultant vector of an individual’s flight) by the individual’s proportion of flight time (PFT; proportion of recorded video identified as flight). We find significant differences in individual flight directionality between groups (Figure 1B; Kruskal-Wallis: p = 2.2e-16). Fall migrants show stronger individual directionality compared to summer non-migrants (Figure 1B; Mann-Whitney: p = 0.01157).
Variation in weighted r was high among fall migrants (CV = 0.925 for fall migrants). In previous analyses of monarch directional flight, the distinction between “directional” versus “non-directional” individuals is recognized as an important exclusion criterion.38 In order to identify directional individuals in our study in an unbiased manner, we performed unsupervised clustering across all individuals tested in sunny conditions. In this analysis, (marginally) significantly more fall individuals are identified as directional (membership score (weighted r) > 0.5) compared to summer (Figures 1C and 1D; Fisher’s exact: p = 0.053): 11 of 33 (33%) fall migrants are identified as directional, whereas two of 22 (9%) summer monarchs were identified as directional.
We evaluated group orientation using Moore’s modified Rayleigh test for uniformity,39 which leverages the fact that we have both vector direction and magnitude in order to describe group directionality. The modified Rayleigh test requires ranking individuals by a weighted metric. While our main results present individuals ranked by weighted r, results were consistent across multiple different weighting factors (Table 1). Importantly, when all fall individuals that were tested in sunny conditions are considered, group-level orientation in the southwest direction is highly significant (Figure 1D; modified Rayleigh: n = 33, R∗(weighted r) = 1.337, mean vector [MV] = 229.675°; p < 0.005). As well, specifically directional fall migrants show significant group orientation to the southwest (Figure S1B; modified Rayleigh: n = 11, R∗(weighted r) = 1.427 MV = 232.712°; p < 0.005). In contrast, summer non-migrants flown in sunny conditions do not show significant group-level orientation, either as a combined group (Figure 1C; modified Rayleigh: n = 22, R∗(weighted r) = 0.319; p > 0.05) or when only directional individuals are considered (Figure S1A; modified Rayleigh: n = 2, R∗(weighted r) = 0.386; p > 0.5). Neither summer non-migrants nor fall migrants show significant group orientation under cloudy conditions (Figure S2; Table S2; modified Rayleigh R∗ (weighted r): p > 0.1 and 0.5 for summer and fall, respectively). We note that these results match the expectations from both Mouritsen and Frost,30 in which an altered UV condition used in those analyses led to no significant group orientation, and,31 in which UV-transmissive material allows for robust orientation. We learned after testing that the polyethylene material used for our cylinder has low UV-A and UV-B transmissibility. Therefore, we anticipate that there was likely interference with magnetic inclination compass ability, leading to the inability to detect southward orientation in outdoor cloudy conditions. Finally, a hallmark of monarch migratory flight is that individuals orient in the south-southwest direction independent of the time of day, a pattern that we recover in our results (Figure 2; Table S4; circular-circular regression: n = 33; p = 0.2399).
Table 1.
Rayleigh test for uniformity (R) and Modified Rayleigh test for uniformity (R∗) calculated across four different extrinsic weighting factors (weighted r, r, PFT, and TFT)
| R | R∗ (weighted r) | R∗ (r) | R∗ (PFT) | R∗ (TFT) | |
|---|---|---|---|---|---|
| Summer, sunny; n = 22 | 0.154, p = 0.597 | 0.319, p > 0.5 | 0.130, p > 0.95 | 0.487, p > 0.5 | 0.487, p > 0.5 |
| Fall, sunny; n = 33 | 0.373, 226.437°, p = 0.009 | 1.337, 229.675°, p < 0.005 | 1.309, 224.074°, p < 0.01 | 1.386, 237.997°, p < 0.005 | 1.386, 237.997°, p < 0.005 |
| Overwinter, sunny; n = 22 | 0.241, p = 0.283 | 0.847, p > 0.1 | 0.756, p > 0.1 | 0.682, p > 0.1 | 0.682, p > 0.1 |
TFT = total individual flight time. Values presented are the Rayleigh R or R∗ statistic, mean vector θ (for significant p values), p value.
Figure 2.

Time-of-day-dependent flight orientation reveals differential skylight cue processing throughout the annual cycle
(A–C) Orientation plots for all summer, fall, and overwintering monarchs tested in sunny conditions. All individuals tested in sunny conditions are included regardless of individual directionality or flight scores. Circle color indicates the time of day at which the individual was tested (ranging from ∼900 to 1,600 h). Individuals within a group were randomly tested across several days.
An expanded description of migratory flight behavior
Increased flight capacity is characteristic of insect migrants (e.g., studies by Zhang et al. and Coats et al.).40,41 Circumstantial evidence suggests that flight duration positively correlates with migration status in monarchs. ENA monarchs fly for longer duration and distance (as measured by flight mill assays) compared to Western North American (WNA) monarchs,42 which undertake a substantially shorter migration. “Redder” migratory monarchs fly greater distances in a flight mill compared to stronger-hued, non-migratory monarchs.43 Here we directly demonstrate that fall migrants tend to fly for longer duration compared to summer non-migrants in either sunny (Figure 3A; Wilcoxon rank sum: p = 0.0002) or cloudy (Figure S3B; Wilcoxon rank sum: p = 0.0020) conditions as determined by PFT. Fall migrants show high sustained flight propensity in cloudy conditions as well (Figure S3B), indicating that sustained flight does not require sunny conditions. This matches the observation and expectation that monarchs undertake migratory flights on cloudy days.
Figure 3.

Sustained flight and orientation-motivated flight are additional components of migratory flight behavior
(A) Proportion flying time (PFT) for summer non-migrants, fall migrants, and overwintering migrants tested in sunny conditions. ∗p < 0.05. Boxplots are as in Figure 1.
(B) Weighted r versus theta for fall migrants tested in sunny conditions. Grey shading represents 95% confidence interval.
(C–E) Orientation plots for all individuals tested in sunny conditions. Individual butterflies are represented by circles colored by membership score based on PFT.
The distinction between “sustained” versus “trivial” fliers has been recognized in at least one other insect migrant (western corn rootworm);44 variation in flight intensity among migrants has been reported in a number of species (reviewed in a study by Naranjo).45 We performed an unbiased clustering analysis across all tested individuals (including both sunny and cloudy conditions) in order to identify sustained fliers in an unbiased manner (Figures 3C–3E and S1D–S1F). In this analysis, considering only individuals flow in sunny conditions, 18 of 33 (54.5%) fall migrants are identified as “sustained” fliers, which is significantly greater than the 4 of 22 (18.2%) summer monarchs identified as sustained fliers (Figures S1D and S1E; Fisher’s exact: p = 0.011). When individuals flown in either sunny or cloudy conditions are considered, 37 of 56 (66%) fall migrants are identified as sustained fliers, which is significantly greater than the 5 of 30 (16.7%) summer monarchs identified as sustained fliers (Figures 3C–3E, S2A, and S2B; Fisher’s exact: p < 0.0001). Of the 18 fall migratory sustained fliers flown in sunny conditions, 11 (61%) were directional. Sustained flying fall migrants tested in sunny conditions show significant group orientation (Table S5; modified Rayleigh: n = 18, R∗(weighted r) = 1.472, MV = 237.536, p < 0.005).
Fall migratory monarchs appear to fly with stronger directionality when oriented in a southwest direction compared to other directions (Figure 3B; p = 0.249). Although not reaching the statistical significance threshold, likely due to low sample size, we note that nine of the ten most directional fall migratory individuals flew in a somewhat southerly direction, and six of ten flew specifically southwest (Figure 3B). Directionality was uncorrelated to theta in summer non-migrants (Figure S4; p = 0.796). These results imply that southwestward heading, specifically, is reinforced in fall migratory monarchs. This finding is consistent with the expectation that migratory monarchs maintain specific flight heading in a “balanced state” as proposed by Shlizerman et al.46 in their neuronal integration model, and with the recent discovery of goal-directed neurons in monarchs.47
Taken together, migratory flight is characterized by a suite of traits: (1) individual directionality (weighted r here; Z elsewhere), (2) compass-directed individual orientation (which leads to group-level orientation), (3) propensity for sustained flight (as determined by classification based on PFT), and (4) direction-biased flight reinforcement. Clustering analysis indicates that fall migrants may be heterogeneous for directionality and flight propensity. Previous studies have documented such phenotypic variability in directional flight assays,31 but presumably attribute this variation to experimental noise and exclude these individuals. We note that we are still able to recover characteristic migratory behavioral traits while not discarding data from these individuals.
Sustained flight behavior temporarily ceases at Mexican overwintering sites
We next sought to determine how migratory flight behavior changes when monarchs have reached their overwintering destination. All simulator-based directional flight tests thus far have occurred where monarchs are far away from the overwintering sites. The farthest south any directional flight testing has been reported is by Guerra and Reppert who tested northward flying remigrants in Seadrift, Texas (28°42′N, 96°71′W), which is 1,369 km north of the primary Mexican overwintering sites.36 Directions of southward migrants were recorded by disappearance bearing in central Mexico along the Sierra Madre Oriental by Bill Calvert in fall 1977 and 1978.48 We tested individuals (in sunny conditions only) from the primary overwintering cluster at the Sierra Chincua site in December 2021. Significantly fewer overwintering monarchs (1 out of 22, 4.54%) are classified as sustained fliers in the clustering analysis compared to fall migrants (37/56 [66%] overall; 18/33 [54.5%] in sunny conditions) (Table S5; Fisher’s exact: p = 0.0001; comparison is to fall migrants flow in sunny conditions). Since migrants cluster as strong or trivial fliers, we interpret these results as a significant shift in the frequency of strong fliers among overwintering monarchs, indicating a reduced propensity for sustained, directional flight at the overwintering site. While monarchs may reduce sustained flight propensity at the overwintering site, they must eventually revert back to sustained flight behavior because northward spring remigrants show sustained flight propensity similar to fall migrants.36 Therefore, one switch of monarch migratory flight behavior is sustained flight.
Directional flight control switching reveals multi-state orientation control
Overwintering monarchs are overwhelmingly not individually directional. Only two individuals of 22 (9.1%) are identified as strongly directional through clustering analysis (Figures 1E and S1C). As such, overwintering monarchs do not show significant consistent group orientation as do fall migrants in Michigan (Figure 1E; Table 1; modified Rayleigh: n = 2; R∗ (weighted r) = 0.847; p > 0.1). However, as a group, there is evidence of a tendency to orient axially in either northwest or southeast directions as indicated by bimodality analyses (Figure 2; Table S3, homogeneous symmetric bimodal model: q1 = 305.329°, k1 = 3.172, q2 = 125.306°, k2 = 3.172, AIC = 73.463). The best performing models for the overwintering data are all bimodal models; unimodal models are significantly poorer fits to these data, unlike other groups (Table S3). As well, there is time-of-day dependent flight orientation among all overwintering monarchs (Figure 2C; Table S4, κ = 2.0825, p = 0.0249), indicating that axial flight direction may be time-dependent.
Interestingly, the two directional overwintering individuals oriented toward the northwest (Figure 3E), which is the same direction of the larger cluster from which the tested individuals was derived (approximately 750 m away). This raises the possibility that these individuals may have been orienting with respect to the overwintering cluster. As well, despite the low frequency of strong directionality among overwintering individuals, directionality appears to be maximized in the northwest and southeast directions (Figure S4), and direction (theta) may be a reasonable predictor of weighted r in this group (Figure S4; p = 0.148, 54% deviance explained). Monarchs undertake daily directed flights to and from the main clusters at the overwintering sites called butterfly “streams” or “rivers” that often involve many thousands of individuals (studies by Calvert and Brower, and Sánchez-Tlacuahuac et al;49,50 personal observation). It is suspected that these flights serve as opportunities for monarchs to drink water from nearby streams, moisture patches and reservoirs in order to metabolize lipid reserves.49,50 It is also possible that these movements reflect flights to nectar sources. While nectaring at the overwintering sites has previously been considered negligible due to the insufficiency of available nectar resources for the large concentration of butterflies,51 monarchs do indeed nectar on a variety of plants at and near the overwintering sites.50 This behavior corroborates the conclusion that local directional, time-of-day dependent flight behavior is maintained at the overwintering site. Some migratory Palearctic bird species are known to make facultative movements during winter to identify favorable breeding sites52 or other environments or resources.53
Altogether, these results confirm that southwest directional flight observed in fall migrants ceases at the overwintering sites. Directional tuning is dynamic and can occupy at least three different states: (1) “undirected” heading (summer non-migrants), (2) long-range, constant heading (fall migrants), and (3) local time-of-day-informed heading (overwinterers).
The fact that overwintering monarchs do not show significant group directionality, yet spring remigrants fly robustly in a northeast orientation,36 indicates that there is at least a transient period at the overwintering sites during which monarchs suspend time-independent, compass-directed orientation. What controls switching between these states? Although extended cold temperature is sufficient to switch direction of compass-based orientation,36 a different mechanism must be responsible for switching between time-of-day and compass-based orientation. Previous work has shown that migratory monarchs with disrupted antennae display axial orientation in the same directions that we observe here.34,36 It was hypothesized that butterflies with black-painted antennae orient axially due to interaction between entrained brain and free-running antennal circadian clocks.34 Following this hypothesis, we predict that the interaction and relative prioritization of antennal and brain circadian clocks determines the switch between local and long-distance orienting behavior (Figure 4). Local orientation (time-of-day informed heading) relies on entrained brain clocks and operates independently of antennal circadian clocks, whereas constant heading during long-distance flight requires synchronization of antennal and brain clocks. Beetz et al.54 demonstrated a neuronal basis for switching between a sun-bearing network to a global compass coding heading-direction network during quiescence versus steering flight, respectively. It will be important to determine if heading-direction network remapping is equally dynamic between seasonal generations or sustained versus local flight states.
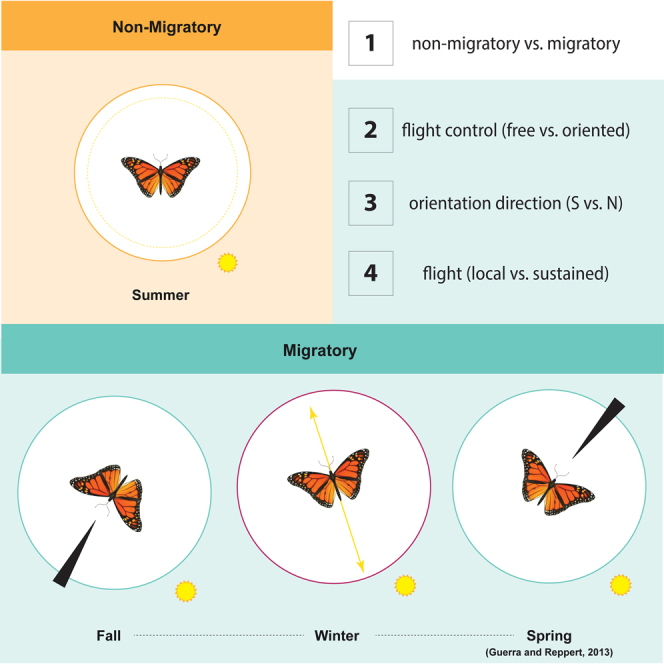
Figure 4.

A model for modular dynamic switches that control migratory flight behavior
Four distinct switches characterize differences between monarch behavioral states. Switch 1: seasonal plasticity generates spring/fall non-migratory versus fall migratory monarchs. Switch 2: differential processing of skylight cues leads to time-of-day dependent versus independent (or oriented) flight. Switch 3: Specific orientation direction, in both time-of-day dependent (sun-directed versus axial) and independent (southwest versus northeast) flight. Switch 4: sustained versus local flight behavior.
Discussion
In all, we demonstrated that a modified MF flight simulator allows for increased behavioral freedom can successfully detect characteristic migratory and non-migratory flight behaviors (individual directionality and group orientation), as well as identify additional features of migratory flight including sustained flight and orientation-motivated flight. By testing monarchs at their Mexican overwintering sites for the first time, we provide some additional insight into the mechanisms of monarch arrival biology. We show that monarchs reduce propensity for sustained flight behavior and change how they interpret skylight cues, from time-independent, unidirectional compass-oriented flight to time-dependent, axial flight, upon reaching their overwintering sites. We note, however, that we do not directly measure the transition to the termination state in this study, which would require measuring monarchs earlier in the winter season as they initially arrive to the overwintering sites. Nevertheless, decomposing migratory behavior into separable components presents the opportunity to identify how distinct environmental and behavioral inputs control specific aspects of migratory flight behavior, as well as uncover the genetic and cellular bases behind these controls.
Our results support a model of a dynamic, modular migratory behavioral phenotype (Figure 4). Long-distance migratory flight requires dynamic changes that may be described in four “switches.” First, seasonal developmental plasticity yields spring/summer non-migratory monarchs that perform fixed, local orienting behaviors driven by immediate cues (e.g., phototactic or motion-detecting flights) versus fall migratory monarchs that exhibit more flexible, state-dependent orientation and flight behaviors. Sun compass brain regions of fall migratory monarchs appear to undergo experience-dependent neuroplasticity,55 demonstrating this increased plastic potential. Next, skylight cues can be differently used to direct movement, either in a single consistent direction or in time-of-day-dependent patterns. Third, specific orientation direction is environmentally programmable. In addition to general southwest orientation in fall29,30 and northeast orientation in spring (from a study by Guerra and Reppert),36 group-level orientation changes along the southward migratory route, which is predicted to allow monarchs to navigate geographical barriers en route to their overwintering sites.48 Finally, monarchs adopt sustained versus “local” flight modes, which may reflect differences not only in flight motivation, but also in how energy resources are metabolized during flight. Flight metabolism is important for distinguishing migratory and non-migratory monarchs.56,57,58 Importantly, these switches show a degree of modularity such that monarchs can adopt different phenotypes that match the conditions and requirements of their migratory phase.
Migrating monarchs must transition between multiple states throughout their adult life cycle. Long distance flight is interspersed with local orienting decisions, such as identifying nectaring sources, roosting locations, or their ultimate overwintering locations. Moreover, migratory flight is interrupted by diapause, an extended non-migratory period, at the overwintering sites. In this way, monarch migration closely resembles the state-dependent behavioral changes of birds (reviewed in studies by Ramenofsky and Wingfield, and Terrill)11,59 and other vertebrates,60,61 as well as of Bogong moths (Agrotis infusa), which make bidirectional long-distance migrations in Australia that are interrupted by a 3- or 4-month aestivation period (reviewed in a study by Warrant et al.).62 Across these migrants, while seasonal cues (e.g., photoperiod) mediated through neurohormones often control transitions between migratory and non-migratory states, particularly migration initiation, other factors control more dynamic transitions. Density dependence (e.g., in elk,63 deer,64 and salmon65), resource limitation (reviewed in a study by Chapman et al.66 for vertebrates; and Menz et al. 67 for invertebrates), age (e.g., in salmon68), and predation vulnerability (e.g., in the common bream69) all influence individual-level migration propensity. Migration termination is typically influenced by dynamic local factors such as atmospheric condition70 or weather,71 and can also differ by intrinsic factors such as sex.72 Arrival biology in Arctic birds is known to be highly flexible such that major behavioral and reproductive transitions are enabled yet there is still flexibility to respond to environmental uncertainty.2 It may be likely that determination of spatially explicit termination sites such as wintering or breeding locations lies at the intersection of local, dynamic termination mechanisms and longer-term, seasonally and neurohormonally controlled life history transitions. Such integration mechanisms have yet to be described (although glucocorticoids are predicted to play an important role in birds5) but are important for our ability to mechanistically link habitat selection patterns (as measured by tracking studies) to process and thus predict how migratory habitats will change.73,74 Describing the specific behavioral transitions that monarchs undergo at their overwintering sites provides an important step toward developing monarch migration as a useful model to uncover these mechanisms of termination.
It is interesting to consider how monarchs acquire and subsequently control this behavioral plasticity. Hormones mediate intraindividual behavioral and physiological migratory transitions in birds,75,76,77 eels,78 fish,79 and sea turtles.80 JH signaling is a strong candidate controller of dynamic migratory flight behavior in monarchs. High JH titer is associated with migration termination in the oriental armyworm Mythimna separata.12 JH activity is positively correlated with migratory flight and flight intensity in oriental armyworms81 and milkweed bugs,82 but is negatively correlated with flight intensity in monarchs.83 Gene expression analyses in overwintering monarchs suggest that JH metabolism is dynamic throughout the winter season, involving both endogenous and environmental controls.28 Nevertheless, JH has not been directly linked to flight orientation in any insect and has been shown to be dispensable for maintenance of directional orientation in monarchs.38 It is likely that distinct mechanisms control different components of migratory behavioral plasticity. Behavioral flexibility is a well-known feature of avian migrants.
Modularity of migratory flight behavior components has implications for the evolution of long-distance migratory flight. Migratory flight behavior modularity is consistent with the long-distance navigation hypothesis posited by Mouritsen84 in which long-distance navigation comprises three distinct phases (long-distance, homing, and pinpointing phases) that each engage a unique combination of sensory modalities. If different components of migratory flight behavior are controlled by distinct physiological and genetic processes, this may provide a mechanism to rapidly diversify migration components.85
As the number of monarchs arriving at traditional overwintering sites has fallen over the past several decades (World Wildlife Federation Monarch Counts; the Xerxes Society Western Monarch Counts), understanding how monarchs make decisions to choose specific locations, and how environmental factors impact these decisions, is, indeed, a timely pursuit. Temperature, humidity, proximity to water sources, and local wind conditions within the oyamel fir forests at 2,900–3,400 m are believed to be primary determinants of a specific microclimate that is responsible for the remarkable fidelity of overwintering site location.86 Ecological niche modeling has successfully identified potential overwintering habitat,87,88 and how this area may change under different climate change models but does not delineate specific parameters of this niche. Complicating this picture, monarchs have not been found in all locations expected to satisfy these conditions; as well, migratory monarchs from different populations (e.g., WNA monarchs) are known to overwinter in different locations and conditions. The ability to test for specific behavioral changes both at the overwintering sites and in the laboratory provides an important opportunity to identify the specific overwintering site conditions that define a critical component of this iconic phenomenon.
Limitations of the study
Given several constraints of testing, this study profiles behaviors of monarchs from one specific overwintering cluster, which is one of the largest and most consistent clusters in the area. It will be important to pursue, as possible, testing of individuals from other major clusters to determine if behaviors differ between sites, as well as to understand additional aspects of flight behavior such as site fidelity. Moreover, increasing temporal resolution of testing will give a more complete picture of the dynamic behavioral transitions at the overwintering sites.
STAR★Methods
Key resources table
| REAGENT or RESOURCE | SOURCE | IDENTIFIER |
|---|---|---|
| Deposited data | ||
| Summary of orientation data | Supplementary information for this paper | N/A |
| Software and algorithms | ||
| R Software (v. 4.1.2) | R Core Team | https://www.r-project.org/ |
| Behavioral analysis software | Supplementary resource for this paper | https://github.com/greendeilab/iScience_2024_Green |
Resource availability
Lead contact
Further information and requests for resources and reagents should be directed to and will be fulfilled by the lead contact, Dr. Delbert A. Green II (dgreenii@umich.edu).
Materials availability
This study did not generate new unique reagents.
Data code and availability
-
•
All of the data used for this study are publicly available as presented in the supplemental information as of the date of publication.
-
•
All original code has been deposited at github and is publicly available as of the date of publication. URLs are listed in the key resources table.
-
•
Any additional information required to reanalyze the data reported in this paper is available from the lead contact upon request.
Experimental model and study participant details
Adult maintenance & testing
Summer adult monarch butterflies (Danaus plexippus) were wild caught on July 10th 2023 near Ann Arbor, MI and kept in an environmental chamber with summer-like conditions (15 h light cycle at constant 25°C). Summer flight tests were performed between July 17th and July 25th, 2023 in Ann Arbor, Michigan (MI). Fall migratory individuals were caught between September 9th and September 11th, 2021 during peak migration in Detroit, MI. Adults were collected into glassine envelopes and held in an environmental chamber with a 12 h light cycle, and alternating temperature (21°C day, 12°C night). Summer and fall Michigan butterflies were tested in the simulator after at least 5 days of LD entrainment in environmental chambers. Individuals were tested under sunny or cloudy conditions between September 27th and October 12th, 2021 in Ann Arbor, MI. For cloudy conditions, we required total cloud cover with no clear indication of sun position. Overwintering monarchs were collected directly from tree branches within the primary overwintering cluster at the Sierra Chincua (Mexico) monarch reserve. Butterflies were collected into glassine envelopes approximately two hours prior to testing. Individuals were acclimated to the external surroundings in a small cage for at least 30 minutes prior to testing in the simulator. Testing occurred under sunny conditions on December 10th and 11th, 2021 within the Sierra Chincua reserve approximately 750m away from the cluster collection site. No individuals in any groups were fed prior to testing.
Method details
Flight simulator design
Monarchs were tested in a flight simulator modified from the Mouritsen & Frost design.30 The simulator comprised three primary components: 1) a large plastic cylinder, 2) an attachment apparatus consisting of acrylic rods, rubber tubing, and thread and 3) an AKASO EK7000 camera affixed to a tripod for monitoring butterfly flight. These components were assembled atop a solid-base table. The cylinder was a cutout of a white, 32 gallon, Uline trash can (linear low-density polyethylene) with dimensions of 48 cm × 41 cm (diameter x height). The attachment apparatus consisted of three perpendicular acrylic rods placed on top of the cylinder. A silicon tube was attached to the end of the center perpendicular rod. At the time of testing, the tether was secured within the silicon tube and glued to a small bandage (Band-Aid) piece attached to the thorax of the individual (as described in37). No anesthetic was required for attachment. Component setup prevented potentially biasing string tension as well as allowed constant sun visibility within the simulator.
Flight simulator trials
We allowed monarchs to start and stop in our assay, recognizing that this introduces noise and ultimately reduces flight time and directionality (r) scores. In previous studies, “weak” or non-fliers were eliminated from analysis. Guerra et al. observed 31% first trial success rate (5+ min flier) in outdoor testing,36 and 92% directional fliers of those 5+min fliers.31 In contrast, we chose to keep all individuals tested that yielded a video that passed quality control. Videos under 5 min were considered “failed” videos and were not further processed. Indeed, we considered flight time a key parameter in this work. We reason that if flight propensity as measured by willingness to fly in the simulator is due to random effects or testing artifacts, the frequency of “failed” fliers should be constant across all groups. However, this is not what is observed, as given by significant differences in PFT across groups (Figure 3A).
Image acquisition
Trials were recorded with an AKASO EK7000 camera at 120 frames per second with a resolution of 1280 × 720 pixels. Videos were recorded for different lengths across groups (10, 15, and 15 min for summer, fall, and overwinterers, respectively). For analysis, fall and overwintering videos were truncated at 10min. In cases where technical considerations led to videos of shorter lengths, these videos were removed from analysis. Prior to testing, the camera was centered directly over the simulator using an adjustable tripod and the AKASO GO phone app. The ‘origin’ was denoted by a large black dot over the center perpendicular rod. For origin and direction calibration, short videos were obtained of an Explorer 2.0 Compass being held over the origin.
Model development
We used the convolutional neural network YOLO for efficient processing of videos. This network is implemented in the open-source library Pytorch, which when used with Google Colab and Google Drive, makes for a cost-free environment. We started with two separate pre-trained models, YOLOv5s and YOLOv5m. The former would go on to be used for the detection of the origin, monarch’s head, and body while the latter would be involved in the identification of the compass’s north needle and jewel bearing. We used two different models to account for the different demands of each identification task. The flight videos are typically 10 minutes to 15 minutes long with well-defined objects (such as the monarch’s body and head). Due to this, we could afford to use a smaller architecture such as the v5s to increase processing speeds. Conversely, the compass videos are only five seconds long with more subtle objects (such as the north needle). Since these videos are very short relative to the monarch videos, we were able to use a deeper model to allow for more complexities to be captured with a small tradeoff to processing time.
To increase model robustness, we trained over two rounds. First round training set images were obtained through the random extraction of frames using a random subset of the summer and fall videos. To prevent performance bias for the v5s model, we chose an equal number of flying & non-flying frames (n = 300). For the v5m model, we chose frames with differing placements of the compass needle (n = 74). To annotate our images, we used the open-source software LabelImg (70). Images and labels were flipped horizontally & vertically, contrast & brightness was randomly adjusted, and grayscale was applied. This procedure increased the number of datapoints in our training set by a factor 32, as well as added variation to prevent overfitting. After the first round of training was completed, we analyzed our fall and summer videos. Any problematic frames, for example where the monarch or north needle were not properly identified), were identified, randomly selected, and then combined with the first-round dataset to produce an expanded training set. The final training sets were 19,488 frames and 3,680 frames for the monarch and compass models, respectively. The v5s (monarch) model was trained for 250 epochs with a batch size of 32, while the final v5m (compass) model was trained for 100 epochs with a batch size of 16. Model training performance was assessed using Weights & Biases. The final v5s model had an average prediction success of 96%. (i.e., on average 4% of frames did not receive a monarch or origin prediction).
Flight detection
To identify periods of flight and non-flight, we selectively analyzed the bounding boxes that corresponded to the monarch’s body. Over the course of the video, the area of this box will fluctuate with the activity of the individual (Figure 1A). To be able to automatically identify these periods of activity, we developed the following algorithm.
For a given video, we have an array of boxes (B), an array of areas (A), and an array of seconds (T) all with a length of n. For every ith box (Bi), there is a corresponding area (Ai) and second (Ti). We create an array of slopes (S) that has a length of n – 1 using the following equation:
We then iterate through A and determine if each Ai is a minimum within an i ± 5 interval. To ensure that this “local minimum” corresponds to a period of activity (i.e., it should represent the wing clap of an individual), we check the slopes (Si) that are within the same interval. If there is at least one slope greater than 20000, and another that is less than −20000, we consider this a “true” local minimum. To identify intervals of flight, we iterated through the collection of local minimums (M), where Mi = Ti. The start of an interval was defined as the moment where 0 minimums are found between (Ti – 1, Ti) and at least 5 minimums are found between (Ti, Ti + 1). Conversely, a stop was defined as 5 or more minimums between (Ti – 1, Ti) and 0 minimums between (Ti, Ti + 1).
Only flight intervals lasting greater than 5 s were considered valid flight and kept for analysis. The intervals and parameters selected above were validated empirically by comparing performance of a range of parameters to visual inspection of actual flight intervals as observed in the videos. Wingbeat frequency was calculated by dividing the total number of local minimums by the total flight time of the individual.
Directionality definition and clustering
The standard for determining directional flight has been the Rayleigh Z score cutoff of 500 where Z is given by Z = nr2 where n is the number of points and r is the mean resultant vector. While this threshold method for designating directional flight has produced robust patterns, we note that this method is dependent upon the data sampling frequency. Moreover, the underlying assumption of independent data points required for calculating the Rayleigh Z statistic for individuals is violated (and is more egregiously violated as sampling frequency increases). We, therefore, chose to perform unbiased fuzzy clustering in order to classify individuals’ directionality and flight propensity. We used the R packages ‘ppclust’ (1.1.0) and ‘fclust’ (2.1.1) for these analyses. In the former, we used the function fcm to cluster our data using the Fuzzy C-Means clustering algorithm. This is similar to K-means clustering except that it allows for individuals to be assigned membership scores to all clusters versus being automatically assigned to specific clusters. Algorithm performance was assessed assuming k = 2–7 clusters, and the highest partition coefficient determined the optimal k. Optimal k was 2 in all cases. To prevent bias when choosing the correct number, we chose the one which had the highest partition coefficient.89
Quantification and statistical analysis
All statistics were calculated in R (v.4.1.2) unless otherwise indicated. Rayleigh test of uniformity was used to determine group-oriented directionality. Circular-circular regression was used to determine significance of relationship between flight angle and time of day. Differences in group means of weighted r were assessed among groups using the Kruskal-Wallis rank-sum test. Subsequent pairwise comparisons were made with the Wilcoxon rank-sum test. To investigate the relationship between weighted r and theta for all groups, we fit the data with a generalized additive model using the ‘mgcv’ package. Group distributions were assessed for bimodality using the CircMLE (0.3.0). From this, we used the function circ_mle, which fits the group data to 10 different orientation models and ranks them based on AIC.90 We report the model with the best fit (lowest AIC within a group) as well as models with AIC difference less than or equal to 2. These models are generally considered to be not significantly different from one another.90
Acknowledgments
We are grateful to the contemporary Monarch Butterfly Biosphere Reserve Director (Felipe Martínez Meza), Reserve staff, and community partners for granting access to the overwintering site. We thank M. Isabel Ramírez, Raúl Zubieta, and Osmar O. Ramírez for assistance at the overwintering site. We specifically acknowledge and appreciate the technical assistance provided by Ilia Popov. Members of the Green D.E.I. lab provided helpful discussions during the preparation of this manuscript. Funds from National Geographic Society grant NGS 61319T-19 and NSF-IIBR 2045017 indirectly supported components of this work.
Author contributions
D.A.G. designed the research and secured funding. D.A.G., S.P., and S.M.S. conducted experiments. S.P. wrote the analysis software with input and supervision from D.A.G. S.P., S.M.S., and D.A.G. conducted analyses. D.A.G. and S.P. wrote the initial draft of the paper with subsequent critical input from S.M.S.
Declaration of interests
The authors declare no competing interests.
Published: February 1, 2024
Footnotes
Supplemental information can be found online at https://doi.org/10.1016/j.isci.2024.109063.
Supplemental information
References
- 1.Ramenofsky M., Wingfield J.C. Regulation of Migration. Bioscience. 2007;57:135–143. doi: 10.1641/b570208. [DOI] [Google Scholar]
- 2.Ramenofsky M., Wingfield J.C. Behavioral and physiological conflicts in migrants: the transition between migration and breeding. J. Ornithol. 2006;147:135–145. doi: 10.1007/s10336-005-0050-4. [DOI] [Google Scholar]
- 3.Rankin M.A. Endocrine effects on migration. Am. Zool. 1991;31:217–230. [Google Scholar]
- 4.Sharma A., Tripathi V., Kumar V. Control and adaptability of seasonal changes in behavior and physiology of latitudinal avian migrants: Insights from laboratory studies in Palearctic-Indian migratory buntings. J. Exp. Zool. A Ecol. Integr. Physiol. 2022;337:902–918. doi: 10.1002/jez.2631. [DOI] [PubMed] [Google Scholar]
- 5.Cornelius J.M., Boswell T., Jenni-Eiermann S., Breuner C.W., Ramenofsky M. Contributions of endocrinology to the migration life history of birds. Gen. Comp. Endocrinol. 2013;190:47–60. doi: 10.1016/j.ygcen.2013.03.027. [DOI] [PubMed] [Google Scholar]
- 6.Saino N., Ambrosini R., Albetti B., Caprioli M., De Giorgio B., Gatti E., Liechti F., Parolini M., Romano A., Romano M., et al. Migration phenology and breeding success are predicted by methylation of a photoperiodic gene in the barn swallow. Sci. Rep. 2017;7 doi: 10.1038/srep45412. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Delmore K., Illera J.C., Pérez-Tris J., Segelbacher G., Lugo Ramos J.S., Durieux G., Ishigohoka J., Liedvogel M. The evolutionary history and genomics of European blackcap migration. Elife. 2020;9 doi: 10.7554/elife.54462. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Merlin C., Liedvogel M., Jundi B. el, Kelber A., Webb B. The genetics and epigenetics of animal migration and orientation: birds, butterflies and beyond. J. Exp. Biol. 2019;222:jeb191890. doi: 10.1242/jeb.191890. [DOI] [PubMed] [Google Scholar]
- 9.Chapman J.W., Reynolds D.R., Wilson K. Long-range seasonal migration in insects: mechanisms, evolutionary drivers and ecological consequences. Ecol. Lett. 2015;18:287–302. doi: 10.1111/ele.12407. [DOI] [PubMed] [Google Scholar]
- 10.Ketterson E.D., Nolan V. Bird Migration. Physiology and Ecophysiology. 1990:117–129. doi: 10.1007/978-3-642-74542-3_8. [DOI] [Google Scholar]
- 11.Ramenofsky M., Wingfield J.C. Regulation of complex behavioural transitions: migration to breeding. Anim. Behav. 2017;124:299–306. doi: 10.1016/j.anbehav.2016.09.015. [DOI] [Google Scholar]
- 12.Jiang X., Luo L., Zhang L., Sappington T.W., Hu Y. Regulation of Migration in Mythimna separata (Walker) in China: A Review Integrating Environmental, Physiological, Hormonal, Genetic, and Molecular Factors. Environ. Entomol. 2011;40:516–533. doi: 10.1603/en10199. [DOI] [PubMed] [Google Scholar]
- 13.Lohmann K.J., Putman N.F., Lohmann C.M.F. Geomagnetic imprinting: A unifying hypothesis of long-distance natal homing in salmon and sea turtles. Proc. Natl. Acad. Sci. USA. 2008;105:19096–19101. doi: 10.1073/pnas.0801859105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Putman N.F., Lohmann K.J., Putman E.M., Quinn T.P., Klimley A.P., Noakes D.L.G. Evidence for Geomagnetic Imprinting as a Homing Mechanism in Pacific Salmon. Curr. Biol. 2013;23:312–316. doi: 10.1016/j.cub.2012.12.041. [DOI] [PubMed] [Google Scholar]
- 15.Brothers J.R., Lohmann K.J. Evidence for Geomagnetic Imprinting and Magnetic Navigation in the Natal Homing of Sea Turtles. Curr. Biol. 2015;25:392–396. doi: 10.1016/j.cub.2014.12.035. [DOI] [PubMed] [Google Scholar]
- 16.Scholz A.T., Horrall R.M., Cooper J.C., Hasler A.D. Imprinting to Chemical Cues: The Basis for Home Stream Selection in Salmon. Science. 1976;192:1247–1249. doi: 10.1126/science.1273590. [DOI] [PubMed] [Google Scholar]
- 17.Bett N.N., Hinch S.G., Dittman A.H., Yun S.-S. Evidence of Olfactory Imprinting at an Early Life Stage in Pink Salmon (Oncorhynchus gorbuscha) Sci. Rep. 2016;6 doi: 10.1038/srep36393. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Robart A.R., Zuñiga H.X., Navarro G., Watts H.E. Social environment influences termination of nomadic migration. Biol. Lett. 2022;18 doi: 10.1098/rsbl.2022.0006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Winger B.M., Auteri G.G., Pegan T.M., Weeks B.C. A long winter for the Red Queen: rethinking the evolution of seasonal migration. Biol. Rev. 2019;94:737–752. doi: 10.1111/brv.12476. [DOI] [PubMed] [Google Scholar]
- 20.Schmaljohann H., Eikenaar C., Sapir N. Understanding the ecological and evolutionary function of stopover in migrating birds. Biol. Rev. 2022;97:1231–1252. doi: 10.1111/brv.12839. [DOI] [PubMed] [Google Scholar]
- 21.Van Huis A., Cressman K., Magor J.I. Preventing desert locust plagues: optimizing management interventions. Entomol. Exp. Appl. 2007;122:191–214. doi: 10.1111/j.1570-7458.2006.00517.x. [DOI] [Google Scholar]
- 22.Vidal O., Rendón-Salinas E. Dynamics and trends of overwintering colonies of the monarch butterfly in Mexico. Biol. Conserv. 2014;180:165–175. doi: 10.1016/j.biocon.2014.09.041. [DOI] [Google Scholar]
- 23.Urquhart F.A., Urquhart N.R. Autumnal migration routes of the eastern population of the monarch butterfly (Danaus p. plexippus L.; Danaidae; Lepidoptera) in North America to the overwintering site in the Neovolcanic Plateau of Mexico. Can. J. Zool. 1978;56:1759–1764. doi: 10.1139/z78-240. [DOI] [Google Scholar]
- 24.Calvert W.H., Zuchowski W., Brower L.P. The impact of forest thinning on microclimate in monarch butterfly ( Danaus pexippus L.) overwintering areas of Mexico. Bot. Sci. 1982;11:11–18. doi: 10.17129/botsci.1257. [DOI] [Google Scholar]
- 25.Leong K.L.H., Frey D., Brenner G., Baker S., Fox D. Use of Multivariate Analyses to Characterize the Monarch Butterfly (Lepidoptera: Danaidae) Winter Habitat. Ann. Entomol. Soc. Am. 1991;84:263–267. doi: 10.1093/aesa/84.3.263. [DOI] [Google Scholar]
- 26.Guerra P.A. The Monarch Butterfly as a Model for Understanding the Role of Environmental Sensory Cues in Long-Distance Migratory Phenomena. Front. Behav. Neurosci. 2020;14 doi: 10.3389/fnbeh.2020.600737. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Herman W.S., Lessman C.A., Johnson G.D. Correlation of juvenile hormone titer changes with reproductive tract development in the posteclosion Monarch butterfly. J. Exp. Zool. 1981;218:387–395. doi: 10.1002/jez.1402180310. [DOI] [Google Scholar]
- 28.Green D.A., Kronforst M.R. Monarch butterflies use an environmentally sensitive, internal timer to control overwintering dynamics. Mol. Ecol. 2019;28:3642–3655. doi: 10.1111/mec.15178. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Perez S.M., Taylor O.R., Jander R. A sun compass in monarch butterflies. Nature. 1997;387:29. doi: 10.1371/journal.pbio.0060004. [DOI] [Google Scholar]
- 30.Mouritsen H., Frost B.J. Virtual migration in tethered flying monarch butterflies reveals their orientation mechanisms. Proc. Natl. Acad. Sci. USA. 2002;99:10162–10166. doi: 10.1073/pnas.152137299. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Guerra P.A., Gegear R.J., Reppert S.M. A magnetic compass aids monarch butterfly migration. Nat. Commun. 2014;5:4164. doi: 10.1038/ncomms5164. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Froy O., Gotter A.L., Casselman A.L., Reppert S.M. Illuminating the Circadian Clock in Monarch Butterfly Migration. Science. 2003;300:1303–1305. doi: 10.1126/science.1084874. [DOI] [PubMed] [Google Scholar]
- 33.Sauman I., Briscoe A.D., Zhu H., Shi D., Froy O., Stalleicken J., Yuan Q., Casselman A., Reppert S.M. Connecting the Navigational Clock to Sun Compass Input in Monarch Butterfly Brain. Neuron. 2005;46:457–467. doi: 10.1016/j.neuron.2005.03.014. [DOI] [PubMed] [Google Scholar]
- 34.Merlin C., Gegear R.J., Reppert S.M. Antennal Circadian Clocks Coordinate Sun Compass Orientation in Migratory Monarch Butterflies. Science. 2009;325:1700–1704. doi: 10.1126/science.1176221. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Guerra P.A., Merlin C., Gegear R.J., Reppert S.M. Discordant timing between antennae disrupts sun compass orientation in migratory monarch butterflies. Nat. Commun. 2012;3:958–967. doi: 10.1038/ncomms1965. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Guerra P.A., Reppert S.M. Coldness Triggers Northward Flight in Remigrant Monarch Butterflies. Curr. Biol. 2013;23:419–423. doi: 10.1016/j.cub.2013.01.052. [DOI] [PubMed] [Google Scholar]
- 37.Parlin A.F., Stratton S.M., Guerra P.A. Assaying lepidopteran flight directionality with non-invasive methods that permit repeated use and release after testing. Methods Ecol. Evol. 2021;12:1699–1704. doi: 10.1111/2041-210x.13648. [DOI] [Google Scholar]
- 38.Zhu H., Gegear R.J., Casselman A., Kanginakudru S., Reppert S.M. Defining behavioral and molecular differences between summer and migratory monarch butterflies. BMC Biol. 2009;7:14. doi: 10.1186/1741-7007-7-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Moore B.R. A modification of the Rayleigh test for vector data. Biometrika. 1980;67:175–180. doi: 10.1093/biomet/67.1.175. [DOI] [Google Scholar]
- 40.Zhang L., Cheng L., Chapman J.W., Sappington T.W., Liu J., Cheng Y., Jiang X. Juvenile hormone regulates the shift from migrants to residents in adult oriental armyworm, Mythimna separata. Sci. Rep. 2020;10 doi: 10.1038/s41598-020-66973-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Coats S.A., Tollefson J.J., Mutchmor J.A. Study of Migratory Flight in the Western Corn Rootworm (Coleoptera: Chrysomelidae) Environ. Entomol. 1986;15:620–625. doi: 10.1093/ee/15.3.620. [DOI] [Google Scholar]
- 42.Talla V., Pierce A.A., Adams K.L., de Man T.J.B., Nallu S., Villablanca F.X., Kronforst M.R., de Roode J.C. Genomic evidence for gene flow between monarchs with divergent migratory phenotypes and flight performance. Mol. Ecol. 2020;29:2567–2582. doi: 10.1111/mec.15508. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Davis A.K., Chi J., Bradley C., Altizer S. The Redder the Better: Wing Color Predicts Flight Performance in Monarch Butterflies. PLoS One. 2012;7 doi: 10.1371/journal.pone.0041323.s003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Coats S.A.O. Iowa State University; 1986. Migratory Flight Potential and the Role of Juvenile Hormone in Flight Regulation in the Western Corn Rootworm. [DOI] [Google Scholar]
- 45.Naranjo S.E. Assessing Insect Flight Behavior in the Laboratory: A Primer on Flight Mill Methodology and What Can Be Learned. Ann. Entomol. Soc. Am. 2019;112:182–199. doi: 10.1093/aesa/say041. [DOI] [Google Scholar]
- 46.Shlizerman E., Phillips-Portillo J., Forger D.B., Reppert S.M. Neural Integration Underlying a Time-Compensated Sun Compass in the Migratory Monarch Butterfly. Cell Rep. 2016;15:683–691. doi: 10.1016/j.celrep.2016.03.057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Beetz M.J., Kraus C., El Jundi B. Neural representation of goal direction in the monarch butterfly brain. Nat. Commun. 2023;14:5859. doi: 10.1038/s41467-023-41526-w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Calvert W.H. Monarch Butterfly (Danaus plexippus L. Nymphalidae) Fall Migration: Flight Behavior and Direction in Relation to Celestial and Physiographic Cues. J. Lepid. Soc. 2001;55:162–168. [Google Scholar]
- 49.Calvert W.H., Brower L.P. The Location of Monarch Butterfly (Danaus plexippus L.) Overwintering Colonies in Mexico in Relation to Topography and Climate. J. Lepidopterists' Soc. 1986;40:164–187. [Google Scholar]
- 50.Sánchez-Tlacuahuac N., Pimentel-Equihua J.L., Espinosa-Hernández V., Vibrans H. What do monarchs feed on in winter? Nectar sources at hibernation sites. J. Insect Conserv. 2023;27:181–191. doi: 10.1007/s10841-022-00433-z. [DOI] [Google Scholar]
- 51.Brower L.P., Calvert W.H., Hedrick L.E., Christian J. Biological Observations on an Overwintering Colony of Monarch Butterflies (Danaus plexippus, Danaidae) in Mexico. J. Lepid. Soc. 1977;31:232–242. [Google Scholar]
- 52.Barras A.G., Liechti F., Arlettaz R. Seasonal and daily movement patterns of an alpine passerine suggest high flexibility in relation to environmental conditions. J. Avian Biol. 2021;52 doi: 10.1111/jav.02860. [DOI] [Google Scholar]
- 53.Terrill S.B. Evolutionary aspects of orientation and migration in birds. EXS. 1991;60:180–201. doi: 10.1007/978-3-0348-7208-9_9. [DOI] [PubMed] [Google Scholar]
- 54.Beetz M.J., Kraus C., Franzke M., Dreyer D., Strube-Bloss M.F., Rössler W., Warrant E.J., Merlin C., El Jundi B. Flight-induced compass representation in the monarch butterfly heading network. Curr. Biol. 2022;32:338–349.e5. doi: 10.1016/j.cub.2021.11.009. [DOI] [PubMed] [Google Scholar]
- 55.Heinze S., Florman J., Asokaraj S., El Jundi B., Reppert S.M. Anatomical basis of sun compass navigation II: The neuronal composition of the central complex of the monarch butterfly. J. Comp. Neurol. 2013;521:267–298. doi: 10.1002/cne.23214. [DOI] [PubMed] [Google Scholar]
- 56.Schroeder H., Majewska A., Altizer S. Monarch butterflies reared under autumn-like conditions have more efficient flight and lower post-flight metabolism. Ecol. Entomol. 2020;45:562–572. doi: 10.1111/een.12828. [DOI] [Google Scholar]
- 57.Zhan S., Zhang W., Niitepõld K., Hsu J., Haeger J.F., Zalucki M.P., Altizer S., de Roode J.C., Reppert S.M., Kronforst M.R. The genetics of monarch butterfly migration and warning colouration. Nature. 2014;514:317–321. doi: 10.1038/nature13812. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Tenger-Trolander A., Julick C.R., Lu W., Green D.A., Montooth K.L., Kronforst M.R. Seasonal plasticity in morphology and metabolism differs between migratory North American and resident Costa Rican monarch butterflies. Ecol. Evol. 2023;13 doi: 10.1002/ece3.9796. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Terrill S.B. Bird Migration. Physiology and Ecophysiology. 1990:130–143. doi: 10.1007/978-3-642-74542-3_9. [DOI] [Google Scholar]
- 60.Brönmark C., Hulthén K., Nilsson P.A., Skov C., Hansson L.A., Brodersen J., Chapman B.B. There and back again: migration in freshwater fishes 1. Can. J. Zool. 2014;92:467–479. doi: 10.1139/cjz-2012-0277. [DOI] [Google Scholar]
- 61.Xu W., Barker K., Shawler A., Van Scoyoc A., Smith J.A., Mueller T., Sawyer H., Andreozzi C., Bidder O.R., Karandikar H., et al. The plasticity of ungulate migration in a changing world. Ecology. 2021;102 doi: 10.1002/ecy.3293. [DOI] [PubMed] [Google Scholar]
- 62.Warrant E., Frost B., Green K., Mouritsen H., Dreyer D., Adden A., Brauburger K., Heinze S. The Australian Bogong Moth Agrotis infusa: A Long-Distance Nocturnal Navigator. Front. Behav. Neurosci. 2016;10:77. doi: 10.3389/fnbeh.2016.00077. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Eggeman S.L., Hebblewhite M., Bohm H., Whittington J., Merrill E.H. Behavioural flexibility in migratory behaviour in a long-lived large herbivore. J. Anim. Ecol. 2016;85:785–797. doi: 10.1111/1365-2656.12495. [DOI] [PubMed] [Google Scholar]
- 64.Mysterud A., Loe L.E., Zimmermann B., Bischof R., Veiberg V., Meisingset E. Partial migration in expanding red deer populations at northern latitudes – a role for density dependence? Oikos. 2011;120:1817–1825. doi: 10.1111/j.1600-0706.2011.19439.x. [DOI] [Google Scholar]
- 65.Sorel M.H., Murdoch A.R., Zabel R.W., Kamphaus C.M., Buhle E.R., Scheuerell M.D., Converse S.J. Effects of population density and environmental conditions on life-history prevalence in a migratory fish. Ecol. Evol. 2023;13 doi: 10.1002/ece3.10087. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Chapman B.B., Brönmark C., Nilsson J.-Å., Hansson L.-A. The ecology and evolution of partial migration. Oikos. 2011;120:1764–1775. doi: 10.1111/j.1600-0706.2011.20131.x. [DOI] [Google Scholar]
- 67.Menz M.H.M., Reynolds D.R., Gao B., Hu G., Chapman J.W., Wotton K.R. Mechanisms and Consequences of Partial Migration in Insects. Front. Ecol. Evol. 2019;7:403. doi: 10.3389/fevo.2019.00403. [DOI] [Google Scholar]
- 68.Bond M.H., Miller J.A., Quinn T.P. Beyond dichotomous life histories in partially migrating populations: cessation of anadromy in a long-lived fish. Ecology. 2015;96:1899–1910. doi: 10.1890/14-1551.1. [DOI] [PubMed] [Google Scholar]
- 69.Skov C., Baktoft H., Brodersen J., Brönmark C., Chapman B.B., Hansson L.-A., Nilsson P.A. Sizing up your enemy: individual predation vulnerability predicts migratory probability. Proc. Biol. Sci. 2011;278:1414–1418. doi: 10.1098/rspb.2010.2035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Cheng Y. 2022. Migration and Survival in a Changing World: Insights on a Long-Distance Avian Migrant. [Google Scholar]
- 71.Mallon J.M., Bildstein K.L., Fagan W.F. Inclement weather forces stopovers and prevents migratory progress for obligate soaring migrants. Mov. Ecol. 2021;9:39. doi: 10.1186/s40462-021-00274-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Holberton R.L. An Endogenous Basis for Differential Migration in the Dark-Eyed Junco. Condor. 1993;95:580–587. doi: 10.2307/1369602. [DOI] [Google Scholar]
- 73.Northrup J.M., Vander Wal E., Bonar M., Fieberg J., Laforge M.P., Leclerc M., Prokopenko C.M., Gerber B.D. Conceptual and methodological advances in habitat-selection modeling: guidelines for ecology and evolution. Ecol. Appl. 2022;32 doi: 10.1002/eap.2470. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Winkler D.W., Jørgensen C., Both C., Houston A.I., McNamara J.M., Levey D.J., Partecke J., Fudickar A., Kacelnik A., Roshier D., Piersma T. Cues, strategies, and outcomes: how migrating vertebrates track environmental change. Mov. Ecol. 2014;2:10. doi: 10.1186/2051-3933-2-10. [DOI] [Google Scholar]
- 75.Goymann W., Lupi S., Kaiya H., Cardinale M., Fusani L. Ghrelin affects stopover decisions and food intake in a long-distance migrant. Proc. Natl. Acad. Sci. USA. 2017;114:1946–1951. doi: 10.1073/pnas.1619565114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Pant K., Chandola-Saklani A. A role for thyroid hormones in the development of premigratory disposition in redheaded bunting, Emberiza bruniceps. J. Comp. Physiol. B. 1993;163:389–394. doi: 10.1007/bf00265643. [DOI] [Google Scholar]
- 77.Fudickar A.M., Greives T.J., Abolins-Abols M., Atwell J.W., Meddle S.L., Friis G., Stricker C.A., Ketterson E.D. Mechanisms Associated with an Advance in the Timing of Seasonal Reproduction in an Urban Songbird. Front. Ecol. Evol. 2017;5:85. doi: 10.3389/fevo.2017.00085. [DOI] [Google Scholar]
- 78.Sudo R., Suetake H., Suzuki Y., Utoh T., Tanaka S., Aoyama J., Tsukamoto K. Dynamics of Reproductive Hormones During Downstream Migration in Females of the Japanese Eel, Anguilla japonica. Zool. Sci. 2011;28:180–188. doi: 10.2108/zsj.28.180. [DOI] [PubMed] [Google Scholar]
- 79.McCormick S.D. Evolution of the hormonal control of animal performance: Insights from the seaward migration of salmon. Integr. Comp. Biol. 2009;49:408–422. doi: 10.1093/icb/icp044. [DOI] [PubMed] [Google Scholar]
- 80.Wibbels T., Owens D.W., Limpus C.J., Reed P.C., Amoss M.S. Seasonal changes in serum gonadal steroids associated with migration, mating, and nesting in the loggerhead sea turtle (Caretta caretta) Gen. Comp. Endocrinol. 1990;79:154–164. doi: 10.1016/0016-6480(90)90099-8. [DOI] [PubMed] [Google Scholar]
- 81.Zhang L., Luo L., Jiang X. Starvation influences allatotropin gene expression and juvenile hormone titer in the female adult oriental armyworm, Mythimna separata. Arch Insect Biochem. 2008;68:63–70. doi: 10.1002/arch.20255. [DOI] [PubMed] [Google Scholar]
- 82.Rankin S.M., Rankin M.A. The hormonal control of migratory flight behaviour in the convergent ladybird beetle, Hippodamia convergens. Physiol. Entomol. 1980;5:175–182. doi: 10.1111/j.1365-3032.1980.tb00224.x. [DOI] [Google Scholar]
- 83.Lessman C.A., Herman W.S. Flight enhances juvenile hormone inactivation inDanaus plexippus plexippus L. (Lepidoptera: Danaidae) Experientia. 1981;37:599–601. doi: 10.1007/bf01990072. [DOI] [Google Scholar]
- 84.Mouritsen H. Long-distance navigation and magnetoreception in migratory animals. Nature. 2018;558:50–59. doi: 10.1038/s41586-018-0176-1. [DOI] [PubMed] [Google Scholar]
- 85.Green D.A. Monarch Butterfly Migration as an Integrative Model of Complex Trait Evolution. Am. Nat. 2021;198:142–157. doi: 10.1086/714526. [DOI] [PubMed] [Google Scholar]
- 86.Pérez-Miranda R., Arriola-Padilla V.J., Romero-Sanchez M.E. Characterizing New Wintering Sites for Monarch Butterfly Colonies in Sierra Nevada, Mexico. Insects. 2020;11:384. doi: 10.3390/insects11060384. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Oberhauser K., Peterson A.T. Modeling current and future potential wintering distributions of eastern North American monarch butterflies. Proc. Natl. Acad. Sci. USA. 2003;100:14063–14068. doi: 10.1073/pnas.2331584100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Sáenz-Romero C., Rehfeldt G.E., Duval P., Lindig-Cisneros R.A. Abies religiosa habitat prediction in climatic change scenarios and implications for monarch butterfly conservation in Mexico. For. Ecol. Manage. 2012;275:98–106. doi: 10.1016/j.foreco.2012.03.004. [DOI] [Google Scholar]
- 89.Ferraro M.B., Giordani P. A toolbox for fuzzy clustering using the R programming language. Fuzzy Set Syst. 2015;279:1–16. doi: 10.1016/j.fss.2015.05.001. [DOI] [Google Scholar]
- 90.Schnute J.T., Groot K. Statistical analysis of animal orientation data. Anim. Behav. 1992;43:15–33. doi: 10.1016/s0003-3472(05)80068-5. [DOI] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.