

Abstract
The biomarker cortisol assesses the impact of biopsychosocial stressors that activate the stress response system. Hair has emerged as a valid and non-invasive means of gauging cumulative cortisol deposited over month-long periods of time. Established protocols for the extraction of hair cortisol are being validated and refined in humans, yet methodological information about hair characteristics on cortisol remains limited. In addition to external hair exposures (e.g. dye, time spent outside), we examined hair categorization or type (e.g. kinky, straight) by extending a hair typing methodology for scientific use that is currently popular among hair care professionals. We then examined the interaction between hair type and race on cortisol levels with a hair questionnaire. Three studies were pooled to investigate how sample weight, hair type, race, heat exposures, and hair treatments impacted cumulative hair cortisol concentrations. Study 1 consisted of Adult Kenyan Medical Workers (N=44); Study 2 Mexican and Mexican Americans (N=106); and Study 3 American Youth (N=107). We found significantly higher cortisol in 5mg of hair when compared to larger sample weights, and higher cortisol in those who spent more time outdoors. Cortisol concentrations differed between racial groups and varied by hair type; moreover, there were directional differences in cumulative cortisol from straighter to curlier hair types which depended on racial group. In addition to demonstrating the impact of relatively novel control factors like hair sample weight, outdoor exposure, and hair type, the present study illustrates the importance of disentangling hair type and race to understand variability in cumulative hair cortisol. These influences should be included in future studies that measure hair cortisol.
Keywords: Hair Cortisol, Hair Hormone Methodology, Race, Hair Typing System
1. Introduction:
Hair has been used to measure hormones consistently since the mid-70’s (Ebling, 1976; Schweikert & Wilson, 1974) to gauge hormone levels in wild animal populations (Azevedo et al., 2019) and non-human primates in laboratory settings (Dettmer et al., 2014). Despite the ease of obtaining hair, it was not until the late-90’s to early-2000’s that validation of hormone stability in the hair of humans (Yang et al., 1998); (Koren et al., 2002) and non-human primates took root (Davenport et al., 2006). Hair has grown into an established biospecimen that can be collected non-invasively to measure monthly cumulative cortisol concentrations which complements snapshots of hormone release associated with the stress response system (Sharpley et al., 2011).
Cortisol is the hormonal end-product of the hypothalamic-pituitary-adrenal axis which is activated to cope with stressful stimuli (McEwen, 1998). After encountering physical stressors, such as pain (Zimmer et al., 2003) or psychosocial stressors, such as experiences of racial discrimination (Adam et al., 2015; Palmer-Bacon et al., 2020), the stress response system triggers limbic neurocircuitry activation. Alongside emotion-related structures, the hypothalamus is prompted to initiate a hormonal cascade (Tsigos & Chrousos, 1994) that culminates in the production and secretion of glucocorticoids from the adrenal glands (Munck et al., 1984), including cortisol. Cortisol is an established lipid-soluble biomarker for physiological and emotional stress, measurable in a variety of biospecimens (Meyer & Novak, 2012).
1.1. Why is the measurement of cortisol in hair important and practical?
There are advantages to measuring cortisol with hair compared to traditional biofluids like blood, plasma or saliva. Collection is straightforward and can be implemented in multiple point-of-collection settings (e.g., lab, field, and home). Hair provides a cumulative measure of cortisol (Koren et al., 2002) that indexes chronic stress exposure across developmental stages (Bates et al., 2017). Hair captures hormone levels over approximate month-long timeframes in a single sample (Meyer & Novak, 2012), which minimizes cost for obtaining basal concentrations without sacrificing reliability (E. Russell et al., 2012). Unlike biofluids like saliva and blood, hair does not capture cortisol release that occurs after the immediate exposure to stressors like skydiving (White et al., 2019), social evaluative threat (Kudielka et al., 2004), and exercise (Hayes et al., 2015) or as part of circadian rhythmicity or momentary daily stress changes (Engert et al., 2018). Hair cortisol essentially averages across experiences of stress to index chronic release over 1 to 4 month intervals.
Noninvasive sampling and ease of collection is important when researching vulnerable human populations such as children (Bates et al., 2017), pregnant women (Marceau et al. 2020), or individuals with needle phobia. Saliva as a biospecimen solved many of the limitations of venipuncture and phlebotomy by eliminating pain and discomfort of repeated blood draws in, for example, children (Taddio et al. 2009; Orenius et al. 2018), pregnant women (McLenon and Rogers 2019), and the general population (Love and Love 2021). Over time and with repeated narrowly-timed sampling, saliva collection becomes inconvenient which contributes to missing data and systematic error due to lack of adherence to collection protocol (Fernandes et al. 2013). Individuals with inflexible schedules (e.g. multiple jobs, student workers, and busy caregivers) may prefer the ease of collecting a single biospecimen sample, thereby reducing attrition and bias. Likely reflecting the inconvenience of collecting multiple tightly timed saliva samples, individuals in vulnerable lower socioeconomic status (SES) populations showed less compliance to collection adherence than those of higher SES (Hill Golden et al., 2014). Such sources of error are avoidable with hair where it is unnecessary to record and validate precise collection time points (Hall et al., 2011; Skinner et al., 2011).
Hair remains stable over long periods of time at room temperature (Gow et al., 2010; E. Russell et al., 2012) and cortisol deposited in hair remains detectable over millennia (Webb et al., 2015), which reduces concern about sample degradation. Hair does not call for refrigeration or freezing immediately after collection or during transportation, allowing samples to be shipped from rural or economically disadvantaged regions that may not have the infrastructure to conduct assays nearby (Greff et al., 2019; E. Russell et al., 2012). Room temperature stability also eliminates the need for storage in ultracold freezers which may not be available (e.g. during work hours, remote locations, participant homes) or feasible during transportation. Hair allows researchers to eliminate financial fees of overnight shipping, heavy packaging, and hazard costs from shipping with dry ice. Finally, in-home collection is feasible because it is easy to obtain hair collection tools (e.g. scissors, rubber bands). This is imperative when in-lab collection is impracticable, as with the recent COVID-19 pandemic.
1.2. Methodological challenges in collecting hair.
Hair collection methodologies are not without challenges. Insufficient sample weights can indicate poor sample quality. This may occur during self-collection, with inexperienced research assistants uncomfortable collecting larger sample volumes when samples are allotted for multiple hormone extractions, or when shorter segments of hair are required to meet ideal sample weight (Meyer & Novak, 2012). Insufficient sample weights can result in systemic error as hair hormones from distal segments do not correlate as strongly with proximal scalp-region segments (Wang, van Dammen, Moody, Kiesner, et al., 2020; Wang, van Dammen, Moody, Neiderhiser, et al., 2020). Similarly, in chimpanzees, lower hair weights resulted in greater cortisol concentrations (Yamanashi et al., 2016). Sample weight differences within studies can exert deleterious systematic error. Inadequate sample weights as a source of error can be minimized with clear instructions and ample training for participants or researchers to collect “tiny snips” of hair taken from various parts of the scalp (SpitLab H.A.I.R. Project, n.d.). Collection of multiple smaller samples may also alleviate discomfort from the perception of hair loss or “noticeable divots”.
Bias can be introduced from superficial variables impacting hair, including exposure to heat, time spent outside, and chemical treatment (e.g. dye, perms, relaxers). Hair exposed to natural sunlight and UV radiation exposure reduces concentrations of glucocorticoids like cortisol (Wester et al., 2016). Heat-based treatments are linked with lower cortisol in a community sample (Fischer et al., 2017). Regarding hair dye and other chemical treatments, studies have found lower cortisol in dyed hair versus undyed hair (Sauvé et al., 2007) and in dyed and/or chemically treated hair (Gidlow et al., 2016). External hair exposures are not yet routinely reported as control measures in studies with hair as a biospecimen, leaving ambiguity about the interpretation of cortisol concentrations.
Given that hair varies in absorption of lipids (Martí et al., 2016), and hormones like cortisol are lipid-soluble (Kudielka et al., 2012), it is important to establish if lipid-soluble hormone concentrations differ by hair type. Studies that explore hair absorption are largely limited to forensic research in illicit drug use (Boumba et al., 2006; Joseph et al., 1996; Mieczkowski, 1999). This research shows greater lipid absorption with “African” hair (Coderch et al., 2019; Cruz et al., 2013), though it was unclear if this was due to hair thickness and or shape – both of which influence lipid absorption. Little research tests the impact of hair type on hormones in psychosocial sciences (Koch et al., 2020). This may be due to the novelty of hair hormones, lack of variability in hair types within studies, and dearth of widely adopted methodology for categorizing hair types in biosocial fields. Developing a systematic characterization of participant hair type across the full range of human hair types addresses these limitations.
1.3. Historic hair categorization methods require a critical update
The knowledge gap about hair type illustrates a dire need for updated methods to categorize human hair. To eliminate inaccurate methodologies, this update is critical for historic, social/pragmatic, and biological reasons. One early hair typing system was developed by Eugen Fischer as a means to classify hair by color and texture to determine “greater” or “lesser racial value” when compared to the blonde-haired Nordic “ideal” (Evans, 2008; UCL, 2016). Fischer’s typing system was developed during German occupation of what is now Namibia and coincides with the genocide of the Herero and Nama people who resisted German occupation of their land between 1904 and 1908 (Africa, 1908; Correa, 2011; Melber, 2017; Onishi & Eddy, 2021). Methods to categorize hair were likewise developed by biological and anthropological scientists based on historical categorization of humans into “racial subspecies” (e.g. negroid, mongoloid, caucasoid hair) which conflated hair type and racial groups (Coon, 1963; Jackson, 2001). These terms originate from a historically limited and racist view of categorizing varying “species” of humans (Cell Editorial Team, 2020; A. Fuentes, 2021; Sheth, 2019), perpetuating a well-disputed biological basis of racial classification using debunked science like craniology (Bhopal, 2007).
Hair classification that relies on archaic language and pseudoscience to describe and “classify” race (Eddy, 1938; Matiegka & Malý, 1929) is problematic because it does not acknowledge that race is a social construct (C. Russell, 2021). Pragmatically, using language such as mongoloid or negroid to describe hair type is objectionable to participants and researchers, risking widespread missing data. This pattern is well documented across scientific history that perpetuates discriminatory methodological practices (M. Randall et al., 2021). Narrowly categorizing hair as “negroid, mongoloid, and caucasoid” is also biologically dubious. In using these terms “type” become synonymous with race and the variability of hair types within racial groups becomes impossible to disentangle from race. These categorizations also do not take into account differences in hormone absorption within hair types because they do not recognize the variability in thickness and shape within or between racial groups across kinky, curly, and straight hair textures (Koch et al., 2020).
Seemingly, more scientific approaches for hair typing emerged which relied on the shape of hair (e.g. number of twists per centimeter) (De la Mettrie et al., 2007) and variability of curls within groups (Loussouarn et al., 2007). These methods, unfortunately, are largely impractical. Categorization based on absorption is not easily inferred outside of specialized fields and are still often based in historically racist ideology such as formulating research questions only by racial groups, not allowing variability in hair within racial groups, and classifying hair as “African” and “Caucasian” (Coderch et al., 2019; Martí et al., 2016). The property of hair changes as its structure becomes curlier and some inherent nesting of hair types within racial groups persists (Loussouarn et al., 2016). Since cortisol is a lipid (Pardridge & Mietus, 1979) and higher lipid concentration are found in “African” hair (Coderch et al., 2019), within racial group hair considerations are pertinent to understanding how cortisol differs by hair types beyond race.
Experiences of stress are tied to systemic laws and policies that create social, economic, and health inequality in Black and minoritized communities due to racial discrimination and systemic structural racism (Bailey et al., 2021). Complex social structures culminate in structural racism whereby policies and laws perpetuate economic and social disadvantages based on color and/or ethnic origin (Mueller, 2013). Higher hair cortisol is related to discrimination within Black (Lehrer et al., 2020) as well as Latino/Hispanic, Asian, and White samples (Yip et al., 2021). Structural racism and discrimination also leads to within racial group differences in chronic stress exposure and cortisol due to income (Samuel et al., 2018; Cohen et al., 2006), educational levels (Ursache et al., 2017), discrimination (Yip et al., 2021; Palmer-Bacon et al., 2020), and the intersectionality of gender (Kudielka et al., 2004). However, race remains a persistent predictor of cortisol even after controlling for education, gender, and income (Cohen et al., 2006; Hajat et al., 2010). Testing methodological variables like hair type will better probe within-group racial differences in racism and discrimination exposure, and more broadly psychosocial and biological stressors.
This study introduces methodology for hair typing based on established categories used by haircare professionals that remains novel within scientific settings (Kymberlee et al., 2018). Developed to tailor hair products for specific hair density, curl, and thickness, the stylist Andre Walker (Walker et al., 1997) classifies hair type categorically from 1 (straight) to 4 (tightly coiled); the addition of subcategories a, b, and c for hair type carry unique meaning, yet remain useful to classify hair thickness or pattern within each hair type. Type 4 hair represents kinky hair with type-”a” indicating a tightly coiled but fine curl and type-”b” representing a Z-angled coil that is frizzier compared to type-”a”. Walker (1997) describes type 3 hair as curly, with type-“a” indicating a loose curl and type-”b” representing a corkscrew curl. Type 2, classified as wavy hair, ranges from type-”a” fine and thin hair, to type-”c” coarse and frizzy hair. Type 1 represents straight hair that is resistant to curling, ranging from type-”a” indicating fine & fragile hair to type-”c” indicating coarse & thin hair. It is important to note that Walker’s original scale did not include “c” categories for hair types 3 and 4 hair. Walker’s method for typing hair eliminates outdated and objectionable methods, while capturing variability within hair shape patterns. Especially for those already familiar with the system, hair typing using haircare professionals’ system is easily inferred (Kymberlee et al., 2018).
1.4. Hypothesis:
We examine how hair type, race, and their interaction influences absorption of hair cortisol. We hypothesize that hair types will show differences in hair cortisol, with curlier hair types showing a higher concentration of cortisol (H1). We expect that Black participants will have higher cortisol than Multiracial (Mixed)1, LatinX2 and White participants (H2), and higher concentrations in curlier hair types persisting beyond racial group identification (H3). We predict that hair samples which were exposed to heat (H4.1), greater time spent outside (H4.2), greater hair dryer usage (H4.3), was dyed (H4.4), and permed or chemically treated (H4.5) will have lower concentrations of hormones. A secondary aim evaluates whether weight, a measure of sample collection quality, influences concentrations of hair hormones, such that smaller samples weights produce higher concentrations of hair cortisol (H5). Hypotheses are tested with three studies to look at a diverse sample set of hair types.
2. Methods:
2.1. Sample collection and Population Descriptions:
2.1.1. Kenyan Population (Study 1): Field Collection
Hair was collected from workers in maternity units in a rural county in Western Kenya (N = 44), consisting of 43 females and one male, with a mean age of 33.98 years (range = 24–50y). All participants worked in a hospital setting and included nurses, midwives, doctors, clinical officers and support staff. All participants were Black Africans. Hair was securely banded close to the scalp before being cut and secured with tape on the non-scalp end in secure breathable packaging. Hair was collected from the parietal region perpendicular to the center of the ear. All samples were collected by field researchers and took place between July 1st and August 20th, 2019 during Kenya’s “Winter” season. Samples were shipped to the United States for processing.
2.1.2. Mexican Immigrant Population (Study 2): Field Collection
Study 2 consisted of 57 first generation adults and 51 first or second generation children (N = 108) Mexican Americans. Adults were 73 percent female, with a mean age of 40.27 years (Range = 27–65y). Children were 69 percent female, with a mean age of 10.08 years (Range = 5–18y). All parents were born in Mexico and resided in a Midwestern state. Participants took part in a community-based healthy lifestyles intervention aimed at preventing obesity among low-income Latino/Hispanic families (Sanchez et al., 2019; Teran-Garcia et al., 2019). All participants identified as Mexican or Mexican-American. Hair samples were collected by trained research assistants at community hospitals or community centers. Alongside other biospecimen collections (e.g. blood draws), hair samples were collected by sectioning off small sections using the “tiny snips” method (SpitLab H.A.I.R. Project, n.d.) before banding and cutting hair close to the scalp. Samples were packaged in aluminum foil, labeled, and placed in a box for transportation. Samples were collected by trained research assistants. Sample collection took place over three time points. Eighty-three percent of samples were collected in November 2017 during the Fall (N= 90) and 17% were collected during the Spring in March/April (N=18).
2.1.3. Youth Population (Study 3): In-Home Collection
Hair was collected in a racially/ethnically heterogeneous participant group of children (American Indian/Alaskan-Native = 1, Black/African-American = 14, White = 56, Hispanic/Latino = 13, and More than 1 race = 2; N= 107). These children resided in the United States (Age: M = 13.12 years; R = 10–16y). Participants were a part of a larger cohort of youth enrolled in the Early Growth and Development Study (EGDS) aimed at understanding the effects of environmental health exposure on youth populations (Leve et al., 2019; Whitesel et al., 2020). Thirty-seven percent of hair samples were collected during the 2019 Summer (N= 39), 30% in the Fall of 2019 (N= 32), 17% during the 2020 Winter (N= 18), and 16% in the Spring of 2020 (N= 17). Hair samples were all initially collected by trained research assistants. Due to COVID-19, hair samples were also collected at home by participant family members. Twenty percent of hair samples (N=21) were collected by family members of youth participants in their home and 80% percent of hair samples (N=85) were collected by research assistants. Collection information was not available for one participant. Families and researchers had access to hair training videos commissioned and produced by the study, described below (SpitLab H.A.I.R. Project, n.d.).
2.2. Hair Collection Supplies:
Supplies for collection varied based on the study (Table 1). At minimum, scissors designed for cutting hair, aluminum foil, an envelope, sanitizer (when collecting from multiple participants), and small elastic bands were necessary for hair collection. Combs, gloves, and ziplock bags were useful depending on the collection site, but these tools were optional and at times caused difficulty. For instance, researchers found gloves increased static electricity especially in medium to long hair which impeded hair banding. Researchers often opted to wash and dry hands between participants. Depending on hair type and length, combs became a hindrance when sectioning into “smaller snips”; it was more efficient to part hair with scissor tips. Plastic airtight storage bags are not recommended because they introduce moisture which degrades hair during transport and storage (Wilson et al., 2007).
Table 1.
Supplies by study
| Supplies | Study 1 | Study 2 | Study 3 |
|---|---|---|---|
|
| |||
| Scissors | ✓ | ✓ | ✓ |
| Aluminum Foil | ✓ | ✓ | ✓ |
| Rubber Bands | ✓ | ✓ | ✓ |
| Ziptop Bags | ✓ | ✓ | |
| Combs | ✓ | ✓* | ✓ |
| Gloves | ✓* | ||
| Metal Hair Clips | ✓ | ✓ | |
| Shipping Box | ✓ | ✓ | ✓ |
| Envelopes | ✓ | ✓ | ✓ |
| Barbicide | ✓ | ||
| Hand Sanitizer | ✓ | ✓ | |
| Scale | ✓ | ||
| Mock Sample | ✓ | ||
Discontinued Use
Collection instructions obtained hair least affected by the sun. Collection from the middle of the scalp or roughly parallel to the center of the ear minimizes embarrassment (SpitLab H.A.I.R. Project, n.d.). To obtain a sample from the mid-section of the scalp, unbound hairstyles are ideal, yet culturally sensitive techniques should be prioritized when hair is styled such that accessessing the middle of the scalp is improbable, as with styles like braids, locs, weaves, slicked/gelled ponytails, or extensions (Manns-James & Neal-Barnett, 2019; Wright et al., 2018) as well as fades and pixie cuts where sufficiently long hair is nearer the top of the head.
2.3. H.A.I.R. Questionnaire and Sample Collection:
Study 2 and Study 3 asked participants to indicate hair type (Figure 1) following Walker’s original hair typing system. Category types included 4a and 4b; 3a and 3b; 2a, 2b, and 2c; and 1a, 1b, and 1c (Walker et al., 1997). We instead described 1, 2, 3, 4 as referring to straight, wavy, curly, and kinky/tightly coiled hair (respectively) with a, b, and c indicating tightness/thickness of the strand or curl within each range. Given that Study 1 consisted of an exclusively Black Kenyan population where hair types leaned toward type 4, we reduced participant burden with a hair type chart limited to type 1, 2a, 2b, 3, and 4a, 4b, 4c, and 4d with type “a” representing a looser curl and “d” representing a tighter curl (Figure 2). No Study 1 participants chose outside of range 4.
Figure 1.

Category types 4a and 4b (Kinky Hair); 3a and 3b (Curly Hair); 2a, 2b, and 2c (Wavy Hair); and 1a, 1b, and 1c (Straight Hair)
Figure 2.

Category types: 1(straight); 2a, 2b (wavy); 3 (curly); and 4a, 4c, 4b, and 4d (kinky)
Five external exposure items from the H.A.I.R questionnaire were used: 1) “Is your hair currently dyed? (Yes, No)”, and if yes, “when was your hair last dyed?”, 2) “Is your hair currently chemically treated (e.g. permed/relaxed/straightened)? (Yes, No)”, and if yes “when was your hair chemically treated (e.g. permed/relaxed/straightened)?”, 3) “How many days a week on average do you blow dry your hair? (1–7)”, 4) “How many times a week on average do you straighten your hair using heat (i.e. flat iron)? (1–7)”, and 5) “How many hours a day on average do you usually spend outside? (1–12)”. Both study 1 and 2 were homogenous in the racial and ethnic make-up of the study participants (Kenyan and Mexican American, respectively) making the population ideal for examining differences in hair type within racial groups.
For Study 2, items on the H.A.I.R. questionnaires were presented in English, Spanish, or both. In Study 3, “How many days a week on average do you blow dry your hair?” and “How many times a week on average do you straighten your hair using heat (i.e. flat iron)?” were given the option of answering “0–7”. The item “How many hours a day on average do you usually spend outside?” ranged from “0–12”. Study 3 only measured items 4 and 5.
Items 3–5 were measured as continuous; however, participant responses were largely categorical. Seventy-eight out of 93 participants reported blow drying their hair never or less than once a week (84%), 152 out of 187 (81%) reporting using heat to style their hair ‘never or less than once a week’, and 90 out of 183 (49%) reported being outside two hours or less. Given reduced variability, items 3–5 were analyzed as functionally dichotomized. For each measure, “0” indicated no use. Heat was coded where “1” indicated heat used to style hair on a daily average. Dryer use was coded where “1” indicated heated dryers were used typically on 1 or more days/week. Dyed was coded so that “1” represented dyed hair. Permed or chemically processed was coded so that “1” represented that hair had been chemically treated. Lastly, Time spent outside was coded where “0” represented 0–2 hours outside on an average day and “1” indicated 3+ hours/day on average spent outdoors.
2.4. Hair Training Videos:
All studies were given access to a series of short training videos created via the EGDS project (link). The videos were used to explain best practices along with materials needed for collection of hair samples (SpitLab H.A.I.R. Project, n.d.). Videos include cutting methods ideal for a range of hair types instead of a ‘one-size fits all’ approach. Created initially as training supplements for researchers, instructional videos became essential during the COVID-19 pandemic when sample collection largely became remote and participants and families benefited from visual aids to successfully collect from home. Videos relevant to studies 1 and 2 hair types were shared with both researchers and participants. For study 2, Spanish closed captioning aided Spanish speaking researchers and participants in future sample collections.
2.5. Lab Protocols:
Segmented samples were washed twice in 5ml of high-performance liquid chromatography (HPLC) isopropanol with constant rotation. Hair was dried under forced air for 3–4 hours in a fume hood. Dried samples measured between 5 and 15 mg depending on their amounts. Weighed samples were ground in a ball mill for 8 minutes at 30hz. The powdered hair was then extracted in 1.5ml of methanol over a 24-hour incubation period using continuous inversion at room temperature. After incubation, samples were centrifuged at 5000rpm for 5 minutes before 1ml of supernatant was transferred to a new clean vial and dried under a nitrogen evaporator in a water bath (50°C). The extract was reconstituted with 150μL of assay diluent, then assayed using commercially available enzyme-immunoassay kits (Salimetrics, PA). Results were converted from μg/dL to picograms/milligrams (pg/mg) before log transformation in final analysis.
All samples for Study 1 were successfully assayed for cortisol. Nineteen samples were assayed at the length of 3cm and recommended weight of 15mg, 5 samples at a length less than 3cm (Range = 1.5cm-2.9cm) at 15mg, 7 samples at the length of 3cm at 10mg, 5 samples at a length less than 3cm (Range = 1.7cm-2.cm) at 10mg, 7 samples at the length of 3cm at 5mg, and finally 4 samples at a length less than 3cm (Range = 1.7cm-2.5cm) at 5mg. All 1A hair type samples for Study 2 were successfully assayed for cortisol at the length of 3cm at 15mg per sample. Study 3 successfully completed 90 percent of participants’ hair assays for 5 hormones.
2.6. Statistical Analysis:
Cumulative hair cortisol (pg/mg) was assessed for normality using visual inspection, skewness (5.00), and kurtosis (27.84). To reduce skewness (1.10) and kurtosis (1.42), hair cortisol was log transformed with the constant 3 added to adjust all values above zero. To test whether external variables influenced hair cortisol, 6 one-way ANOVAs were run in SPSS, where log-transformed hair cortisol was the dependent variable and heat, time outside, dryer, dyed, permed/chemically treated, and weight were independent variables. Post hoc Tukey’s gauged differences across weights of 5, 10, and 15 mg of hair.
To examine differences in hair cortisol between racial groups, hair types, and hair types within each racial group, we ran two separate regressions with cortisol in the total sample as the dependent variable. Race was coded as Black =0, Latino/Hispanic=1, Multi-Racial =2, White=3. Hair type was transformed into an ordinal scale that represented hair types 1a-4d. Given expanded variability in kinky hair types in Study 1, type 4 hair varied from 4a-4d with more limited types across 1–3. Ordinal coding of “Hair Type” resulted in “1a=1, 1b=1.33, 1c=1.66, 2a=2, 2b=2.33, 2c=2.66, 3a=3, 3b=3.5, 4a=4, 4b=4.33, 4c=4.66, and 4d=5.
We then built three univariate models beginning with a within racial group comparison to test differences across race, hair type, and hair type nested within each race. Univariate models were run separately on an interaction between hair type and each racial category in the sample set. While statistically our models predict Black individuals’ cortisol level for type 1a hair, only white and Latino/Hispanic individuals reported type 1a hair. Lastly, we ran regression models including Hair Type, Race, and Race*Hair Type to graph differences in cortisol across hair types, separately by race. For brevity, regression analyses are reported in supplementary material.
3. Results
3.1, 3.2: Cortisol variability within hair types and racial group (Univariate Models)
Regarding hypothesis 1 (H1), we found significant differences in hair cortisol concentrations between hair types (H1) R2=0.16, F(1, 221)=45.23, p<0.001 and hair cortisol concentrations between racial groups (H2), R2=0.06, F(1, 233)=15.97, p<0.001 (Table 3 and Figure 3). As expected, collinearity tests revealed race and hair type were highly nested. Univariate models testing within racial group comparisons (H2) revealed a significant difference in hair cortisol between Black participants and LatinX/Hispanic [F(3, 234)=17.57, p<0.001], Mracial [F(3, 234)=17.57, p=0.004], and White participants [F(3, 234)=17.57, p<.001]. Adding hair type into our model found no significant differences when controlling for hair type between racial groups. Thus, the univariate models suggest no difference in hair cortisol concentrations for type 3 hair collected from a Latino/Hispanic person versus type 3 hair collected from a White person.
Table 3.
Differences in Levels of Cumulative Hair Cortisol(pg/mg) Between Racial Groups, assessed using Univariate Models using combined studies (1,2, and 3)
| β | Std. E | p | Mean | Std. E | |
|---|---|---|---|---|---|
|
| |||||
| Black Intercept | 3.35 | 0.10 | <.01 | 3.35 | 0.10 |
| White | −0.82 | 0.15 | <.01 | 2.53 | 0.11 |
| Mracial | −0.60 | 0.20 | <.01 | 2.75 | 0.18 |
| Latino | −0.87 | 0.12 | <.01 | 2.48 | 0.07 |
| White Intercept | 2.53 | 0.11 | <.01 | 2.53 | 0.11 |
| Black | 0.82 | 0.15 | <.01 | 3.35 | 0.10 |
| Mracial | 0.22 | 0.21 | 0.29 | 2.75 | 0.18 |
| Latino | −0.05 | 0.13 | 0.71 | 2.48 | 0.07 |
| Mracial Intercept | 2.75 | 0.18 | <.01 | 2.75 | 0.18 |
| Black | 0.60 | 0.20 | <.01 | 3.35 | 0.10 |
| White | −0.22 | 0.21 | 0.29 | 2.53 | 0.11 |
| Latino | −0.27 | 0.19 | .15 | 2.48 | 0.07 |
| Latino Intercept | 2.48 | 0.07 | <.01 | 2.48 | 0.07 |
| Black | 0.87 | 0.12 | <.01 | 3.35 | 0.10 |
| White | 0.05 | 0.13 | 0.71 | 2.53 | 0.11 |
| Mracial | 0.27 | 0.19 | 0.15 | 2.75 | 0.18 |
| R2 = .18 | |||||
Figure 3.
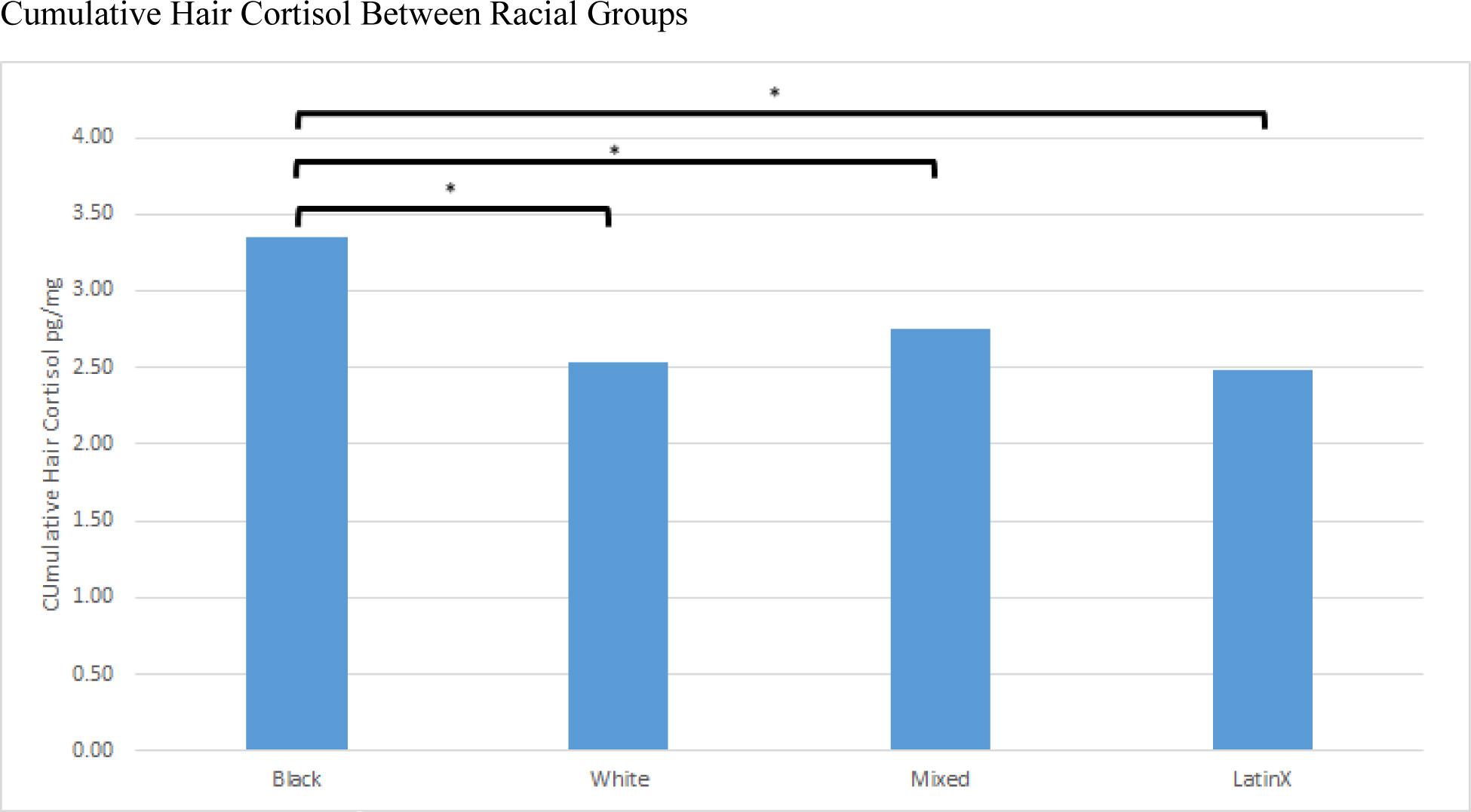
Differences in cumulative hair cortisol
3.3: Higher concentrations of cortisol in hair types across all races
When compared to Black individuals, an interaction between race and hair type showed a significant decrease in hair cortisol concentrations for Multiracial individuals [F(7, 220)=9.86, p=0.044]. There was also a trend for differences with White individuals [F(7, 220)=9.86, p=0.051]. When compared to Latino/Hispanic individuals, both White [F(7, 220)=9.86, p=0.001] and Multiracial [F(7, 220) = 9.86, p= .001] hair type interactions with race showed a significant decrease in hair cortisol concentrations. Within racial groups there were differences in hair cortisol concentrations depending on hair type (H3). However, the impact of hair type on cortisol concentration differed depending on race (Figure 4).
Figure 4:
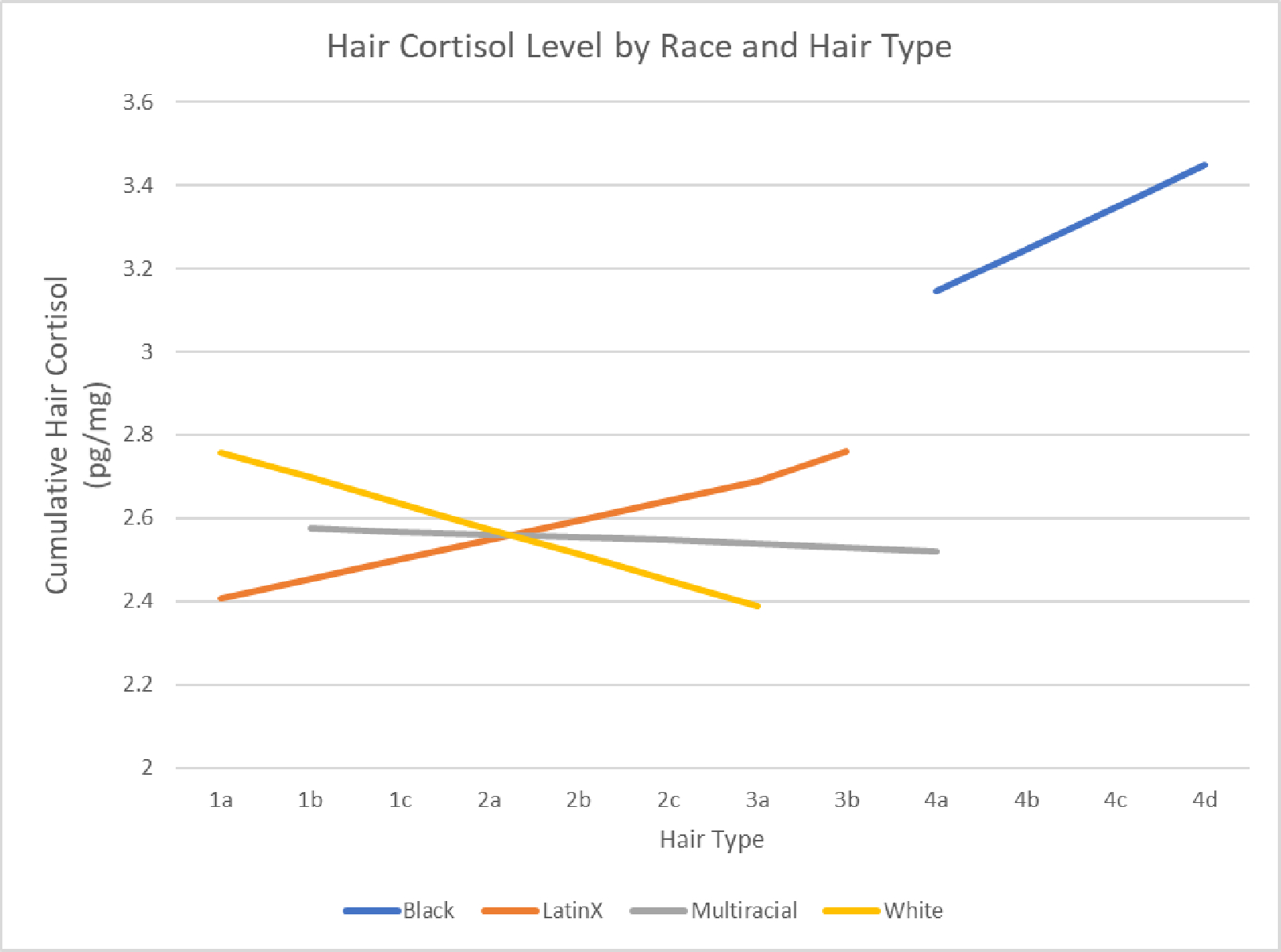
Differences in Hair Cortisol by Hair Type within Race
3.4. External Hair Exposures
Heated Styling, Hair Dryer Use, Hair Dye Use, Hair Treatment Use
In the pooled analysis or within individual studies, there was no significant difference in hair cortisol concentrations between groups who used heat compared to those who did not use heat (H 4.1); dryer versus no dryer use (H4.3); undyed versus dyed hair (H 4.4); and untreated versus permed/chemically treated hair (H 4.5).
Time Spent Outside
None of the studies showed a significant difference in hair cortisol between groups who spent 3+ hours outside compared to those who did not (H 4.2), Study 1: (F(1, 37) = 2.065, p = .159), Study 2: (F(1, 45) = 2.989, p = .091), Study 3: (F(1, 77) = .962, p = .330). After pooling the studies, there was a significant difference in hair cortisol based on time spent outside: (F(1, 163)=9.487, p=.002) (Table 2).
Table 2.
Mean concentration of hair cortisol pg/mg and stand deviation for external hair exposures and weight. Effect Size reported in Cohen’s D for External exposures and R2 for Hair Weight.
| Study 1 | Study 2 | Study 3 | Study 1|2|3 | |||||
|---|---|---|---|---|---|---|---|---|
|
| ||||||||
| Heat | Mean(SD) | Mean(SD) | Mean(SD) | Mean(SD) | ||||
| No | n=38 | 3.26 (.85) | n= 37 | 2.80 (.58) | n= 62 | 2.70 (.80) | n= 137 | 2.88 (.80) |
| Yes | n=1⥈ | 4.83 (-) | n= 14 | 2.86 (.90) | n= 17 | 2.87 (.84) | n= 32 | 2.93 (.90) |
| Total | p= 0.08 | 3.30 (.89) | p= .76 | 2.81 (.67) | p= .45 | 2.73 (.81) | p= .77 | 2.89 (.82) |
| Effect S. | NA⥈ | −.09 | −.21 | −.29 | ||||
| Outside | Study 1|2|3 | |||||||
| No | n= 9 | 2.93 (.73) | n= 25 | 2.63 (.34) | n= 44 | 2.65 (.78) | n= 78 | 2.68 (.67) |
| Yes | n= 30 | 3.41 (.90) | n= 22 | 2.92 (.80) | n= 35 | 2.83 (.84) | n= 87 | 3.05 (.87) |
| Total | p= .16 | 3.30 (.88) | p= .09 | 2.76 (.60) | p= .33 | 2.73 (.81) | p = .002 | 2.88 (.80) |
| Effect S. | −.55 | −.40 | −.23 | −.29 | ||||
| Dryer | Study 1|2 | |||||||
| No | n= 36 | 3.27 (.89) | n= 39 | 2.77 (.58) | - | - | n= 75 | 3.01 (.78) |
| Yes | n= 3⥈ | 3.64 (.70) | n= 12 | 2.96 (.93) | - | - | n= 15 | 3.10 (.91) |
| Total | p= .49 | 3.30 (.88) | p= .38 | 2.81 (.67) | - | - | p= .69 | 3.04 (.80) |
| Effect S. | −.43⥈ | −.27 | - | −.42 | ||||
| Dyed | Study 1|2 | |||||||
| No | n= 37 | 3.34 (.88) | n= 41 | 2.75 (.67) | - | - | n= 78 | 3.03 (.81) |
| Yes | n= 2⥈ | 2.48 (.14) | n= 10 | 3.04 (84) | - | - | n=12 | 2.97 (.79) |
| Total | p= .18 | 3.30 (.88) | p= .18 | 2.81 (.67) | - | - | p= .82 | 3.02 (.80) |
| Effect S. | NA⥈ | .44 | .30 | |||||
| Perm/CT | Study 1|2 | |||||||
| No | n= 33 | 3.27 (.88) | n= 46 | 2.85 (.70) | - | - | n= 79 | 3.02 (.80) |
| Yes | n= 6⥈ | 3.44 (.94) | n= 5⥈ | 2.52 (.06) | - | - | n=11 | 3.02 (.82) |
| Total | p = .67 | 3.30 (.88) | p = .32 | 2.81 (.67) | - | - | p= 1.00 | 3.02 (.80) |
| Effect S. | .20⥈ | −.44 | −.16 | |||||
| Weight | Study 1|3 | |||||||
| 5 | n= 10 | 3.93 (.95) | - | - | - | - | n= 10 | 3.93 (.95) |
| 10 | n= 10 | 3.46 (.63) | - | - | n= 21 | 2.80 (.61) | n= 31 | 3.02 (.68) |
| 15 | n= 21 | 2.90 (.71) | - | - | n= 67 | 2.69 (.83) | n= 88 | 2.74 (.81) |
| Total | p< .01 | 3.29 (.86) | - | - | p= 55 | 2.72 (.78) | p < .01 | 2.71 (.82) |
| Effect S. | .21 | −.007 | .12 | |||||
Note: Cells below N=6 are marked to indicate a low sample size within study. Statistics should be interpreted with caution; data are reported to aid in future meta-analyses. NA=Not applicable.
3.5: Hair Sample Weight
We compared samples weights to understand if hair cortisol concentrations differed (H5) between 5, 10, or 15mg of hair. Study 2 was excluded because only 2 samples were less than 15mg. In the pooled dataset, there was a significant difference in hair cortisol across weights (F(2. 126)=6.667, p<0.001), such that samples assayed at smaller weights had higher hair cortisol. Higher hair cortisol was found in 5mg of hair when compared to 10mg (Mdiff=.916, SE=.287, p<.005) and 15mg (Mdiff=1.19, SE=.263, p<.001). There were no significant differences in cortisol between 10 and 15mg of hair (Mdiff=−.276, SE=0.165, p=0.218) (Table 2). Study 1 reinforced a significant difference in hair cortisol across weights (F(2, 38)=6.622, p<0.001), but Study 3 did not (F(1, 86)=0.355, p=0.553).
5. Discussion
The current study reports how weights, external hair exposures (i.e., heat, time spent outside, chemical treatment, and dyes), and hair type influenced hair cortisol collected in lab, home, and field settings. A hair typing system that is feasible and popular amongst hair-care specialists but novel to scientific endeavors was used to examine hair type and their interactions with race.
5.1. Weight
We found greater hair cortisol concentrations in samples with lower weights and determined that 5mg of hair may not be adequate for assaying cortisol. While mitigation efforts can reduce the impact of small sample weights, such as expanding segmentations to 4–5 centimeters of hair, error is introduced because the association between hormone concentrations diminishes the further away that hair is collected from the scalp (Wang et al., 2018). It is essential for sample weights to consistently be large enough, as well of adequate quality, for successful assays.
Consistent hair sample collection can be achieved through instructions that are clear and easy to understand for participants and lab technicians. High success rates in Study 2 may have been due to practices such as visual aids (e.g. a mock sample of hair) as a means of comparison, hair training collection videos (SpitLab H.A.I.R. Project, n.d.), and extensive training before collection in the field. We recommend researchers practice with mock participants (Ford et al., 2016) to gain comfort with collection. We have also enhanced hair collection with a mobile scale for immediate sample weighing and “false” hair samples of all hair types to use as reference in the field or at home. It is important to consider hair type impacting collection success.
Another consideration is how hair styles influence hair sample collection. Braids, cornrows, locs, weaves, extensions, and styles more common amongst Black populations may require specific collection methods (Wright et al., 2018) or risk systematic missing data and misrepresentation of hormone concentrations via inadequate sample volume. Cultural sensitivity and attention to hair type differences is needed when collecting hair across minority groups. One approach to enhance cultural sensitivity is to collect hair from different regions of the scalp, such as the nape of the neck (SpitLab H.A.I.R. Project, n.d.), which is advantageous when hair is styled where access to the parietal region of the scalp is inaccessible or difficult. Advocating for this approach hinges on learning how hair hormone concentrations vary across areas of the scalp (Meyer & Novak, 2012).
Because hair thickness influences sample weight, the approximation of 100 strands may be adequate for thick hair strands to obtain 100mg, but may be inadequate for thinner hair strands. Erring on the side of collecting large volumes may not be prudent, as willingness to provide hair may be diminished due to discomfort with hair loss. Systematic sources of biases, especially for longitudinal studies interested in multiple hair collections over time, may occur due to attrition from participant refusal and inadequate sample weights. We have found that participant discomfort is reduced by taking “small snips” or “tiny snips” from multiple smaller sections of the hair; this has been especially successful for individuals with type 4 hair (SpitLab H.A.I.R. Project, n.d.). It is important to understand how variability in hair sample weights and hair types can influence study results.
Future studies are needed to confirm correspondence between hair collected from different sections of the head and body. The nape of the neck, facial, leg, or arm hair could be a viable substitute for individuals uncomfortable providing scalp samples as with many hair styles or for bald(ing) individuals. Consulting with hair care professionals about collection protocol design, training and implementation may be beneficial given that their knowledge of hair styling may educate researchers to reduce visibility of missing hair. It is also beneficial to inform participants of hair collections a priori to permit time to prepare. Black participants have been found to be more willing to provide samples between styles and before wash days (Manns-James & Neal-Barnett, 2019).
5.2. External Exposure
The recommendation for sample collection from specific regions of the scalp is based on evidence that exposure to sunlight degrades hormones in hair (Feng et al., 2019; Wester et al., 2016). Indeed, our pooled-study analyses found participants who spent more time outside – and presumably more outdoor sun exposure – had higher cortisol. Results were not aligned with current studies in which lower hair cortisol concentrations are associated with greater sunlight exposure (Feng et al., 2019; Wester et al., 2016). While time spent outdoors may be a good proxy for sunlight-based degradation, it may be pertinent to consider sources of degradation. Sweat exposure, washing frequency, exercise, and hair style all influence how the outdoors may impact individual differences in hair hormone concentrations. For instance, while washing decreases sweat-related increases in hair cortisol (Grass et al., 2015), certain hair styles (e.g. braids, locs, and cornrows) require little shampooing. Hair kept beneath “wig styles” may have elevated sweat exposure and less sun exposure. Thus, time spent outdoors may be a useful yet insufficient proxy for degradation and the meaning and impact of sun, sweat, and washing may differ widely across populations of interest.
Prior literature has found that segmented hair exposed to bleach, dyes, and heat decreased hair cortisol. Our study did not find a connection between cortisol concentrations and permed/chemically straightened treatments, dyed hair, heated-treated, nor blow dried hair. This may be due to limits of self-reporting hair exposures instead of systemically subjecting archived samples to bleach and dyes (Hoffman et al., 2014) and points to a need to parse out different “treatments” (e.g. heat versus chemical).
Studies have revealed heat-based treatment differences with dichotomous scales in predominantly White groups (Fischer et al., 2017). Conclusions about Black participants could not be generalizable as sample collection was inadequate in 71% of Black participants (Fischer et al., 2017). Studies where heat is collected as a continuous variable reported no differences in “frequent” users of heat for styling (over three times a week) from those who style hair less often with heat (Liu et al., 2016). Regardless, type of heat treatment (e.g. flat ironing or blow dryer) should be recorded in an abundance of caution so future studies can maximize replicability.
Regarding hair treatments, meta-analysis are inconsistent, revealing only trends for lower concentrations based on treatments (Stalder et al., 2017) in predominantly White populations (Manenschijn et al., 2011; Stalder et al., 2012; Stalder et al., 2013; Staufenbiel et al., 2014). Studies have found within-group differences in Black individuals where permed hair had lower cortisol (Wosu et al., 2015), but other studies examining Black populations have not shown differences in dyed, permed, nor straightened hair (Lehrer et al. 2016). Conclusions about hair treatments are inconclusive due to race and potentially hair type. One reason for variability in the effect of “treatment” is that the measure is inconsistent across studies, lumping dyed hair, permed hair, and heat treated hair into the same category. Treatments such as hair bleaching and chemical straightening may be considerably harsher on hair than other treatments (Lee, 2011; Joseph et al., 1996; Casati et al., 2021). Considering that some groups may frequently use different types of treatment (e.g. chemical straighteners could be more common in Black individuals or bleaching treatments could be more common amongst women), it may be important to inquire about the type and number of chemical treatments. For instance, cortisol may be differently influenced by dye if hair dye steps also involve chemical bleaching. Future studies should detail hair treatments and consider parsing heat-based from chemical treatments.
Reporting time since last treatment and frequency of chemical treatment may be important for controlling the accumulation of these exposure in each hair segment. Hair should be segmented close to the scalp (Wang et al., 2018), so hair treatments that occurred months prior may not be influential. A continuous variable that queries frequency of use of heat and chemical based hair styles/treatments may increase reliability. Finally, common heat-based styles (blow drying and flat ironing) should record frequency and time since last heat exposure. We acknowledge the limitation to parse multiple hair exposure categories and that collapsing dyed, chemically treated, and heat treated hair may have impacted findings. Larger studies are needed.
5.3. Hair Type
We were able to find only one study which examined hair shape and cortisol based on hair type (Wosu et al. 2015), despite evidence for differences in lipid concentration depending on the shape of hair strands (Martí et al. 2016). Wosu and colleagues (2015) found higher cortisol in tightly curled hair and lower concentrations in “wavy hair”, which align with expected outcomes for greater lipid absorption in tightly coiled hair (Martí et al. 2016). However, race was not considered which may have conflated tightly coiled hair with Black populations.
H1 was supported such that hair shape, based on the Andre Walker (Walker et al., 1997) hair typing scale, impacted cortisol levels. These findings remained significant even when accounting for race, a major contributor to cortisol levels (Hajat et al., 2010; O’Brien et al., 2013) (H2 and H3). The impact of hair type replicated across Black, White, Multiracial, and Latino/Hispanic samples, but with a twist. Whereas both race and hair type impacted hair cortisol, Black and Latino/Hispanic participants had higher hair cortisol in tighter coiled hair types whereas Multiracial, and to a lesser extent White, participants had greater hair cortisol with straighter hair types. Supplemental regression analysis within racial groups showed the impact of hair type within Multiracial groups was significant and trended within White individuals in the same direction. Our findings generally supported Wasu and colleagues (Wosu et al., 2015), as well as mechanisms for effects based on lipid solubility. Yet, our findings add within racial group differences and interactions between hair type and racial groups that influence cortisol deposits.
Our finding for differences in cortisol concentrations by both racial group and hair type may be indirect or inadvertent evidence of colorism. Colorism is defined as the allocation of societal privileges based on how light or dark a person’s skin tone is (Burke, 2015; Norwood, 2014). Colorism can also extend to other features. In addition to skin tone, curlier or kinkier hair may be a secondary indicator of darker skin tones which impact societal treatment in Latino/Hispanic (Atroon et al., 2019; Charles, 2021; M. A. Fuentes et al., 2021) and Black populations (Moffitt, 2020; Norwood, 2014; Stalnaker- Shofner, 2020). This theory purports that curlier or kinkier hair in Latino/Hispanic and Black individuals corresponded with higher cortisol to indicate stress-related effects of greater discrimination. Such colorism effects would not be expected in White nor, possibly, in Multiracial populations who pass as White. Indeed, we found that curlier hair showed lower cortisol concentrations for these racial groups. It is important that future studies consider how cortisol may get under the skin differently not only by race but by skin tone and hair type. Future studies should test hair cortisol differences within populations with similar ethnic background and hair types but divergent skin tones. Future studies should also consider collecting kinkier hair samples in lighter individuals and looser curls in darker skinned individuals alongside variable hair types within all racial groups. This may reveal whether hair type and skin tone can shed light on a biological marker (cumulative cortisol) of colorism and classism inherent in caste systems (Khubchandani et al., 2018; Pulla et al., 2020; Wilkerson, 2020).
In addition to hair type/shape, another reason for racial group differences in cortisol concentrations may be variability in rates of growth by season and hair type, hair density, or hair loss (Loussouarn et al., 2016; V. A. Randall, 1994). Studies have found group differences in hair growth depending on geographic origins and density variability within racial groups (Loussouarn et al., 2016). Research that tests between group differences in hair hormones must consider that variables such as region, time of year that hair is collected (season), ethnic group, and hair type may impact absorption of hormones both within and between groups. To date, there is no research that examines how hair density and growth rate influence hormone absorption. As alluded to above, time spent outdoors as a proxy for sun exposure may not provide enough information on regional effects of sunlight exposure, such as distance from the equator or intensity of sun exposure (Sukmawati & Pramudya, 2019) as well as the amount of available time for sun exposure while outdoors (Caccin et al., 1970).
Rather than “reinvent the wheel” (Kymberlee et al., 2018), the Andre Walker HTS used by haircare professionals is a good starting point, yet it is not the only step needed for scientific studies on hair to reduce ethnoracial centrism. Further differentiating/expanding type 4 hair to include subtypes a-d was helpful (e.g., our Kenyan sample) for increasing variability within hair type 4. Researchers should also reduce racist language by, for example, eliminating hair descriptors such as mongoloid, negroid, or African. Culturally sensitive language surrounding hair scaled 1–4 should also clarify to participants that straight hair is not considered “better” because it is labeled as number 1. Instead, the number system indicates the amount of curl and the underlying mechanism is lipid absorption. Adopting a system that categorizes hair in a simple, respectful, and consistent way will be essential to answering questions surrounding control variables needed to account for methodological gaps in hair hormone absorption. For ease of understanding, we recommend describing types 1, 2, 3, 4 as referring to straight, wavy, curly, and kinky/tightly coiled hair (respectively) with a, b, c, d indicating thickness of the hair strand within each range. This helps eliminate confusion caused by different meanings of the a, b, and c subtypes, such as “a” being used to describe “thickness” in type 1 hair but “coil tightness” in type 4 hair.
5.4. Limitations of Analysis
We were unable to consider hair color as a potential contributing factor or in interaction with hair type. Prior research on hair binding of cocaine showed similar levels within black/brown hair regardless of race while blonde participants showed less binding (Joseph et al., 1996). Lighter hair shades are typically more porous and may be less resistant to extraneous factors. Future studies should measure hair color or consider strategies for recording this information around the time of assay. The present study did not have enough variability to gauge differences by hair color within hair type and race, so this remains an important future direction.
Effect sizes for external control variables were small and p-values were somewhat underwhelming. Even when statistically significant, this poses the question of whether findings were meaningful. For example, low effect sizes were found for sample weight and even measures with higher effect sizes such as dryer use were still underpowered and did not necessarily yield statistical significance. Our most robust findings, where time spent outside culminated in greater hair cortisol, may have been an indication of sweat exposure. This exploratory study identifies a need for larger analysis of external hair exposures that are not collapsed across differing hair treatments (e.g. chemical straightening versus bleaching). Contrarily, racial group, hair type, and within racial group hair type differences in cortisol were revealed to illustrate the importance of hair type when collecting hair. Our recommendation is that individual studies should err on the side of caution and collect these potential control metrics prior to evaluating specific impact.
5.5. Conclusion
Laboratory methodologies for hair cortisol assays have allowed this novel biospecimen to be a frequent tool in the arsenal of stress research. Laboratory rigor must be accompanied by feasible, reproducible and valid collection protocols and adequate control metrics. The current study revealed that hair type and external exposures impact cumulative hair cortisol concentrations. It is beneficial to explore the meaning and breadth of external exposures and to be considerate of the full range of hair types with the ultimate goal to approach hair collection and analysis with a wide cultural lens that does not assume homogeneity in the way hair is exposed to external variables.
Supplementary Material
Table 4:
Differences in Levels of Cumulative Hair Cortisol (pg/mg) Between Racial Groups and Hair Type (HT), assessed with Univariate Models using combined studies (1,2, and 3).
| β | Std. E | p | Mean | Std. E | |
|---|---|---|---|---|---|
|
| |||||
| Black Intercept | 2.77 | 0.47 | <.01 | 3.06 | 0.25 |
| Hair Type | 0.14 | 0.11 | 0.21 | X | X |
| White | −0.45 | 0.32 | 0.16 | 2.61 | 0.13 |
| Mracial | −0.48 | 0.26 | 0.07 | 2.58 | 0.22 |
| LatinX | −0.46 | 0.34 | 0.18 | 2.60 | 0.11 |
| White Intercept | 2.31 | 0.22 | <.01 | 2.61 | 0.13 |
| Hair Type | 0.14 | 0.11 | 0.21 | - | - |
| Black | 0.45 | 0.32 | 0.16 | 3.06 | 0.25 |
| Mracial | −0.02 | 0.27 | 0.93 | 2.58 | 0.22 |
| Latin X | 0.01 | 0.14 | 0.94 | 2.61 | 0.13 |
| Mracial Intercept | 2.29 | 0.38 | <.01 | 2.58 | 0.22 |
| Hair Type | 0.14 | 0.11 | 0.21 | - | - |
| Black | 0.48 | 0.26 | 0.07 | 3.06 | 0.25 |
| White | 0.02 | 0.27 | 0.93 | 2.61 | 0.13 |
| LatinX | 0.01 | 0.28 | 0.96 | 2.60 | 0.11 |
| LatinX Intercept | 2.30 | 0.16 | <.01 | 2.60 | 0.11 |
| Hair Type | 0.14 | 0.11 | 0.21 | - | - |
| Black | 0.46 | 0.34 | 0.18 | 3.06 | 0.25 |
| White | 0.01 | 0.14 | 0.94 | 2.61 | 0.13 |
| Mracial | −0.01 | 0.28 | 0.96 | 2.58 | 0.22 |
| R2 = .17, HT = 2.17 | |||||
Table 5:
Differences in Levels of Cumulative Hair Cortisol(pg/mg) Between Racial Groups, Hair Type (HT) and Race × Hair Type interaction, assessed with Univariate Models using combined studies (1,2, and 3).
| β | Std. E | p | Mean | Std. E | ||
|---|---|---|---|---|---|---|
|
| ||||||
| Black Intercept | 1.41 | 1.32 | 0.29 | Black | 2.39 | 0.66 |
| White | 1.59 | 1.37 | 0.25 | White | 2.42 | 0.15 |
| Mracial | 2.21 | 1.48 | 0.14 | Mracial | 2.97 | 0.26 |
| Latino | 0.26 | 1.35 | 0.84 | LatinX | 2.98 | 0.15 |
| Hair Type | 0.45 | 0.31 | 0.14 | - | - | - |
| White × HT | −0.72 | 0.37 | 0.051 | - | - | - |
| Mracial × HT | −0.75 | 0.37 | 0.04 | - | - | - |
| Latino × HT | 0.15 | 0.35 | 0.66 | - | - | - |
| R2 = .22 | ||||||
| White Intercept | 3.00 | 0.36 | 0.00 | White | 2.42 | 0.15 |
| Black | −1.59 | 1.37 | 0.25 | Black | 2.39 | 0.66 |
| Mracial | 0.62 | 0.76 | 0.41 | Mracial | 2.97 | 0.26 |
| Latino | −1.32 | 0.43 | <.01 | LatinX | 2.98 | 0.15 |
| Hair Type | −0.27 | 0.20 | 0.18 | - | - | - |
| Black × HT | 0.72 | 0.37 | 0.051 | - | - | - |
| Mracial × HT | −0.03 | 0.29 | 0.91 | - | - | - |
| Latino × HT | 0.87 | 0.26 | <.01 | - | - | - |
| Mracial Intercept | 3.62 | 0.66 | <.01 | Mracial | 2.97 | 0.26 |
| Black | −2.21 | 1.48 | 0.14 | Black | 2.39 | 0.66 |
| White | −0.62 | 0.76 | 0.41 | White | 2.42 | 0.15 |
| Latino | −1.95 | 0.71 | 0.01 | LatinX | 2.98 | 0.15 |
| Hair Type | −0.30 | 0.21 | 0.15 | - | - | -- |
| Black × HT | 0.75 | 0.37 | 0.04 | - | - | - |
| White × HT | 0.03 | 0.29 | 0.91 | - | - | - |
| Latino × HT | 0.90 | 0.27 | <.01 | - | - | - |
| Latino Intercept | 1.68 | 0.24 | <.01 | LatinX | 2.98 | 0.15 |
| Black | −0.26 | 1.35 | 0.84 | Black | 2.39 | 0.66 |
| White | 1.32 | 0.43 | <.01 | White | 2.42 | 0.15 |
| Mracial | 1.95 | 0.71 | 0.01 | Mracial | 2.97 | 0.26 |
| Hair Type | 0.60 | 0.17 | <.01 | - | - | - |
| Black × HT | −0.15 | 0.35 | 0.66 | - | - | - |
| White × HT | −0.87 | 0.26 | <.01 | - | - | - |
| Mracial × HT | −0.90 | 0.27 | <.01 | - | - | - |
| R2= .22, HT= 2.17 | ||||||
Highlights:
Cortisol levels in 5mg of hair were higher when compared to 10mg and 15mg weights
Higher hair cortisol levels were found in samples with more outdoor exposure
Black hair types had higher cortisol than White, LatinX, and Multiracial hair types
Hair types differed in levels of cortisol and varied depending on racial group
For White and Multiracial participants straight hair had higher cortisol than curly hair
Funding and Acknowledgments:
Participants in Study 1 were recruited from the Kenya Study and was supported by the Eunice Kennedy Shriver National Institute of Child Health & Human Development of the National Institutes of Health under Award Number R00HD093798. The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health. The funders had no role in the study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Participants in Study 2 were recruited from a larger research project, Abriendo Caminos, that was supported by the Agriculture and Food Research Initiative Competitive grant no. 2015–68001-23248 from the US Department of Agriculture National Institute of Food and Agriculture under the Childhood Obesity Prevention Challenge Area, and led by Margarita Teran-Garcia, MD, PhD (PI) at University of Illinois at Urbana-Champaign. Kimberly Greder, PhD, Iowa State University, served as the PD for Abriendo Caminos in Iowa. Collection and analysis of hair cortisol data for Study 2 was supported by the College of Human Sciences Collaborative Seed Grant Program (Kimberly Greder, PI; Marian Kohut, Co-PI; Elizabeth Shirtcliff, Co-PI) and a Strategic Initiative Grant from the Vice President for Extension and Outreach at Iowa State University (Kimberly Greder (PI), Himar Hernandez (Co-PI).
R01 DA045108. 4/1/2018–1/31/2023. Participants in Study 3 were recruited from an adoption study of the development of early substance use: the joint roles of genetic influences, prenatal risk, rearing environment, and pubertal maturation”. Type: R01. Principal Investigator: Jenae Neiderhiser. Agency: National Institute of Drug Abuse. Role: Co-Investigator. Total Direct costs: $2,892,828. Total ISU Direct Cost: $595,980.
Special thanks to Jenny Phan, and Tor Finseth for lending an ear to questions and giving encouraging feedback. Ebony Williams contributed as our diversity and inclusion consultant and provided valuable feedback on the use of sensitive language.
Footnotes
Multiracial is used interchangeably with “Mixed” and the authors acknowledge the complexity of each term. Ultimately multiracial was decided on to strive for cultural sensitivity. 2 We further recognize the cultural complexity in using Latino/a/e/X as well as the controversy of their use within both Spanish speaking and academic communities.
Publisher's Disclaimer: This is a PDF file of an article that has undergone enhancements after acceptance, such as the addition of a cover page and metadata, and formatting for readability, but it is not yet the definitive version of record. This version will undergo additional copyediting, typesetting and review before it is published in its final form, but we are providing this version to give early visibility of the article. Please note that, during the production process, errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References:
- Adam EK, Heissel JA, Zeiders KH, Richeson JA, Ross EC, Ehrlich KB, Levy DJ, Kemeny M, Brodish AB, Malanchuk O, Peck SC, Fuller-Rowell TE, & Eccles JS (2015). Developmental histories of perceived racial discrimination and diurnal cortisol profiles in adulthood: A 20-year prospective study. Psychoneuroendocrinology, 62, 279–291. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Africa GSW (1908). Herero and Namaqua genocide. Target, 1904, 1. [Google Scholar]
- Atroon N, Eteka F, Spikes M, & Williams S (2019). Do I GOT too Much Melanin?? (Asking for a friend): The Effects of Colorism within the Latinx Community. Iowa State Conference on Race and Ethnicity, 20(1). https://www.iastatedigitalpress.com/iscore/article/id/99/print/ [Google Scholar]
- Azevedo A, Bailey L, Bandeira V, Dehnhard M, Fonseca C, de Sousa L, & Jewgenow K (2019). Age, sex and storage time influence hair cortisol levels in a wild mammal population. PloS One, 14(8), e0221124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bates R, Salsberry P, & Ford J (2017). Measuring Stress in Young Children Using Hair Cortisol: The State of the Science. Biological Research for Nursing, 19(5), 499–510. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bhopal R (2007). The beautiful skull and Blumenbach’s errors: the birth of the scientific concept of race. BMJ, 335(7633), 1308–1309. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boumba VA, Ziavrou KS, & Vougiouklakis T (2006). Hair as a biological indicator of drug use, drug abuse or chronic exposure to environmental toxicants. International Journal of Toxicology, 25(3), 143–163. [DOI] [PubMed] [Google Scholar]
- Burke J (2015). Colourism as an intra-racial phenomenon: The case of Tanzania. African Studies Association of Australasia and the Pacific (AFSAAP) 37th Annual Conference, 1–7. [Google Scholar]
- Caccin B, Falciani R, Moschi G, & Rigutti M (1970). Temperature difference between the equator and the poles of the sun. In Solar Physics (Vol. 13, Issue 1, pp. 33–41). 10.1007/bf00963940 [DOI] [Google Scholar]
- Cell Editorial Team. (2020). Science Has a Racism Problem. Cell, 181(7), 1443–1444. [DOI] [PubMed] [Google Scholar]
- Charles J (2021). Colorism and the Afro-Latinx Experience: A Review of the Literature. Hispanic Journal of Behavioral Sciences, 43(1–2), 8–31. [Google Scholar]
- Coderch L, Oliver MA, Carrer V, Manich AM, & Martí M (2019). External lipid function in ethnic hairs. Journal of Cosmetic Dermatology, 18(6), 1912–1920. [DOI] [PubMed] [Google Scholar]
- Coon CS (1963). Origin of races. Science, 140(3563), 208. [DOI] [PubMed] [Google Scholar]
- Correa S. M. de S. (2011). History, memory, and commemorations: on genocide and colonial past in South West Africa. Revista Brasileira de Historia / Orgao Da Associacao Nacional Dos Professores Universitarios de Historia, ANPUH, 31(61), 85–103. [Google Scholar]
- Cruz CF, Fernandes MM, Gomes AC, Coderch L, Martí M, Méndez S, Gales L, Azoia NG, Shimanovich U, & Cavaco-Paulo A (2013). Keratins and lipids in ethnic hair. International Journal of Cosmetic Science, 35(3), 244–249. [DOI] [PubMed] [Google Scholar]
- Davenport MD, Tiefenbacher S, Lutz CK, Novak MA, & Meyer JS (2006). Analysis of endogenous cortisol concentrations in the hair of rhesus macaques. General and Comparative Endocrinology, 147(3), 255–261. [DOI] [PubMed] [Google Scholar]
- De la Mettrie R, Saint-Léger D, Loussouarn G, Garcel A, Porter C, & Langaney A (2007). Shape variability and classification of human hair: a worldwide approach. Human Biology, 79(3), 265–281. [DOI] [PubMed] [Google Scholar]
- Dettmer AM, Novak MA, Meyer JS, & Suomi SJ (2014). Population density-dependent hair cortisol concentrations in rhesus monkeys (Macaca mulatta). Psychoneuroendocrinology, 42, 59–67. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ebling FJ (1976). Hair. The Journal of Investigative Dermatology, 67(1), 98–105. [DOI] [PubMed] [Google Scholar]
- Eddy MW (1938). HAIR CLASSIFICATION. Proceedings of the Pennsylvania Academy of Science, 12, 19–26. [Google Scholar]
- Engert V, Ragsdale AM, & Singer T (2018). Cortisol stress resonance in the laboratory is associated with inter-couple diurnal cortisol covariation in daily life. Hormones and Behavior, 98, 183–190. [DOI] [PubMed] [Google Scholar]
- Evans AD (2008). Race Made Visible: The Transformation of Museum Exhibits in Early-Twentieth-Century German Anthropology. German Studies Review, 31(1), 87–108. [Google Scholar]
- Feng X-L, Che H-L, Ning X, Ba X-Y, Li J, Zhang J-F, Wang Y, Hu Z-F, Hu X-T, & Ren X-F (2019). Direct sunlight exposure reduces hair cortisol levels in rhesus monkeys (Macaca mulatta). Zoological Research, 40(6), 583–586. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fischer S, Duncko R, Hatch SL, Papadopoulos A, Goodwin L, Frissa S, Hotopf M, & Cleare AJ (2017). Sociodemographic, lifestyle, and psychosocial determinants of hair cortisol in a South London community sample. Psychoneuroendocrinology, 76, 144–153. [DOI] [PubMed] [Google Scholar]
- Ford JL, Boch SJ, & McCarthy DO (2016). Feasibility of Hair Collection for Cortisol Measurement in Population Research on Adolescent Health. Nursing Research, 65(3), 249–255. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fuentes A (2021). Biological anthropology’s critical engagement with genomics, evolution, race/racism, and ourselves: Opportunities and challenges to making a difference in the academy and the world. American Journal of Physical Anthropology, 175(2), 326–338. [DOI] [PubMed] [Google Scholar]
- Fuentes MA, Reyes-Portillo JA, Tineo P, Gonzalez K, & Butt M (2021). Skin Color Matters in the Latinx Community: A Call for Action in Research, Training, and Practice. Hispanic Journal of Behavioral Sciences, 43(1–2), 32–58. [Google Scholar]
- Gidlow CJ, Randall J, Gillman J, Silk S, & Jones MV (2016). Hair cortisol and self-reported stress in healthy, working adults. Psychoneuroendocrinology, 63, 163–169. [DOI] [PubMed] [Google Scholar]
- Gow R, Thomson S, Rieder M, Van Uum S, & Koren G (2010). An assessment of cortisol analysis in hair and its clinical applications. Forensic Science International, 196(1–3), 32–37. [DOI] [PubMed] [Google Scholar]
- Grass J, Kirschbaum C, Miller R, Gao W, Steudte-Schmiedgen S, & Stalder T (2015). Sweat-inducing physiological challenges do not result in acute changes in hair cortisol concentrations. Psychoneuroendocrinology, 53, 108–116. [DOI] [PubMed] [Google Scholar]
- Greff MJE, Levine JM, Abuzgaia AM, Elzagallaai AA, Rieder MJ, & van Uum SHM (2019). Hair cortisol analysis: An update on methodological considerations and clinical applications. Clinical Biochemistry, 63, 1–9. [DOI] [PubMed] [Google Scholar]
- Hajat A, Diez-Roux A, Franklin TG, Seeman T, Shrager S, Ranjit N, Castro C, Watson K, Sanchez B, & Kirschbaum C (2010). Socioeconomic and race/ethnic differences in daily salivary cortisol profiles: the multi-ethnic study of atherosclerosis. Psychoneuroendocrinology, 35(6), 932–943. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hall DL, Blyler D, Allen D, Mishel MH, Crandell J, Germino BB, & Porter LS (2011). Predictors and patterns of participant adherence to a cortisol collection protocol. Psychoneuroendocrinology, 36(4), 540–546. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hayes LD, Grace FM, Baker JS, & Sculthorpe N (2015). Exercise-induced responses in salivary testosterone, cortisol, and their ratios in men: a meta-analysis. Sports Medicine, 45(5), 713–726. [DOI] [PubMed] [Google Scholar]
- Hill Golden S, Sánchez BN, Desantis AS, Wu M, Castro C, Seeman TE, Tadros S, Shrager S, & Diez Roux AV (2014). Salivary cortisol protocol adherence and reliability by socio-demographic features: the Multi-Ethnic Study of Atherosclerosis. Psychoneuroendocrinology, 43, 30–40. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jackson JP (2001). “In Ways Unacademical”: The Reception of Carleton S. Coon’s The Origin of Races. Journal of the History of Biology, 34(2), 247–285. [Google Scholar]
- Joseph RE Jr, Su TP, & Cone EJ (1996). In vitro binding studies of drugs to hair: influence of melanin and lipids on cocaine binding to Caucasoid and Africoid hair. Journal of Analytical Toxicology, 20(6), 338–344. [DOI] [PubMed] [Google Scholar]
- Khubchandani J, Soni A, Fahey N, Raithatha N, Prabhakaran A, Byatt N, Moore Simas TA, Phatak A, Rosal M, Nimbalkar S, & Allison JJ (2018). Caste matters: perceived discrimination among women in rural India. Archives of Women’s Mental Health, 21(2), 163–170. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koch SL, Tridico SR, Bernard BA, Shriver MD, & Jablonski NG (2020). The biology of human hair: A multidisciplinary review. American Journal of Human Biology: The Official Journal of the Human Biology Council, 32(2), e23316. [DOI] [PubMed] [Google Scholar]
- Koren L, Mokady O, Karaskov T, Klein J, Koren G, & Geffen E (2002). A novel method using hair for determining hormonal levels in wildlife. Academic Press. http://www.tau.ac.il/lifesci/departments/zoology/members/geffen/documents/40AnimBehav2002.pdf [Google Scholar]
- Kudielka BM, Buske-Kirschbaum A, Hellhammer DH, & Kirschbaum C (2004). HPA axis responses to laboratory psychosocial stress in healthy elderly adults, younger adults, and children: impact of age and gender. Psychoneuroendocrinology, 29(1), 83–98. [DOI] [PubMed] [Google Scholar]
- Kudielka BM, Gierens A, Hellhammer DH, Wüst S, & Schlotz W (2012). Salivary cortisol in ambulatory assessment—some dos, some don’ts, and some open questions. Psychosomatic Medicine, 74(4), 418–431. [DOI] [PubMed] [Google Scholar]
- Kymberlee H, Adesola A, & Prajjwhal D (2018). Towards Creation of a Curl Pattern Recognition System. Proceedings of the International Conference on Image Processing, Computer Vision, and Pattern Recognition (IPCV), 30–33. [Google Scholar]
- Leve LD, Neiderhiser JM, Ganiban JM, Natsuaki MN, Shaw DS, & Reiss D (2019). The Early Growth and Development Study: A Dual-Family Adoption Study from Birth Through Adolescence. Twin Research and Human Genetics: The Official Journal of the International Society for Twin Studies, 22(6), 716–727. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Loussouarn G, Garcel A-L, Lozano I, Collaudin C, Porter C, Panhard S, Saint-Léger D, & de La Mettrie R (2007). Worldwide diversity of hair curliness: a new method of assessment. International Journal of Dermatology, 46 Suppl 1, 2–6. [DOI] [PubMed] [Google Scholar]
- Loussouarn G, Lozano I, Panhard S, Collaudin C, El Rawadi C, & Genain G (2016). Diversity in human hair growth, diameter, colour and shape. An in vivo study on young adults from 24 different ethnic groups observed in the five continents. European Journal of Dermatology: EJD, 26(2), 144–154. [DOI] [PubMed] [Google Scholar]
- Manns-James L, & Neal-Barnett A (2019). Development of a culturally informed protocol for hair cortisol sampling in Black women. Public Health Nursing, 36(6), 872–879. [DOI] [PubMed] [Google Scholar]
- Martí M, Barba C, Manich AM, Rubio L, Alonso C, & Coderch L (2016). The influence of hair lipids in ethnic hair properties. International Journal of Cosmetic Science, 38(1), 77–84 [DOI] [PubMed] [Google Scholar]
- Matiegka J, & Malý J (1929). Negroid hair in Central Europe. L’Anthropologie, 7(1/2), 108–118. [Google Scholar]
- McEwen BS (1998). Stress, adaptation, and disease. Allostasis and allostatic load. Annals of the New York Academy of Sciences, 840, 33–44. [DOI] [PubMed] [Google Scholar]
- Melber H (2017). Genocide matters-negotiating a Namibian-German past in the present. https://repository.up.ac.za/handle/2263/64266
- Meyer JS, & Novak MA (2012). Minireview: Hair cortisol: a novel biomarker of hypothalamic-pituitary-adrenocortical activity. Endocrinology, 153(9), 4120–4127. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mieczkowski T (1999). Drug Testing Technology: Assessment of Field Applications. CRC Press. [Google Scholar]
- Moffitt KR (2020). “Light-skinned people always win”: An Autoethnography of Colorism in a Mother–Daughter Relationship. Women, Gender, and Families of Color, 8(1), 65–86. [Google Scholar]
- Munck A, Guyre PM, & Holbrook NJ (1984). Physiological functions of glucocorticoids in stress and their relation to pharmacological actions. Endocrine Reviews, 5(1), 25–44. [DOI] [PubMed] [Google Scholar]
- Norwood KJ (2014). Colorism:(Still) Getting Away With Racism. https://psqtest.typepad.com/blogPostPDFs/Colorism_9-8-2014.pdf
- O’Brien KM, Tronick EZ, & Moore CL (2013). Relationship between hair cortisol and perceived chronic stress in a diverse sample. Stress and Health: Journal of the International Society for the Investigation of Stress, 29(4), 337–344. [DOI] [PubMed] [Google Scholar]
- Onishi N, & Eddy M (2021, May 28). A Forgotten Genocide: What Germany Did in Namibia, and What It’s Saying Now. The New York Times. https://www.nytimes.com/2021/05/28/world/europe/germany-namibia-genocide.html
- Palmer-Bacon J, Willis-Esqueda C, & Spaulding WD (2020). Stress, trauma, racial/ethnic group membership, and HPA function: Utility of hair cortisol. The American Journal of Orthopsychiatry, 90(2), 193–200. [DOI] [PubMed] [Google Scholar]
- Pardridge WM, & Mietus LJ (1979). Transport of steroid hormones through the rat blood-brain barrier. Primary role of albumin-bound hormone. The Journal of Clinical Investigation, 64(1), 145–154. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pulla V, Carter E, & Bhatt S (2020). At the intersections of discrimination of caste and beyond. In Discrimination, Challenge and Response (pp. 17–30). Springer International Publishing. [Google Scholar]
- Randall M, Stern A, & Su Y (2021). Five Ethical Risks to Consider before Filling Missing Race and Ethnicity Data. https://www.urban.org/sites/default/files/publication/103830/five-ethical-risks-to-consider-before-filling-missing-race-and-ethnicity-data-workshop-findings_0.pdf
- Randall VA (1994). Androgens and human hair growth. Clinical Endocrinology, 40(4), 439–457. [DOI] [PubMed] [Google Scholar]
- Russell C (2021). Bioethicists Should Be Helping Scientists Think About Race. Journal of Bioethical Inquiry, 18(1), 109–111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Russell E, Koren G, Rieder M, & Van Uum S (2012). Hair cortisol as a biological marker of chronic stress: current status, future directions and unanswered questions. Psychoneuroendocrinology, 37(5), 589–601. [DOI] [PubMed] [Google Scholar]
- Sanchez A, Bao J, & Greder K (2019). P163 Enhancing Family Mealtime Environments and Routines Among Mexican Immigrant Families Through Abriendo Caminos. Journal of Nutrition Education and Behavior, 51(7, Supplement), S106. [Google Scholar]
- Sauvé B, Koren G, Walsh G, Tokmakejian S, & Van Uum SHM (2007). Measurement of cortisol in human hair as a biomarker of systemic exposure. Clinical and Investigative Medicine. Medecine Clinique et Experimentale, 30(5), E183–E191. [DOI] [PubMed] [Google Scholar]
- Schweikert HU, & Wilson JD (1974). Regulation of human hair growth by steroid hormones. II. Androstenedione metabolism in isolated hairs. The Journal of Clinical Endocrinology and Metabolism, 39(6), 1012–1019. [DOI] [PubMed] [Google Scholar]
- Sharpley CF, McFarlane JR, & Slominski A (2011). Stress-linked cortisol concentrations in hair: what we know and what we need to know. Reviews in the Neurosciences, 23(1), 111–121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sheth MJ (2019). Grappling with racism as foundational practice of science teaching. Science Education, 103(1), 37–60. [Google Scholar]
- Skinner ML, Shirtcliff EA, Haggerty KP, Coe CL, & Catalano RF (2011). Allostasis model facilitates understanding race differences in the diurnal cortisol rhythm. Development and Psychopathology, 23(4), 1167–1186. [DOI] [PMC free article] [PubMed] [Google Scholar]
- SpitLab H.A.I.R. Project. (n.d.). Vimeo. [Google Scholar]
- Stalnaker- Shofner DM (2020). Colorism and Racial Identity Development in Black/African American Women: An Autoethnographic Perspective. Journal of Black Sexuality and Relationships, 7(2), 75–98. [Google Scholar]
- Sukmawati SS, & Pramudya Y (2019). Light Intensity Measurement On Sun-Earth System Model Using Logger Pro Light Sensor. Journal of Physics. Conference Series, 1254(1), 012067. [Google Scholar]
- Teran-Garcia M, Hammons A, Greder K, Olvera N, Matos NJC, Plaza-Delestre M, Wiley A, & Fiese BH (2019). NP9 “Abriendo Caminos”: Opening Opportunities for Healthy Pathways. Journal of Nutrition Education and Behavior, 51(7, Supplement), S13–S14. [Google Scholar]
- Tsigos C, & Chrousos GP (1994). Physiology of the hypothalamic-pituitary-adrenal axis in health and dysregulation in psychiatric and autoimmune disorders. Endocrinology and Metabolism Clinics of North America, 23(3), 451–466. [PubMed] [Google Scholar]
- UCL. (2016, October 24). Eugen fischer’s hair colour gauge. UCL CULTURE. https://www.ucl.ac.uk/culture/ucl-science-collections/eugen-fischers-hair-colour-gauge [Google Scholar]
- Walker A, Wiltz T, & Winfrey O (1997). Andre Talks Hair! Simon & Schuster. [Google Scholar]
- Wang W, Moody SN, Kiesner J, Appiani AT, Robertson OC, & Shirtcliff EA (2018). Assay Validation of Hair Androgens Across the Menstrual Cycle. Psychoneuroendocrinology. 10.1016/j.psyneuen.2018.10.029 [DOI] [PubMed] [Google Scholar]
- Wang W, van Dammen L, Moody S, Neiderhiser JM, Dismukes A, Constantinidis C, & Shirtcliff E (2020). Assessment of progesterone concentration in hair: Validation of an immuno-assay protocol in human and monkey hair samples. Psychoneuroendocrinology, 119, 105036. [Google Scholar]
- Wang W, van Dammen L, Moody SN, Kiesner J, Neiderhiser JM, Dismukes A, Zhu J, Constantinidis C, & Shirtcliff EA (2020). The validation of estradiol extraction and analysis from hair. 10.31234/osf.io/knuxs [DOI]
- Webb EC, White CD, Van Uum S, & Longstaffe FJ (2015). Integrating cortisol and isotopic analyses of archeological hair: reconstructing individual experiences of health and stress. American Journal of Physical Anthropology, 156(4), 577–594. [DOI] [PubMed] [Google Scholar]
- Wester VL, van der Wulp NRP, Koper JW, de Rijke YB, & van Rossum EFC (2016). Hair cortisol and cortisone are decreased by natural sunlight. Psychoneuroendocrinology, 72, 94–96. [DOI] [PubMed] [Google Scholar]
- Whitesel AL, Dismukes A, White D, Guyer S, Ganiban JM, Leve LD, & Neiderhiser JM (2020). The Early Growth and Development Study: Using an adoption design to understand family influences and child development. In The Routledge Handbook Of Adoption (pp. 90–104). Routledge. [Google Scholar]
- White SF, Lee Y, Phan JM, Moody SN, & Shirtcliff EA (2019). Putting the flight in “Fight-or-Flight”: Testosterone reactivity to skydiving is modulated by autonomic activation. Psychoneuroendocrinology, 100, S8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wilkerson I (2020). Caste (Oprah’s Book Club): The Origins of Our Discontents. Random House Publishing Group. [Google Scholar]
- Wilson AS, Dodson HI, Janaway RC, Pollard AM, & Tobin DJ (2007). Selective biodegradation in hair shafts derived from archaeological, forensic and experimental contexts. The British Journal of Dermatology, 157(3), 450–457. [DOI] [PubMed] [Google Scholar]
- Wosu AC, Gelaye B, Valdimarsdóttir U, Kirschbaum C, Stalder T, Shields AE, & Williams MA (2015). Hair cortisol in relation to sociodemographic and lifestyle characteristics in a multiethnic US sample. Annals of Epidemiology, 25(2), 90–95, 95.e1–e2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wright KD, Ford JL, Perazzo J, Jones LM, Mahari S, Sullenbarger BA, & Laudenslager ML (2018). Collecting Hair Samples for Hair Cortisol Analysis in African Americans. Journal of Visualized Experiments: JoVE, 136. 10.3791/57288 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yamanashi Y, Teramoto M, Morimura N, Hirata S, Suzuki J, Hayashi M, Kinoshita K, Murayama M, & Idani G ‘ichi. (2016). Analysis of hair cortisol levels in captive chimpanzees: Effect of various methods on cortisol stability and variability. MethodsX, 3, 110–117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang HZ, Lan J, Meng YJ, Wan XJ, & Han DW (1998). A preliminary study of steroid reproductive hormones in human hair. The Journal of Steroid Biochemistry and Molecular Biology, 67(5–6), 447–450. [DOI] [PubMed] [Google Scholar]
- Zimmer C, Basler H-D, Vedder H, & Lautenbacher S (2003). Sex differences in cortisol response to noxious stress. The Clinical Journal of Pain, 19(4), 233–239. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.