

Abstract
Following a request from the European Commission, the EFSA Panel on Plant Health performed a quantitative risk assessment of Leucinodes orbonalis (Lepidoptera: Crambidae), the eggplant fruit and shoot borer, for the EU. The assessment focused on potential pathways for entry, climatic conditions favouring establishment, spread and impact. Options for risk reduction are discussed but effectiveness was not quantified. L. orbonalis is a key pest of eggplant (aubergine/brinjal) in the Indian subcontinent and occurs throughout most of southern Asia with records mostly from India and Bangladesh. The main pathway of entry is fruit of solanaceous plants, primarily exotic varieties of eggplant, Solanum melongena and turkey berry, S. torvum. The trade in both commodities from Asia is small but nevertheless dwarfs the trade in other Solanum fruits from Asia (S. aethiopicum, S. anguivi, S. virginianum, S. aculeatissimum, S. undatum). Other Solanum fruits were therefore not further assessed as potential pathways. The trade in eggplant from Asia consists of special fruit types and caters mostly to niche markets in the EU, while most eggplant consumed in Europe is produced in southern European and northern African countries, where L. orbonalis does not occur. Using expert knowledge elicitation (EKE) and pathway modelling, the Panel estimated that approximately 3–670 infested fruit (90% certainty range, CR) of S. melongena or fruit bunches of S. torvum enter into regions of the EU that are suitable for L. orbonalis establishment each year. Based on CLIMEX modelling, and using two possible thresholds of ecoclimatic index (EI) to indicate uncertainty in establishment potential, climates favouring establishment occur mostly in southern Europe, where, based on human population, approximately 14% of the imported produce is distributed across NUTS2 regions where EI ≥ 30; or 23% of the produce is distributed where EI ≥ 15. Escape of adult moths occurs mostly from consumer waste. By analysing results of different scenarios for the proportion of S. melongena and S. torvum in the trade, and considering uncertainties in the climatic suitability of southern Europe, adult moth emergence in areas suitable for establishment is expected to vary between 84 individuals per year and one individual per 40 years (based on 90% CR in different scenarios). In the baseline scenario, 25% of the solanaceous fruit from Asia is S. torvum, 75% is S. melongena and EI ≥ 30 is required for establishment. After accounting for the chances of mating, host finding and establishment, the probability of a mated female establishing a founder population in the EU is less than 1 in 100,000 to about 1 event per 622 years (90% CR in baseline scenario). The waiting time until the first establishment is then 622 to more than 100,000 years (CR). If such a founder population were established, the moth is estimated to spread at a rate of 0.65–7.0 km per year after a lag phase of 5–92 years. The impact of the insect on the production of eggplant is estimated to be 0.67%–13% (CR) if growers take no specific action against the insect and 0.13%–1.9% if they do take targeted actions. Tomato (S. lycopersicum) and potato (S. tuberosum) are hosts of L. orbonalis, but the insect does not develop to maturity in tomato fruit, and it does not feed on potato tubers under field conditions; hence, damage to potato can only occur due to feeding on shoots. Tomato and potato are not preferred hosts; nevertheless, impact can occur if populations of L. orbonalis are high and preferred hosts are not available. The Panel did not assess this damage due to insufficient information.
Keywords: @risk, eggplant fruit and shoot borer, expert knowledge elicitation (EKE), impact, meta‐analysis, pathway, quantitative PRA
SUMMARY
Following a request from the European Commission, the EFSA Panel on Plant Health performed a quantitative risk assessment of Leucinodes orbonalis (Lepidoptera: Crambidae), the eggplant fruit and shoot borer, for the EU. The assessment focused on potential pathways for entry, climatic conditions favouring establishment, spread and impact. Options for risk reduction are discussed but effectiveness was not quantified.
Leucinodes orbonalis is a key pest of eggplant (brinjal) in the Indian subcontinent and occurs throughout most of southern Asia with records mostly from India and Bangladesh. The main pathway of entry is fruit of Solanaceous plants, primarily exotic varieties of eggplant, Solanum melongena, turkey berry, S. torvum, but also including other Solanum species (S. aethiopicum, S. anguivi, S. virginianum, S. aculeatissimum, S. undatum). Interceptions have been reported on all these species, but S. melongena and S. torvum are the species with the greatest number of interceptions.
The Panel assessed information in the literature on hosts of L. orbonalis concluding that only species in the genus Solanum provide a pathway as the fruits of these species can be imported under current regulation, while viable larvae and pupae may be transported with the fruit of most species, though not with tomato (S. lycopersicum) as the fruit is too wet for the larvae to develop in. Also, the damage is restricted to species in the genus Solanum.
The trade in eggplant from Asia consists of special fruit types and caters mostly to niche markets in the EU. S. torvum is imported as bunches of small fruit that are used as a spice in exotic dishes. The trade in both commodities from Asia is small but nevertheless dwarfs the trade‐in other Solanum fruit from Asia. These other Solanum fruits were therefore not further assessed as a pathway. The pathway most likely to provide a route for entry of L. orbonalis into the EU was judged to be fresh eggplant and fresh turkey berry from Asia.
Using expert knowledge elicitation (EKE) and pathway modelling, the Panel estimated that in the order of hundreds of thousands of fruit enter the EU each year. In the model, these fruits are distributed across the EU according to population, as the niche markets receiving these products are assumed to be homogeneously distributed across populations in the EU. NUTS regions where climatic conditions are conducive for establishment of L. orbonalis (median estimate with EI ≥ 15) receive approximately 427,000 transfer units; 90% CR approximately 237,000–715,000. With an EI threshold of 30, the number of fruits entering NUTS2 regions where parts are suitable for establishment drops to approximately 260,000 (90% CR approximately 144,000–436,000).
Infested fruit represent a small proportion of the total number of fruits entering the EU (in the order of one in a 2000 fruit are infested). The number of transfer units infested with live L. orbonalis entering NUTS2 areas with EI ≥ 15 is estimated to be approximately 175 per year (90% CR approximately 6–1100); using an EI threshold of 30, the median number of infested transfer units drops to approximately 105 per year (90% CR approximately 3–670). In the scenario where only turkey berry is imported, and using EI ≥ 15, the 95 percentile estimates 84 adults emerge in areas suitable for establishment. In contrast, where only eggplant is imported and using EI ≥ 30, the 5 percentile estimates one adult emerging in 40 years.
Climatic conditions are most suitable for establishment in parts of the southern EU, especially in Spain (Andalucía, Comunidad Valenciana and Extremadura), Portugal (Alentejo), Italy (Sicilia, Calabria, Puglia), Greece (Kriti) and Cyprus. When imports are allocated in proportion to the human population, between 14% and 23% of transfer units enter regions of the EU suitable for establishment (lower estimates based on EI ≥ 30, higher estimate based on EI ≥ 15). Of the infested units entering NUTS regions where EI ≥ 15 approximately 12% are discarded before reaching the final consumer and approximately 50% of infested units are discarded by the consumer (Appendix D). Further, 1.0% (median; 90% CR, 0.2%–1.9%) of larvae survive to adulthood and escape from commercial waste while a median of 5.2% (90% CR 0.98%–12.2%) escape from consumer household waste. When the resulting numbers of adults emerge across NUTS2 regions, the likelihood that a female will find a mate depends on the window of encounter in space and time. In combination with the likelihood that the subsequent progeny survives to initiate a founder population, the number of established founder populations was estimated to be 0.00014 per year (90% CR 0.00000–0.00264). Thus, the Panel would not expect new founder populations within the foreseeable future or the time horizon of 5 years of this assessment. The predicted waiting time between new founder populations is in the Panel's estimation at least approximately 380 years. Given such low estimates, the Panel did not proceed to quantitatively assess the effectiveness of risk reduction options targeting L. orbonalis. However, such options exist, for example, the production of eggplant and turkey berry fruit in pest‐free places of production and designation of L. orbonalis as a quarantine species. The Panel found four notifications in Europhyt (in 2011 and 2012) mentioning that product infested with L. orbonalis was granted entry to the EU. The temporary emergency measures for L. orbonalis that were instated in October 2022 (Commission Implementing Regulation (EU) 2022/1941 of 13 October 2022) will have stopped this practice.
If L. orbonalis would be introduced into the EU, the Panel estimates that it would take between 5 and 92 years (90% CR; median 34.5 years) for populations to grow sufficiently before a steady rate of spread of approximately 2.3 km/year (90% CR 0.65–7.02 km/year) was reached.
In a scenario where L. orbonalis enters, establishes and spreads within the EU and the population reaches an approximate equilibrium such that EU farmers consider the organism a member of the general pest fauna, median eggplant yield losses are estimated to be 4.5% (90% CR 0.67%–13.0%) when no specific control measures are in place, and 0.54% (90% CR 0.13%–1.94%) when growers apply targeted pest control against L. orbonalis.
The Panel did not assess the potential of damage to potato and tomato, major hosts of L. orbonalis that are widely grown in the potential area of establishment. There is sparse information in the literature on damage to these two crops, even though they are widely grown in countries where L. orbonalis is a serious pest, particularly India and Bangladesh. This suggests that the damage is unimportant, though there are few papers that state the contrary. Potato and tomato are known to be incidental hosts of L. orbonalis, accepted in case insects cannot find their favoured hosts, in particular S. melongena. However, with proper control of L. orbonalis in its main host, important spillovers to potato and tomato are not expected. Based on the scant information available, the Panel judges there to be insufficient evidence to regard L. orbonalis as a threat to the production of potato and tomato in the EU.
This PRA on L. orbonalis has several uncertainties as the Panel was unable to find information on (i) specific trade data on the commodities that are a pathway for L. orbonalis, (ii) information on consignment sizes and inspection practices for all the EU countries importing S. melongena and S. torvum, (iii) production practices in the countries of origin for eggplant or turkey berry destined for the European market and (iv) specific data demonstrating the potential for damage to potato and tomato.
In conclusion, L. orbonalis arrives with current measures in the EU with produce from Asian countries exporting eggplant and turkey berry to the EU. The numbers of insects entering are so low that establishment is a very rare event and unlikely in the foreseeable future. Were the insect to establish, it would spread and after it would reach an equilibrium population in the potential area of establishment, which includes a major part of the production area of eggplant in the EU, it would cause damage and add to the pest complex in this crop. Measures are available to reduce the likelihood of entry and consequently establishment, spread and impact.
1. INTRODUCTION
1.1. Background and terms of reference as provided by the requestor
1.1.1. Background
The new Plant Health Regulation (EU) 2016/2031, on the protective measures against pests of plants, is applying from 14 December 2019. Conditions are laid down in this legislation in order for pests to qualify for listing as Union quarantine pests, protected zone quarantine pests or Union regulated non‐quarantine pests. The lists of the EU regulated pests together with the associated import or internal movement requirements of commodities are included in Commission Implementing Regulation (EU) 2019/2072. Additionally, as stipulated in the Commission Implementing Regulation 2018/2019, certain commodities are provisionally prohibited to enter in the EU (high‐risk plants, HRP). EFSA is performing the risk assessment of the dossiers submitted by exporting to the EU countries of the HRP commodities, as stipulated in Commission Implementing Regulation 2018/2018. Furthermore, EFSA has evaluated a number of requests from exporting to the EU countries for derogations from specific EU import requirements.
In line with the principles of the new plant health law, the European Commission with the Member States are discussing monthly the reports of the interceptions and the outbreaks of pests notified by the Member States. Notifications of an imminent danger from pests that may fulfil the conditions for inclusion in the list of the Union quarantine pest are included. Furthermore, EFSA has been performing horizon scanning of media and literature.
As a follow‐up of the above‐mentioned activities (reporting of interceptions and outbreaks, HRP, derogation requests and horizon scanning), a number of pests of concern have been identified. EFSA is requested to provide scientific opinions for these pests, in view of their potential inclusion in the lists of Commission Implementing Regulation (EU) 2019/2072 and the inclusion of specific import requirements for relevant host commodities, when deemed necessary.
1.1.2. Terms of reference (ToR)
EFSA is requested, pursuant to Article 29(1) of Regulation (EC) No 178/2002, to provide scientific opinions in the field of plant health.
EFSA is requested to deliver 50 pest categorisations for the pests listed in Annex 1A, 1B and 1D. Additionally, EFSA is requested to perform pest categorisations for the pests so far not regulated in the EU, identified as pests potentially associated with a commodity in the commodity risk assessments of the HRP dossiers (Annex 1C). Such pest categorisations are needed in the case where there are not available risk assessments for the EU.
When the pests of Annex 1A are qualifying as potential Union quarantine pests, EFSA should proceed to phase 2 risk assessment. The opinions should address entry pathways, spread, establishment, impact and include a risk reduction options analysis.
Additionally, EFSA is requested to develop further the quantitative methodology currently followed for risk assessment, in order to have the possibility to deliver an express risk assessment methodology. Such methodological development should take into account the EFSA Plant Health Panel Guidance on quantitative pest risk assessment and the experience obtained during its implementation for the Union candidate priority pests and for the likelihood of pest freedom at entry for the commodity risk assessment of high‐risk plants.
Annex 1. List of pests.
A)
1. Amyelois transitella
2. Citripestis sagittiferella
3. Colletotrichum fructicola
4. Elasmopalpus lignosellus
5. Phlyctinus callosus
6. Resseliella citrifrugis
7. Retithrips syriacus
8. Xylella taiwanensis
E)
List of pests identified to develop further the quantitative risk assessment (phase 1 and phase 2) methodology followed for plant pests, to include in the assessments the effect of climate change for plant pests (for more details, see Annex 3).
1. Leucinodes orbonalis
2. Leucinodes pseudorbonalis
3. Xanthomonas citri pv. viticola
1.2. Interpretation of the terms of reference
Leucinodes orbonalis is one of the three plant pest species listed in Annex 1E of the terms of reference. The pest categorisation of L. orbonalis concluded that the species satisfies the EU criteria that are within the remit of EFSA to assess, for it to be regarded as a potential Union quarantine pest (EFSA PLH Panel, 2021). Hence, EFSA is to proceed to conduct phase two of the risk assessment.
Adults of L. orbonalis are relatively small moths with predominantly white wings, featuring a characteristic triangular brown patch at half the forewings' length and a dark grey patch at the centre of the outer forewing margin (Figure 1); their wingspan is approximately 25 mm. The species was scientifically described based on specimens from Bangladesh and Java (Indonesia) and occurs widely in southern Asia (Figure 2), confirmed by DNA data (Chang et al., 2014; Sagarbarria et al., 2018; Shashank et al., 2015). These DNA data, however, indicate the existence of three still undescribed species belonging to the genus Leucinodes in the Austral‐Asian region: one in the northern part of Vietnam and two in the northern part of Australia (see Appendix A). Nonetheless, the Panel is almost certain that, in the available literature from Asia, L. orbonalis is correctly identified as this species.
FIGURE 1.

Leucinodes orbonalis adult female. © Donald Hobern – Flickr: Leucinodes orbonalis, CC BY 2.0, https://commons.wikimedia.org/w/index.php?curid=18001695
FIGURE 2.
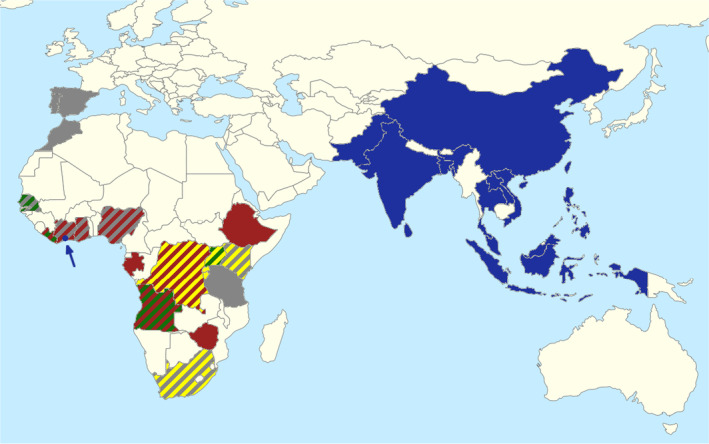
Genetically and morphologically confirmed country‐level occurrences of Leucinodes orbonalis (blue), and of the four African Leucinodes species intercepted in Europe, i.e. L. africensis (red), L. rimavallis (yellow), L. pseudorbonalis (green) and L. laisalis (grey). A single specimen of L. orbonalis has been found in Côte d'Ivoire (blue arrow and dot) outside its native Asian range. Map © Richard Mally.
Asia currently comprises seven species of moths placed in the genus Leucinodes: Leucinodes diaphana, L. labefactalis, L. melanopalis, L. orbonalis, L. perlucidalis, L. sigulalis and L. unilinealis (Nuss et al., 2003–2023). However, all species except L. orbonalis are misplaced in this genus and need to be transferred to different genera in Crambidae (R. Mally, pers. obs.). Adults of the misplaced species can be distinguished externally from L. orbonalis (R. Mally, pers. obs.), so that misidentification is unlikely. Host plants of the misplaced Asian Leucinodes species are unknown except for L. melanopalis, whose larvae are reported to feed on Ficus religiosa (Moraceae) and Anacardium occidentale (Anacardiaceae) (Robinson et al., 2023).
Historically, all African specimens morphologically appearing like L. orbonalis have been attributed to that species. However, Mally et al. (2015) did not find specimens of L. orbonalis in museum material originating from Sub‐Saharan Africa. It is therefore extremely unlikely that L. orbonalis is present in Africa or only locally present through potential unintentional introduction from Asia (Figure 2). Mally et al. (2015) further discovered that the African specimens externally looking like L. orbonalis are in fact a complex of previously undescribed species. Several of these newly discovered species cannot be distinguished from the Asian L. orbonalis based on external morphology of the larvae or adults, and dissection of the male genitalia or analysis of the ‘DNA Barcode’ sequence is necessary for species identification. None of the literature reporting ‘L. orbonalis’ from Africa mentions identification efforts of the investigated African specimens by means of genitalia dissection and/or DNA sequences, and their correct identification is therefore almost impossible; the Panel is currently almost certain that all African specimens identified as L. orbonalis in African literature are misidentifications of the species first described by Mally et al. (2015) or of still undiscovered species. The literature on African Leucinodes published since 2015 appears to be largely unaware of the African species complex described by Mally et al. (2015).
Four of the African Leucinodes species have been intercepted in Europe and identified to species based on male genitalia morphology and/or DNA ‘Barcode’ sequences: L. africensis, L. pseudorbonalis and L. rimavallis, which are externally indistinguishable from each other and from the Asian L. orbonalis, and the greyish‐brown L. laisalis, which was reported as Sceliodes laisalis or Daraba laisalis in earlier literature. The latter species has established in the south of Spain and Portugal, with observations from 1958 to 2023 (see Appendix E, Spread). The Asian L. orbonalis has mainly been intercepted from eggplant/brinjal (Solanum melongena), whereas the African Leucinodes species have mainly been intercepted from bitter tomato (S. aethiopicum).
The Leucinodes (Lepidoptera: Crambidae) taxonomic specialist (Dr Richard Mally) is almost certain that Leucinodes orbonalis from Asia is correctly identified as such, and that the Asian literature on L. orbonalis can reliably be attributed to this species. It is extremely likely that African Leucinodes species looking like (and identified as) L. orbonalis are misidentifications of L. pseudorbonalis, L. africensis, L. rimavallis and L. kenyensis (all described as new in Mally et al., 2015, EFSA PLH Panel, 2021) or of still undescribed species (see ‘Leucinodes spp.’ in Mally et al., 2015); in the larval stage, L. orbonalis can furthermore be confused with the African L. laisalis. It is impossible to determine in retrospect which of the four known African species resembling L. orbonalis were studied in African literature; consequently, host plants and information on the biology reported in African literature cannot be reliably attributed to any of these four African species (unless voucher specimens were kept from the studies, which, however, none of the African studies indicated). Therefore, host plant records for L. orbonalis from countries outside of Asia might be incorrect, and in the case of African records may refer to any of the four known African look‐alike species.
Given this situation, the request to conduct a quantitative pest risk assessment on L. orbonalis has been interpreted as a request to assess the risk from L. orbonalis using the very comprehensive literature from Asian populations. Literature from Africa purporting to concern L. orbonalis will not be considered but will be assessed in a separate opinion. Entry pathways in the current opinion will exclude Africa as a possible source of L. orbonalis as there is a lack of reliable evidence that the species occurs in Africa.
Taxonomic experts know of a single specimen of L. orbonalis, identified based on the male genitalia, that was intercepted on Solanum sp. imported in 2011 from Cote d'Ivoire to France (J.‐M. Ramel, personal communication). The Panel considers the possibility that L. orbonalis may have been unintentionally introduced to Cote d'Ivoire through fruit imports from Asia, and that it may have established a founder population there. At the moment, however, this is speculation and requires further investigation.
Entry, establishment, spread and impact are to be quantitatively evaluated. An analysis of risk reduction options is also required. The Panel will therefore undertake a quantitative pest risk assessment according to the principles laid down in its guidance on quantitative pest risk assessment (EFSA PLH Panel, 2018) while recognising the need of the Commission for an express (i.e. as fast as possible) risk assessment.
In addition, as agreed with the Commission, the effect of climate change will not be examined for L. orbonalis as it has instead been assessed previously for Elasmopalpus lignosellus Zeller (Lepidoptera: Pyralidae) (EFSA PLH Panel, 2023). A brief discussion on consequences of climate change is given in Section 3.6.
2. DATA AND METHODOLOGIES
To obtain a deeper understanding of the organism and to inform the necessary steps in the risk assessment, a literature review was conducted using the Web of Science databases. The review built on the information collected for the pest categorisation (EFSA PLH Panel, 2021). The scientific and common names of the pest were used as search terms, no filters (limits) for either time of publication nor language were implemented and all Web of Science databases were selected. The following search string was used to retrieve results: Leucinodes orbonalis OR Leucinodes pseudorbonalis.
The Web of Science search resulted in 1293 hits after removal of duplicates. An additional search was conducted via the Google Scholar search engine to specifically find literature published in French, with the following French names inserted individually (with number of results in parentheses): foreuse des solanacées (8), perceuse de l'aubergine (2). The Web of Science search was conducted on 23 March 2021, and the Google Scholar search in August 2023. Of the altogether 2164 references found to mention Leucinodes, full texts of 583 references could not be retrieved.
Additional searches to retrieve additional specific documents cited in other literature were run when developing the opinion. The available scientific information, including the previous EFSA pest categorisation (EFSA PLH Panel, 2021) and the relevant literature and legislation, e.g. Regulation (EU) 2016/2031, Commission Implementing Regulation (EU) 2019/2072 and Commission Implementing Regulation (EU) 2022/1941 were taken into account.
In performing the risk assessment, the following assessment steps were distinguished after identifying appropriate pathways:
Estimating the number of infested host fruit that enter the EU,
Identifying the areas where L. orbonalis can establish in the EU,
Quantifying the number of host fruit entering NUTS2 areas of the EU where climatic conditions are suitable for establishment and where the pest could reproduce resulting in transfer to a host in those areas, leading to the initiation of a founder population,
Estimating the duration of the lag period before a founder population begins to spread as well as the steady rate of spread,
Estimating the potential loss in yield of solanaceous host crops in situations with and without specific pest management of L. orbonalis being used by farmers.
Judgements made in each assessment step were based on a combination of literature review, meta‐analysis, information collected during interviews with hearing experts and expert knowledge elicitation (EKE) involving Panel members and EFSA staff to assess quantities that could not be well identified from the literature or databases alone (EFSA, 2014). To link commodity entry volumes into the EU with the assessment of establishment, imported commodities were distributed by apportioning relevant imported plant products to NUTS2 regions on the basis of the human population in each NUTS2 region, on the assumption that consumer demand is proportional to population size. Human population data were sourced from Eurostat (EFSA PLH Panel, 2018).
According to ISPM 5 (FAO, 2018), entry is ‘movement of a pest into an area where it is not yet present, or present but not widely distributed and being officially controlled’ while establishment is ‘perpetuation, for the foreseeable future, of a pest within an area after entry’. Introduction, according to the same ISPM 5, is ‘the entry of a pest resulting in its establishment’. In the assessment of entry, the Panel first identified pathways for entry of L. orbonalis into Europe, finding the main pathways to be Solanum produce, specifically exotic/special cultivars of eggplant, S. melongena and turkey berry, S. torvum. The volume of imports into the EU was estimated based on past imports, as well as the proportion of host fruit infested (Section 2.1). A pathway model was developed. Attention then shifted from pathway modelling of entry to identifying and mapping areas of the EU where establishment is possible following entry. Methods are described in detail in Rossi et al. (2023) – available on the Zenodo platform – and summarised in Sections 2.2.1, 2.2.5. After identification of the areas at risk using CLIMEX, SDM and Köppen–Geiger climate mapping, the pathway modelling was continued in Section 2.2.6. In this section, the entry flow is partitioned to parts of Europe suitable for establishment and not suitable for establishment. Transfer is modelled using a stochastic pathway model only for the areas where establishment is likely, assuming that no populations of L. orbonalis will be founded in areas that are not suitable for its establishment. Section 2.1 presents the overall pathway model for introduction, encompassing both entry and establishment.
2.1. Entry
2.1.1. Identifying pathways
Leucinodes orbonalis is an oligophagous pest that feeds on different plant species in the nightshade family (Solanaceae), with eggplant (Solanum melongena) being by far the most important and impacted plant species (EFSA PLH Panel, 2021). The larvae bore into the stems and fruits, weakening the host plant and rendering the fruits unfit for sale (Appendices B and F). The Panel compiled a list of host plants that are imported into the EU that could plausibly act as vehicles for entry (e.g. Table C.1 in Appendix C). Entry would require the importation of fruits or stems with eggs or pupae attached to the outside or with larvae feeding in or on the fruit. Stems are not a pathway because they are not traded as a commodity. Furthermore, it is not allowed to introduce plants for planting of Solanum or tubers of Solanum spp. into the EU from countries where L. orbonalis is present (EU regulation 2019/2072 Annex VI parts 15–18). EU regulation 2019/2072 Annex VI part 18 bans imports of plants for planting of Solanaceae. Imports of tubers of potato, S. tuberosum and other tuber forming species of Solanum are regulated by EU regulation 2019/2072 Annex IV, parts 15, 16 and 17. No such imports are allowed from countries in which L. orbonalis is present. Hence, efforts to identify plausible pathways focussed on (i) commodities on which interceptions had been found and (ii) hosts that are imported into the EU as fruits from countries (Bangladesh, Brunei Darussalam, China, Indonesia, India, Japan, Cambodia, Lao People's Democratic Republic, Sri Lanka, Myanmar‐Burma, Malaysia, Nepal, Philippines, Pakistan, Singapore, Thailand, Taiwan, Vietnam) where L. orbonalis is known to occur.
Saudi Arabia, the United Arab Emirates and Australia were excluded from the list of countries where L. orbonalis is considered present. In Australia, L. orbonalis appears to be misidentified and likely represents one or two still undescribed species of Leucinodes (Appendix A). Leucinodes in the two Arabian countries are potentially one or several of the African species described in Mally et al. (2015).
Except for an isolated population in northern Vietnam that is genetically distinct from L. orbonalis but has to this date not been described as a separate species (Srinivasan Ramasamy and R. Mally, personal communication), all Leucinodes from Asian countries feeding on solanaceous species are considered to be L. orbonalis. The Asian species of Leucinodes that do not feed on solanaceous species are distinct from Leucinodes are incorrectly placed in this genus (R. Mally, pers. obs.); however, that does not affect this opinion as these species are not reported to feed on Solanum spp.
Interceptions: EU data of interceptions are shown in the pest categorisation (EFSA PLH Panel, 2021). However, some EU member states only make Europhyt notifications for interceptions of quarantine pests and L. orbonalis was listed as a quarantine pest in October 2022. Future trade flow of goods on which interceptions were found in the EU was estimated from Eurostat data. Eurostat aggregates some types of vegetable commodity imports and the accessible eight‐digit codes do not specifically identify S. torvum. Nevertheless, the WG was able to identify and focus on the pathways most likely to lead to pest entry after excluding hosts whose import practice was judged unlikely to provide a pathway.
2.1.2. Scenario definitions for entry
An evidence dossier to support judgements of entry was developed based on literature review. The collected evidence is summarised in Appendix C: Entry was reviewed during the EKE to develop a pathway model for entry.
Estimates of the probability of units of the imported commodity being infested with L. orbonalis were made and uncertainties identified using expert judgement following EFSA guidance (Annex B.8 of EFSA Scientific Committee, 2018).
Scenario description: considering existing practices and phytosanitary measures
To estimate the number of host commodity units entering the EU infested with the pest, the Panel developed a general scenario with the following description:
The vegetable fruit of eggplant (S. melongena) and turkey berry (S. torvum) are considered the only significant possible pathway for introduction of L. orbonalis.
Most of the eggplant (S. melongena) consumed in the EU is produced and traded within the EU, especially in Spain and Italy.
Special and exotic varieties of eggplant (S. melongena) and turkey berry (S. torvum) are imported from Asian countries over the next few years in similar volumes and frequency to imports during the period 2010–2019. Data from 2020 to 2022 were not considered due to potential trade disruption during the Covid‐19 pandemic. Data were sourced from Eurostat.
The proportion of infested fruit is based on information on production practices in countries of origin, literature on impact in countries of origin and the frequency of interceptions in the past.
In tropical and subtropical regions of Asia, L. orbonalis is reproducing year‐round, with the potential of 10 and more overlapping generations per year (Appendix B: Biology).
Production and pest management: Eggplant is grown primarily in the open field, very rarely in protected conditions (greenhouses); in India, there is heavy use of pesticides (personal communication dr S. Ramasamy, A. Jovanovic); population control furthermore uses pheromone mass trapping, but light traps are avoided as these also attract other pests (personal communication Dr S. Ramasamy).
Post‐harvest management: For local markets, post‐harvest treatment is minimal, and fruits are sold within a few hours to days. Fruits determined for export: grading of fruits according to colour and size, storage for 1–4 days in shade at ambient temperature, or 7–10 days at 7–10°C and 85%–95% relative humidity (National Horticulture Board, 2023); sorting is done meticulously by farmers to avoid rejection at market (personal communication Dr S. Ramasamy).
Transport to EU: In containers via airplane, mostly in small quantities (too small to further split up before distribution in the EU) and in mixed consignments; purchasers are mostly restaurants and ethnic food shops (personal communication A. Jovanovic).
An estimate of imports of eggplant and turkey berry was determined from previous trade data. Unfortunately, Eurostat HS codes (8‐digit resolution) do not discriminate between eggplants and turkey berry and they are combined within code HS 0709 3000. As such the PLH Panel cannot estimate the amount of each commodity imported individually. Instead, three simple scenarios were imagined:
Scenario 1: 75% of imports are eggplants, 25% turkey berry. This percentage split is based on number of interceptions on eggplants (222) and number of interceptions on turkey berry (66).
Scenario 2: 100% of imports are turkey berry, 0% are eggplants. While this is an unrealistic scenario it gives the highest likelihood of entry, representing a worst‐case scenario.
Scenario 3: 0% of imports are turkey berry, 100% are eggplants. Again, this is an unrealistic scenario but allows the lower limit of entry to be determined.
In reality, the true proportions of imports are somewhere between Scenario 2 and 3. Scenario 1 was felt reasonable.
EKE was applied to answer a sequence of questions according to the pathway model of Figure 3:
What is the mean weight of each imported commodity (single S. melongena fruit or bunch (cluster) of S. torvum) (kg)? Estimates are then used to provide an estimate of the range of transfer units imported.
What is the proportion of infested transfer units entering the EU? The risk assessment used individual host fruit of S. melongena or bunches (clusters) of S. torvum as the most suitable unit for transfer unit because data are available on sampling procedures used for inspection at entry in the EU. The sampling protocols use eggplant (S. melongena) fruit as a unit of sampling.
FIGURE 3.
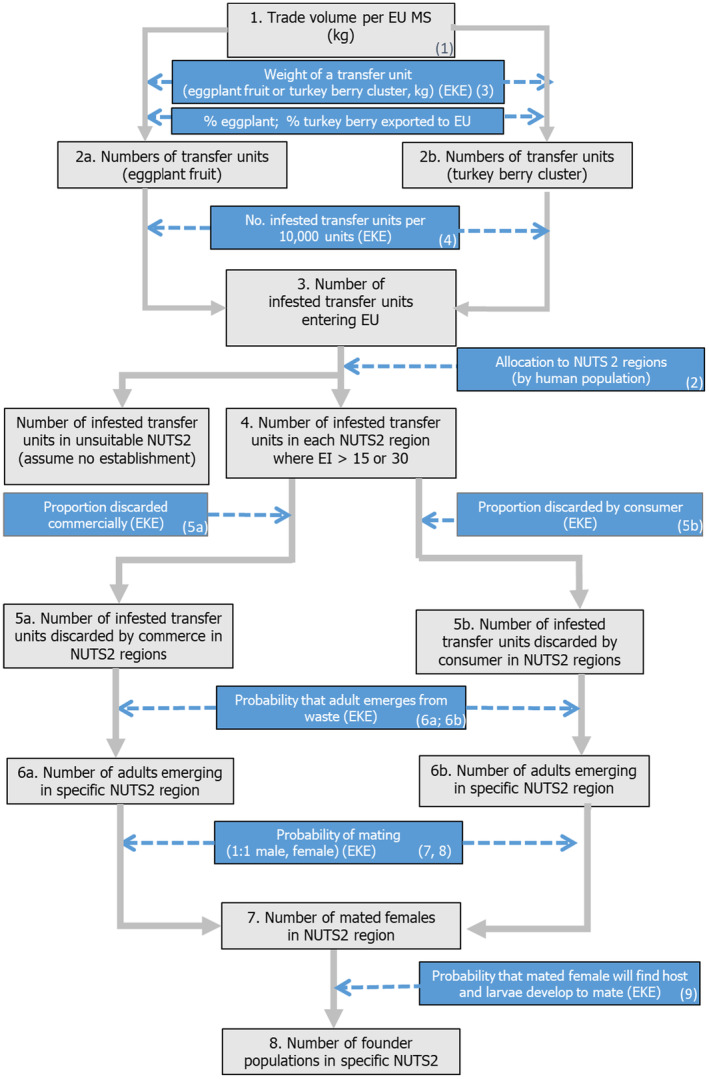
Conceptual diagram of pathway model to quantitatively estimate the likelihood of introduction of L. orbonalis into the EU. Blue is a parameter and grey is a variable (Numbers in brackets correspond to numbering of parameters in spreadsheet model (see Supporting materials – Annex A)). A mathematical description of the pathway model is given in Appendix C section Analysis and Appendix I. The Excel implementation of the pathway model, with a user‐friendly presentation of the parameters and intermediate results of the calculation, is available in the supplementary materials to this opinion.
The uncertainties associated with the EKE were taken into account and quantified in the probability distribution applying the semi‐formal method described in Section 3.5.2 of the EFSA‐PLH Guidance on quantitative pest risk assessment (EFSA PLH Panel, 2018).
When results from the modelling of introduction were obtained which showed the very small likelihood of introduction in the foreseeable future for any of the three scenarios (import is composed of 100% S. melongena, 100% S. torvum or 75% S. melongena and 25% S. torvum), no specific scenarios for risk‐reduction options were quantitatively evaluated. Available options are briefly mentioned (Sections 3.2.8 and 3.5).
2.2. Establishment
A detailed description of the methods used to assess the area in the EU where climatic conditions could support establishment of L. orbonalis is provided in Rossi et al. (2023). The assessment of establishment considered outdoor conditions only. Four methods were used to inform assessment of the area of potential establishment. (1) Mapping of Köppen–Geiger zones in Europe and in the area where the organism is distributed to evaluate whether the insect is present in climate types that are also present in the EU, (2) mapping of accumulated degree days and derived number of generations for Europe and Asia to evaluate whether the insect is present in Asia in areas with cumulated degree days that occur in Europe, (3) the CLIMEX modelling system (Kriticos et al., 2015), to analyse the potential for growth and persistence (i.e. establishment) of the insect and compare model output with known occurrence in Asia and then use the same parameters to show where growth and establishment could occur in the Europe‐Mediterranean region and (4) species distribution modelling (SDM) which uses correlative methods to assess correspondence between predictor variables (e.g. climate variables) and the distribution of the insect in Asia, allowing to derive a climate suitability index for Europe. Developing SDM for the case of L. orbonalis provides a potential valuable comparison of SDM to CLIMEX which will inform its usefulness for exploring areas at risk for African Leucinodes species.
2.2.1. Literature search on the distribution and ecophysiology of Leucinodes orbonalis
An extensive literature search for pest distribution was conducted in Web of Science (all databases, excluding Data Citation Index and Zoological Record) and Scopus on 14 September 2022 (Rossi et al., 2023). The search string was based only on the scientific and common names of the pest. Other keywords such as ‘biology’, ‘physiology’ and ‘temperature’ were not used, so as not to limit the retrieval of distribution data, often reported as secondary information. The review followed a two‐step approach for selecting relevant papers, the first step was based on screening the title and abstract of the paper, while the second step was based on the full‐text analysis. A full description of the literature search methodology is available in Rossi et al. (2023).
2.2.2. Köppen–Geiger climate classification analysis
The SCAN‐Clim tool (EFSA and Maiorano, 2022) was used to produce climate suitability maps based on the Köppen–Geiger climate classification approach. The re‐analysis of Rubel et al. (2017) of the Köppen–Geiger climate classification from Kottek et al. (2006), considering the period 1986–2010 (available at http://koeppen‐geiger.vu‐wien.ac.at/present.htm), was used. The climate types present in the observed locations of L. orbonalis were identified and mapped. Because the PRA area is the EU, the output maps considered only climate types that are also present in the EU (Rossi et al., 2023).
2.2.3. Degree days and number of generations maps
Degree days and number of generations maps were developed for South‐East Asia and Iran (area of distribution of L. orbonalis), and Europe and the Mediterranean Basin. Degree days were calculated by accumulating the positive daily differences between the daily mean temperature and a base temperature (BaseT) of 15°C. If daily mean temperature was below the base temperature, that day did not contribute to temperature accumulation. A linear regression model combining data from life‐table studies (Dhaliwal & Aggarwal, 2021; Islam et al., 2020) was used to estimate the number of degree days to complete one generation. Degree days were calculated using the Copernicus ERA5‐Land data (Muñoz Sabater et al., 2021) for the 30‐year period from 1993 to 2022.
2.2.4. Species distribution modelling
The suitability of the EU territory to the establishment of L. orbonalis was analysed using a species distribution model ensemble developed in rStudio (rStudio Team) using the R sdm package (Naimi and Araújo, 2016). The bioclimatic variables from WorldClim (Table 1), for the period 1970–2000, at the resolution of 10 arcmin (~ 18 km ix 18 km), were used as predictor variables (Fick & Hijmans, 2017) (https://www.worldclim.org/data/worldclim21.html).
TABLE 1.
Bioclimatic variables from WorldClim used in SDM (Fick & Hijmans, 2017).
| Name | Description |
|---|---|
| BIO2 | Mean Diurnal Range (Mean of monthly (max temp–min temp)) |
| BIO3 | Isothermality (BIO2/BIO7) (×100) |
| BIO8 | Mean Temperature of Wettest Quarter |
| BIO9 | Mean Temperature of Driest Quarter |
| BIO13 | Precipitation of Wettest Month |
| BIO15 | Precipitation Seasonality (Coefficient of Variation) |
| BIO17 | Precipitation of Driest Quarter |
| BIO18 | Precipitation of Warmest Quarter |
| BIO19 | Precipitation of Coldest Quarter |
Pest distribution data at the point level were used in the analysis as presence‐only data. These were thinned to include only one point, selected randomly in each grid cell with the same resolution of the predictor variables (10 arcmin or one sixth degree). The area for the training of the SDM model was between the geographical bounding‐box 40° E − 165° E and 10° S–45° N.
Pseudo‐absence data were generated inside the study area with two approaches. In the first approach, a modified convex hull polygon was created including an area that was assumed to be suitable for the organism based on known distribution. In the second approach, pseudo‐absences were created in the entire training area, with the only limitation of a buffer of 10 km around each distribution point.
For the convex‐hull approach, three series of simulations were run based on different number of pseudoabsence points: 4800 pseudo‐absence points (10x number of observations), 2400 (5x) and 480 (1x) random points. To avoid collinearity among predictors, the variance inflation factor (VIF) method was used to exclude all the Bioclimatic variables with collinearity. Then, the species distribution modelling methods were fitted to the predictor variables.
Ten models were used to fit the data: bioclim, brt, cart, domain.dismo, gam, mars, maxent, rf, rpart and svm. Data splitting was achieved through a fivefold cross‐validation process repeated for five times. Therefore, 25 simulations per model were created, yielding a total of 250 model runs. Ensemble modelling of the 250 simulations, based on the weighted average of the True Skill Statistics (TSS), was used to produce the final output.
2.2.5. CLIMEX
CLIMEX model (version 4.1.0.0, Kriticos et al., 2015) was used to investigate the climate suitability of the EU to the establishment of L. orbonalis. CLIMEX is based on the organism distribution and on its ecophysiological requirements to survive and complete the life cycle across a geographic region, given historic climate data. CLIMEX can assess the influence of weather‐related stress factors (cold, heat, drought, humidity), and their interactions, on survival and growth through the calculation of growth‐related indices and stress‐related indices. The two groups of indices are combined into an Ecoclimatic Index (EI), which quantifies suitability for establishment of the pest. Simulations were run using the climate data set CM30 1995H V2 WO (Kriticos et al., 2012), package v4.1 (available at: https://www.climond.org/). This data set is based on the 0.5° world grid of historical meteorological data (30 years centred on 1995) originating from the Climate Research Unit (Norwich, UK), and transformed using the methods of Kriticos et al. (2012). Rossi et al. (2023) give more detail on how the parameters used in CLIMEX were determined.
The ecoclimatic index EI spans the integers from 0 to 100, where 0 means that a place is unsuitable for the organism, whereas 100 means a place is highly suitable. It is expected that with increasing EI, the density and impact of an organism will increase. According to Kriticos et al. (2015), a value of EI greater than 30 demarcates areas where climate is (very) favourable for the species whereas areas where EI < 30 are less favourable. They state, ‘An EI of more than 30 represents a very favourable climate for a species, as it means that during the (say) six months suitable for growth with a maximum Growth Index (GI) of 50, the species has achieved 60% of the potential population growth’. However, a precise threshold value for establishment and impact cannot be given and any cut‐off value of EI may be species‐specific and should be operationally defined on the basis of additional evidence. The Panel used two EI thresholds (≥ 15 and ≥ 30) to identify areas where climate suitability favoured establishment. See also assessment Section 3.4 on Impact.
2.2.6. Transfer and initiation of a founder population
While most fresh eggplant and turkey berry fruit imported will be sold, cooked and consumed, a proportion is discarded at various steps along the supply chain by importers, wholesalers, retailers and the final consumers, e.g. due to damage during handling and transport, physical quality problems, market conditions and pest finds (Gould & Maldonado, 2006). There is a possibility that live larvae in discarded host fruits will develop to adulthood, escape from the discarded material and find a mate resulting in fertilised eggs being laid on a host plant in the neighbourhood of the discarded material, a process referred to as transfer. Should the subsequent progeny develop and reproduce, a potential founder population would have been initiated. The process of transfer and initiation of a founder population was broken down into four steps:
Estimating the proportion of imported host‐plant material discarded by commercial stakeholders in the supply chain due to e.g. infestation, physical damage, substandard quality or oversupply;
Estimating the proportion of infested material discarded by consumers;
The proportion of larvae that develop to adulthood and escape from discarded material;
The proportion of females that find a mating partner and find a suitable host plant in the surrounding environment and lay fertilised eggs;
The likelihood that adults develop from the eggs to reproduce and initiate a founder population.
Information to support judgements relating to these steps, necessary for establishment, was sought within the literature review. The collected evidence was reviewed during EKE and is summarised in Appendix D: Establishment.
2.2.7. Scenarios for establishment
A parameter in the model for pest introduction is the area of the EU where climatic conditions are suitable for L. orbonalis development. The area considered is within NUTS2 regions. Such information was determined using CLIMEX modelling (2.2.5). To capture uncertainty about the threshold for establishment, two thresholds were considered a lower Eco‐climatic Index (EI) threshold of 15, and a higher EI threshold of 30.
2.2.8. Overall model for introduction (entry and establishment)
The pathway model for introduction is a product of the following components:
Mean annual EU import quantity of potential transfer units (eggplant and turkey berry) from countries where L. orbonalis occurs;
Inverse weight of a single transfer unit (to calculate the number of imported fruits as the volume of trade (kg) divided by the weight of a single transfer unit);
Proportion of infested units entering the EU;
Proportion of infested units imported to suitable NUTS2 regions;
Proportion of infested units disposed of as waste;
Probability of larva in discarded unit surviving to become an adult;
Probability of a female mating;
Probability of a mated female initiating a founder population that persists.
Figure 3 illustrates the model for introduction.
With three scenarios for entry, based on the proportion of turkey berry and eggplant imported, and two scenarios for area of suitable establishment, based on different EI thresholds, six scenarios were considered for introduction, Table 2.
TABLE 2.
Key to introduction scenarios.
| % of turkey berry and eggplant imported | Threshold for Ecoclimatic Index | |
|---|---|---|
| EI 15 | EI 30 | |
| 25% turkey berry; 75% eggplant | Scenario 1 | Scenario 4 |
| 100% turkey berry; 0% eggplant | Scenario 2 | Scenario 5 |
| 0% turkey berry; 100% eggplant | Scenario 3 | Scenario 6 |
2.2.9. Distribution of imported infested eggplant and S. torvum in the EU
The Panel did not find information on the final destination of eggplant and S. torvum imported from Asian countries of origin. The Panel therefore developed the pathway model on the assumption that consumers of eggplant and S. torvum from Asia are equally represented across Europe. Thus, it was assumed that the imported product is apportioned to NUTS regions according to the population in each NUTS region.
2.2.10. Identifying NUTS2 regions with suitable climate for establishment
The fractions of CLIMEX grid cells in each NUTS2 region with EI ≥ 15, or EI ≥ 30, were determined. Each full grid cell approximates to 2500 km2. All grids and grid fractions with EI ≥ 15 or ≥ 30 were summed for each NUTS2 area and multiplied by 2500 to give an approximate area where climate may be suitable for establishment. NUTS2 areas were then ranked by suitable area.
2.3. Spread
The area of the colonised territory occupied during spread is expected to follow a sigmoid curve (Figure 4). After an initial lag phase of slow spread during which the founder population builds up, spread accelerates and reaches a constant rate for some time before declining again as the suitable area gets fully colonised (saturation phase). Rather than estimate the parameters for logistic spread (i.e. Figure 4), this assessment followed the method of EFSA PLH Panel (2018) to estimate the duration of the lag phase and the linear rate of range expansion when spread is at its fastest. In this way, the spread assessment is simplified.
FIGURE 4.
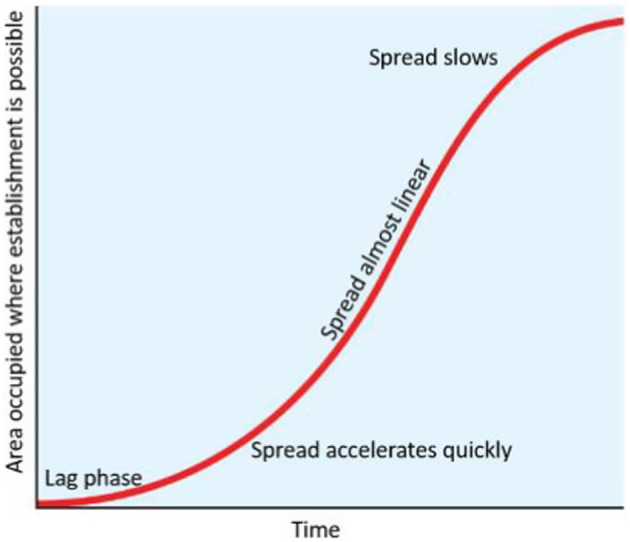
Stages of conceptual logistic spread: Following the lag phase (lag period) spread accelerates, becomes almost linear then slows.
Spread is not just expansion of a contiguous area, but it may also include the generation of distant satellite populations (Herms & McCullough, 2014; Muirhead et al., 2006; Robinet et al., 2009). The Panel assessed the rate of natural spread as both processes together (spilling over at the edge and generation of satellite populations that later merge) determine the spatial expansion of a population.
An evidence dossier on spread was assembled from the literature review. Because no data was available to estimate the natural spread capacities of L. orbonalis, particular attention was given to available data on the spread of L. laisalis, an African species closely related to L. orbonalis that established a population in Southern Spain in the 1950s and has spread further on the southern Iberian Peninsula in the past decades. These spread data were used to estimate the expected spread rate of L. orbonalis. The assessment of spread of L. orbonalis considered both natural dispersal and farm‐scale human‐assisted spread with agricultural equipment (Section 3.3 Spread). Assessors took part in the semi‐formal EKE using behavioural aggregation (EFSA, 2014). The collected evidence was reviewed during the EKE and is summarised in Appendix E: Spread.
2.3.1. Scenario definition for spread
Scenario for spread: considering existing practices and phytosanitary measures
To estimate the lag period and rate of linear range expansion, the Panel developed a general scenario with the following description:
The pest initiates a founder population at a single point somewhere within the area of potential establishment (where the CLIMEX EI is greater than the minimum threshold (EI 15 or 30) (see Sections 2.2.3 and 3.2 Establishment).
L. orbonalis is a specialised feeder of Solanum spp. For the EU, 48 species of Solanum are reported (Valdés, 2012), all of which can be considered putative host plants for L. orbonalis.
During the lag period, the population size increases until it reaches a local steady state in the centre of the population (determined by the habitat‐carrying capacity).
By reaching the local habitat‐carrying capacity (saturation), the population enters the spread phase, pushing the outer edge of the saturated population at a constant rate into suitable, unoccupied neighbouring habitats.
The spread assessment considers the outcome of the combined contributions of natural and local human‐assisted spread. The human‐assisted component only includes operations related to production and local movement (e.g. common agricultural practices) but no post‐harvest movements, such as the trade in commodities (EFSA, 2019).
Spread occurs within regions where the CLIMEX EI Index is greater than the thresholds (EI 15 or 30) (see Establishment).
Uncertainties:
Allee effects (already considered during the assessment of establishment) might have an important impact on the survival/extinction of small founder populations.
2.4. Impact
The scientific literature on L. orbonalis was screened for information on impact of the pest on host plants. An evidence dossier on impact was assembled by EFSA staff and Working Group members. It was analysed to conceptualise the impact elements of risk and to inform the assessment of impact using EKE. Two scenarios were considered in the information retrieval from the literature: (1) yield lost under pesticide‐free treatments, (2) yield lost despite the use of pesticides. Data were extracted and analysed separately in meta‐analyses to determine the damage done by L. orbonalis with and without chemical control. The results of meta‐analysis were used as input for the EKE on impact of L. orbonalis on host plants in the EU in NUTS2 regions where L. orbonalis is able to establish, with and without specific controls in place.
The collected evidence was reviewed during the EKE and is summarised in Appendix F: Impact.
2.4.1. Scenario definition for impact
Scenario Impact‐1 (baseline): assuming no pest control is applied (i.e. artificial situation, akin to experimental no treatment ‘control’ plots in an experimental trial).
To estimate potential impact in terms of yield loss, a scenario with the following characteristics was defined:
The pest has spread to its maximum geographic extent in EU NUTS2 regions with climate suitable for establishment and equilibrium population levels.
Within the area of potential establishment, pest presence depends on the heterogeneity of the patches where the host occurs. It is therefore not necessarily the case that the pest is present in all suitable patches.
In each location where the pest occurs, its abundance is in equilibrium with the available resources (e.g. host plants) and environmental conditions (including climate, ecosystem resistance and resilience) (e.g. Grimm & Wissel, 1997).
No action is taken for pest control – yield loss data (% of fruit yield) in control plots of field trials were extracted from the literature and the subsequent meta‐analysis used to inform losses when no control options are applied (representing worst‐case conditions).
Current crop production practices (e.g. chemical insecticides targeted at L. orbonalis are not used).
The assessment of impact assumes a situation in which L. orbonalis has been established in a climatically suitable area (EI ≥ minimum threshold) for a long enough period of time to have reached carrying capacity and maximum impact.
Potential impact of transient populations was not considered i.e. in NUTS2 regions with low suitability for establishment (EI < minimum threshold) or production of eggplants in greenhouses.
Different susceptibilities of host plants (e.g. eggplants, potatoes and tomatoes), and the detailed biological characteristics of L. orbonalis (e.g. dispersal, feeding activity) were not considered in the assessment of impact.
The focus was on eggplant in the southern EU, largely Mediterranean coastal areas.
Scenario Impact‐2 (with pest management in place): considering existing practices and any additional pest management by farmers to target the pest.
To estimate potential impact in terms of yield loss under scenario 2, the Panel envisaged scenario 1 with the following additional conditions:
Pest control practices would be applied by farmers.
Cropping practices and management options are those currently in place in the area of potential pest distribution, considering differences with those applied in countries where L. orbonalis is present (and evidence was collected).
The effect of currently applied control against other pests is taken into account (e.g. yield losses in EU crops given existing pest pressures were considered – how much more would Leucinodes add to the existing burden of pests in the EU?).
In a scenario where the pest is widely established and there would be no statutory action by NPPOs in the EU against Leucinodes.
2.5. Evaluation of risk reduction options/risk mitigation measures
As noted in Section 1.2, the EFSA PLH Panel planned to evaluate how additional risk mitigation measures (Appendix G) may reduce the likelihood of pest entry. However, results from the entry and establishment modelling reveal that the initiation of L. orbonalis founder populations in the EU is already unlikely given the relatively small quantities of product imported and taking existing practices into account (see Section 3.2.7). Consequently, options for further risk reduction are discussed, but their effectiveness was not quantified. Therefore, options for further risk reduction are discussed, but their effectiveness was not quantified. This opinion therefore presents an assessment of pest risk based on historic trade volumes, existing practices and generic phytosanitary measures.
2.6. Temporal and spatial scales
The pathway model calculates the trade flow of relevant commodities per year, on average, over the next 5 years (2024–2028).
The distribution of potentially infested plant material entering the EU was assessed using NUTS2 spatial resolution using EU census data from 2021 (Eurostat, accessed 31/12/2022). The CLIMEX model used 30 years of climate data, ranging from 1981 to 2010.
3. ASSESSMENT
A description of taxonomic issues relating to the genus Leucinodes is provided in Appendix A. As L. orbonalis occurs only in Asia, all pathways were considered to originate in Asia.
A synthesis of the biology of L. orbonalis based on the literature review is provided in Appendix B together with some exemplary pictures of the pest and the damage it causes. A list of cultivated and wild hosts is provided in the pest categorisation for L. orbonalis (EFSA PLH Panel, 2021). Female adults lay eggs singly or in small clusters of two to four on the lower leaf surfaces of the topmost and middle leaves of its preferred host plant, S. melongena (Ardez et al., 2008). The eggs are oval, about 0.5 mm in diameter, and turn from their initial creamy white colour to a deep orange towards larval hatching, when the black head capsule becomes visible through the eggshell (Ardez et al., 2008; Lall & Ahmad, 1965). The larvae are internal feeders, boring into the shoots and fruits, the latter being the preferred host tissue (Navasero & Calilung, 1990) and usually harbouring one or two larvae (Shukla, 1986). The entry hole into the plant tissue is closed by a plug of excreta. The concealed larval feeding makes infestation difficult to detect. However, in host plants with fruits too small to harbour the growing larva (e.g. Solanum nigrum), the larva exits the fruit and webs together three or four fruits to continue feeding from inside the web (Das & Patnaik, 1971). Such behaviour is seen with infested S. torvum.
3.1. Entry
3.1.1. Analysis of interceptions on produce
The Panel searched for interceptions of L. orbonalis in Europhyt (1995 until May 2020) and TRACES (June 2020 to ongoing database, last check 7 March 2023).
In the years 2004–2023, there were 350 notifications of interceptions of L. orbonalis in Europhyt and TRACES from consignments of plant products originating from Asia. The majority of plant species with interceptions are in the Solanaceae, but a number of species from other families are also reported (Appendix C, Table C.1). The Panel analysed the host status of plants on which interceptions have been reported to distinguish actual pathways of introduction (commodities in which the insect can develop to a viable pupa and adult) and incidental interceptions due to movement of larvae from an actual host to other plant material in the same shipment. The Panel judged all interceptions on plant material from non‐solanaceous plant products as incidental. Details on the underlying evidence are provided below.
The primary potential pathways for introduction based on the numbers of interceptions are fruit of S. melongena (222 interceptions) and S. torvum (66 interceptions). Additional pathways are fruit of S. virginianum (8 interceptions), S. aculeatissimum, S. aethiopicum, S. anguivi and S. macrocarpon (2 interceptions in each of the four species), S. stramoniifolium and S. undatum (1 interception in both species) and undetermined Solanum spp. (26 interceptions). All these Solanum spp. are confirmed hosts of L. orbonalis based on literature reports (Appendix C, Table C.1).
Four interceptions were made on Momordica spp. A comment in the interception data indicates a misidentification, the actual species intercepted being Diaphania indica (Saunders) (Lepidoptera: Crambidae), which is, like L. orbonalis, a predominantly white moth with similar‐looking larvae that primarily feed on Cucurbitaceae such as Momordica. Maureal et al. (1982) observed in a no‐choice trial that third‐instar L. orbonalis larvae did not feed on Momordica charantia fruit offered for 24 h. Momordica was therefore disregarded as a pathway.
Another case considered as potential pathway among non‐Solanaceae interceptions was Ipomoea aquatica (Convolvulaceae). L. orbonalis has been reported to feed on tubers of the related sweet potato, I. batatas, but lab‐rearing experiments showed that L. orbonalis cannot successfully complete its life cycle from egg to adult on this plant (Ardez et al., 2008). I. batatas was therefore judged not to offer a pathway.
One interception was reported on Capsicum annuum. Maureal et al. (1982) showed in a 24‐h no‐choice test that the larvae of L. orbonalis do not feed on C. annuum fruit; hence, C. annuum was not considered a pathway.
An overview of the interceptions on Solanum spp. is provided in Figures 5, 6–7 below.
FIGURE 5.
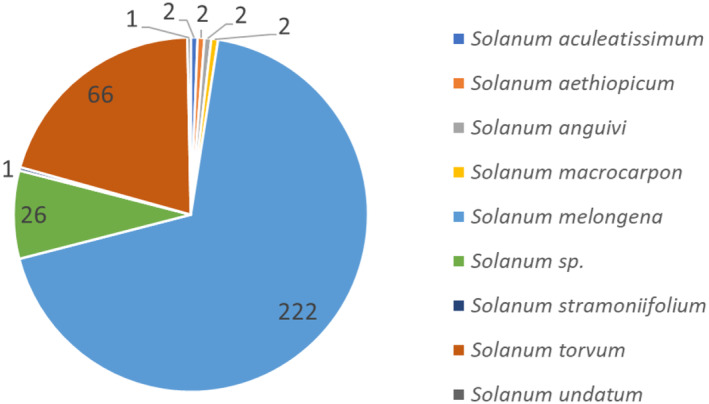
Interceptions of Leucinodes orbonalis on Solanum spp. imported from Asia according to records in Europhyt and TRACES, 2004–2023. The total number of interceptions is 324. While some records in Europhyt and TRACES do not identify the species, the product originates from countries where only L. orbonalis is present (except perhaps Vietnam, which has a genetically distinct Leucinodes population which has to date not been described as a species). This indicates that for all countries (including Vietnam until the species is described), the species is L. orbonalis.
FIGURE 6.
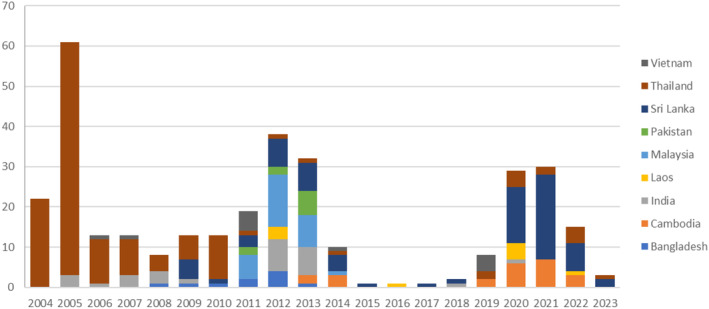
Interceptions of Leucinodes orbonalis and Leucinodes spp. from Asia on different Solanum spp. according to records of Europhyt and TRACES, 2004–2023. The figure highlights that the countries of origin of interceptions vary substantially by date. This variation reflects both variation in trade volume from those countries and variation in the frequency of interceptions per unit product imported (Appendix C). While some records in Europhyt and TRACES do not identify the species, the product originates from countries where only L. orbonalis is present (except perhaps Vietnam, which has a genetically distinct Leucinodes population which has to date not been described as a species). This indicates that for all countries (including Vietnam until the species is described), the species is L. orbonalis.
FIGURE 7.
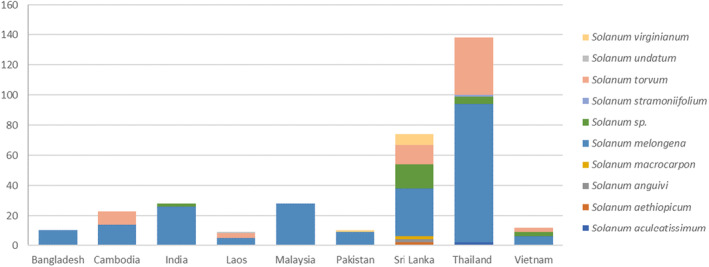
Interceptions of Leucinodes orbonalis and Leucinodes sp. from different countries of origin in Asia on different Solanum spp. according to records of Europhyt and TRACES, 2004–2023. While some records in Europhyt and TRACES do not identify the species, the product originates from countries where only L. orbonalis is present (except perhaps Vietnam, which has a genetically distinct Leucinodes population which has to date not been described as a species). This indicates that for all countries (including Vietnam until the species is described), the species is L. orbonalis.
3.1.2. Identifying pathways (plants for planting)
Plants for planting of Solanaceae, other than seeds, are largely prohibited from entering the EU except from a few European and Mediterranean countries and parts of European Russia (Commission Implementing Regulation (EU) 2019/2072, Annex VI, 18). Annex VI prohibitions also concern potato (S. tuberosum), which has more detailed prohibitions. Thus, the Panel concludes that, under the current regulation, plants for planting are not a pathway for entry of L. orbonalis into the EU.
3.1.3. Identifying pathways: Plant hosts of L. orbonalis
In Asia, the single most important larval host plant of L. orbonalis is Solanum melongena, commonly known as eggplant, brinjal or aubergine. The larvae feed in the stems and fruits of the plant. When given free choice for ovipositing among eggplant, tomato (S. lycopersicum), potato (S. tuberosum), black nightshade (S. nigrum) and three non‐Solanaceae crops, females chose exclusively eggplant (Ardez et al., 2008).
The next most‐preferred host plant in India is S. aethiopicum (= S. gilo, S. integrifolium) (Dr S. Ramasamy, personal communication), commonly known as gilo, or as garden egg in Africa (dr L. C. Nwosu, personal communication). Tejavathu et al. (1991) report an infestation rate of 3.5% of shoots and 4.4% of fruits in India. The fruits have a diameter of 10–50 mm but are usually 15–25 mm in size (Vorontsova, 2023).
Apart from its primary host (eggplant), L. orbonalis is able to successfully complete its life cycle on several other Solanum species such as S. torvum (turkey berry), S. nigrum (black nightshade), S. tuberosum (potato), S. lycopersicum (tomato), S. anomalum, S. macrocarpon, S. myriacanthum (Himalayan nightshade), S. viarum (tropical soda apple) and S. virginianum (yellow‐fruit nightshade); see Appendix C for more details about the biology of L. orbonalis on these plants. In S. lycopersicum, the larvae can only develop to pupation in the stems, but there is uncertainty whether they can develop into viable adults in the fruit due to the high‐water content; hence, tomato fruit are not considered a pathway (personal communication Dr S. Ramasamy; Ardez et al., 2008; Boopal et al., 2013; Das & Patnaik, 1971).
The EU does import a small amount of ware potatoes from Asia (e.g. Israel) and although literature reports L. orbonalis infesting stems of S. tuberosum in India and the larvae being able to feed and complete their development on potato tubers in the laboratory, the larvae do not infest underground tubers under field conditions. Given lack of imports and that larvae do not infest tubers under practical growing conditions, potato tubers were not considered a realistic pathway (Appendix C, Table C.1 provides further details).
Physalis and Capsicum have been cited as host plants in the literature, but the Panel could not find any reference confirming that L. orbonalis larvae are actually feeding on these plant genera; Maureal et al. (1982) demonstrated that the larvae do not feed on Capsicum. Other Solanaceae plants of economic importance, such as tobacco (Nicotiana) and thornapples (Datura), have neither been reported as hosts of L. orbonalis, nor has the species been intercepted in the EU on plants of these genera.
Records of L. orbonalis on non‐Solanaceae host plants appear to be incidental, and the species is unlikely to complete its life cycle on these plants. Ardez et al. (2008) conducted no‐choice experiments including some non‐Solanaceae crops and found that either the larvae could not survive feeding on sweet potato (Ipomoea batatas, Convolvulaceae) and okra (Abelmoschus esculentus, Malvaceae), or that feeding on cowpea (Vigna unguiculata = syn. Vigna sinensis, Fabaceae) was successful, but no adults emerged from the formed pupae.
The only report of L. orbonalis feeding on mango (Mangifera indica) is from an abstract of Hutson (1931), stating: ‘the shoot‐borer, Leucinodes orbonalis, Gn., and the pentatomid, Coptosoma siamica, Wlk. (Hemiptera: Pentatomidae), on mango’. This abstract refers to a 17‐page typescript article that was apparently never published. Mango has not been confirmed as larval host of L. orbonalis by any other resource, and the Panel therefore considers it an erroneous host plant.
Momordica (bitter melon, Goya) was repeatedly mentioned as host plant in the literature, however, always without a reference. In the Europhyt database, an interception L. orbonalis on Momordica was probably a misidentification of Diaphania indica, another species of moth in the Crambidae family. The larvae of Diaphania species commonly feed on Cucurbitaceae such as Momordica (Solis, 2006). The Panel therefore considers Momordica an erroneously reported host for L. orbonalis.
In conclusion, the Panel identified Solanum species in the Solanaceae plant family as potential produce pathway for L. orbonalis to enter the EU. The Panel focused on S. melongena and S. torvum (Table 3) as main entry pathways, but considered that fruit of any species of Solanum might act as a pathway. However, due to less trade and fewer interceptions compared to S. melongena and S. torvum, these pathways were not considered during the quantitative assessment.
TABLE 3.
Host plants considered potential entry pathways for Leucinodes orbonalis.
| Binomial name | Common English names | Interceptions in the EU |
|---|---|---|
| Solanum melongena | Eggplant, aubergine, brinjal | Yes |
| Solanum torvum | Turkey berry, pea eggplant | Yes |
3.1.4. Pathway evaluation (EKE results)
Key results from the entry pathway model are shown in Table 4 below. Results represent model outputs for scenario 1 where 25% of pathway imports are turkey berry and 75% are eggplants; imports are distributed for consumption across the EU according to human population; NUTS2 regions where EI > 15 are considered NUTS regions where establishment is possible. Details of the source of the data used for the estimation and the calculations can be found in Appendix C.
TABLE 4.
Model output results illustrating the range in estimates of mean imports and subsequent range in number of infested host transfer units entering the EU each year into areas where climate may be suitable in scenario 1 (EI > 15).
| Percentile (%) | 1 | 5 | 25 | 50 | 75 | 95 | 99 |
|---|---|---|---|---|---|---|---|
| Mean import of fresh turkey berry from Asia into the EU (t/year) | 18.6 | 25.1 | 34.5 | 41.0 | 47.5 | 56.7 | 63.1 |
| Import of fresh eggplant fruit from Asia into the EU (t) | 55.8 | 75.4 | 103.2 | 122.6 | 142.0 | 170.0 | 189.8 |
| Turkey berry allocated to NUTS2 (t) | 4.3 | 5.8 | 8.0 | 9.5 | 11.0 | 13.1 | 14.6 |
| Eggplant allocated to NUTS2, (t) | 12.9 | 17.4 | 2.9 | 28.4 | 32.9 | 39.3 | 43.8 |
| Range of weight in turkey berry clusters (g) | 18.0 | 20.3 | 27.3 | 34.5 | 42.4 | 51.4 | 55.0 |
| Range in weight of eggplant fruit (g) | 100.1 | 112.6 | 151.7 | 196.6 | 253.3 | 342.8 | 400.6 |
| Number of turkey berry transfer units entering NUTS2 | 107,313 | 146,230 | 211,587 | 271,691 | 354,590 | 511,462 | 626,254 |
| Number of eggplant transfer units entering NUTS2 | 50,103 | 69,456 | 105,016 | 141,054 | 189,933 | 275,507 | 339,899 |
| Number of total transfer units entering NUTS2 | 174,555 | 237,386 | 340,186 | 426,956 | 533,138 | 715,591 | 863,327 |
| Number of infested transfer units per 10,000 units imported | 0.10 | 0.13 | 0.98 | 4.20 | 11.13 | 23.73 | 30.07 |
| Number of infested transfer units entering NUTS2 | 3.3 | 5.5 | 40.4 | 172.6 | 468.8 | 1100.9 | 1627.0 |
Notes: Scenario 1 considers an establishment index, EI > 15 and turkey berry and eggplant representing 25% and 75%, respectively, of total imports of the commodity code HS 0709 3000. Imports are from the following countries: Bangladesh, Brunei, China, Indonesia, India, Japan, Cambodia, Laos, Sri Lanka, Myanmar, Malaysia, Nepal, Philippines, Pakistan, Singapore, Thailand, Taiwan and Vietnam.
Results from Scenario 4 are shown in Table 5.
TABLE 5.
Model output results illustrating the range in estimates of mean imports and subsequent range in number of infested host transfer units entering the EU each year into areas where climate may be suitable (EI > 30).
| Percentile (%) | 1 | 5 | 25 | 50 | 75 | 95 | 99 |
|---|---|---|---|---|---|---|---|
| Import of fresh turkey berry from Asia into the EU (t) | 18.6 | 25.1 | 34.5 | 41.0 | 47.5 | 56.7 | 63.1 |
| Import of fresh eggplant fruit from Asia into the EU (t) | 55.8 | 75.4 | 103.2 | 122.6 | 142.0 | 170.0 | 189.8 |
| Turkey berry allocated to NUTS2 (t) | 2.6 | 3.5 | 4.9 | 5.8 | 6.7 | 8.0 | 8.9 |
| Eggplant allocated to NUTS2 (t) | 7.9 | 10.6 | 14.6 | 17.3 | 20.0 | 240. | 26.7 |
| Range of weight in turkey berry clusters (g) | 18.0 | 20.3 | 27.3 | 34.5 | 42.4 | 51.4 | 55.0 |
| Range in weight of eggplant fruit (g) | 100.1 | 112.6 | 151.7 | 196.6 | 253.3 | 342.8 | 400.6 |
| Number of turkey berry transfer units entering NUTS2 | 65,420 | 89,145 | 128,987 | 165,627 | 216,164 | 311,796 | 381,775 |
| Number of eggplant transfer units entering NUTS2 | 30,543 | 42,342 | 64,019 | 85,989 | 115,786 | 167,954 | 207,208 |
| Number of total transfer units entering NUTS2 | 106,412 | 144,714 | 207,383 | 260,279 | 325,010 | 436,236 | 526,299 |
| Number of infested transfer units per 10,000 units imported | 0.10 | 0.13 | 0.98 | 4.20 | 11.13 | 23.73 | 30.07 |
| Number of infested transfer units entering NUTS2 | 2.0 | 3.4 | 24.6 | 105.2 | 285.8 | 671.2 | 991.8 |
Note: Scenario 4 considers an establishment index, EI ≥ 30, and turkey berry and eggplant representing 25% and 75%, respectively, of total imports of the commodity code HS 0709 3000.
3.1.5. Unquantified uncertainties affecting the assessment of entry
The proportion of imports which are turkey berry (S. torvum) and which are eggplant (S. melongena) is unknown.
Volumes of eggplant imports from Asia may change in future; some Asian type varieties are being grown in the EU already, hence imports may fall.
Growing conditions for material intended for export to the EU; amount of infestation at origin and effectiveness of cleaning/sorting.
On arrival in the EU, there is uncertainty on the distribution of consignment sizes, the percentage of consignments inspected in each country, the sample size at inspection and the chance of detection of infestation if an inspector examines an eggplant or turkey berries.
Estimation of interceptions based on incomplete information (not all EU member states notify interceptions of non‐quarantine pests).
3.1.6. Conclusion on the assessment of entry
The pathway most likely to provide a route for entry of L. orbonalis into the EU was judged to be fresh eggplant and fresh turkey berry from Asia. The number of fruits expected to enter the EU each year and be distributed across NUTS regions where climatic conditions are conducive for establishment of L. orbonalis is expected to be in the order of hundreds of thousands (median estimate with EI≥15 approximately 430,000 transfer units; 90% CR approximately 175,000–865,000). With an EI threshold of 30, the number of fruits entering NUTS2 regions where parts are suitable for establishment drops to approximately 260,000 (90% CR approximately 106,000–526,000).
Infested fruits represent a small proportion of the total number of fruits entering the EU. The number of transfer units infested with live L. orbonalis entering NUTS2 areas with EI ≥ 15 is estimated to be approximately 175 per year (90% CR approximately 6–1100); using an EI threshold of 30, the median number of infested transfer units drops to approximately 105 per year (90% CR approximately 3–670). In the scenario where only turkey berry is imported, and using EI≥ 15, the 95 percentile estimates 84 adults emerge in areas suitable for establishment. In contrast, where only eggplant is imported and using EI ≥ 30, the 5 percentile estimates one adult emerging in 40 years.
3.2. Establishment
Climatic mapping is a common approach to identify new areas that might provide suitable conditions for the establishment of alien organisms (Baker, 2002; Venette, 2017). Climatic mapping is based on combining information on climate in the known distribution of a poikilothermic organism, the organisms' physiological responses to environmental conditions and the climate in the risk area. The current distribution of L. orbonalis is presented in Section 3.2.1. The results of climatic mapping are presented in Sections 3.2.2, 3.2.3, 3.2.4–3.2.5. SDM results were not included in the current opinion because the results were found to be not mature for interpretation. The details about L. orbonalis climate suitability modelling are presented in Rossi et al. (2023) and available online on the ZENODO platform.
3.2.1. Global distribution of Leucinodes orbonalis
L. orbonalis is a tropical and sub‐tropical species native to Asia with India and Bangladesh thought to be its centre of origin (Karthika et al., 2019). Figure 8 shows the distribution of L. orbonalis. Until Hayden et al. (2013) and Gilligan and Passoa (2014) reported that L. orbonalis was restricted to Asia, it was thought that L. orbonalis also occurred in sub‐Saharan Africa. Literature previously reporting L. orbonalis from Africa should be regarded as referring to members of a complex of other species of Leucinodes native to Africa and not as referring to L. orbonalis (see also Section 1.2).
FIGURE 8.
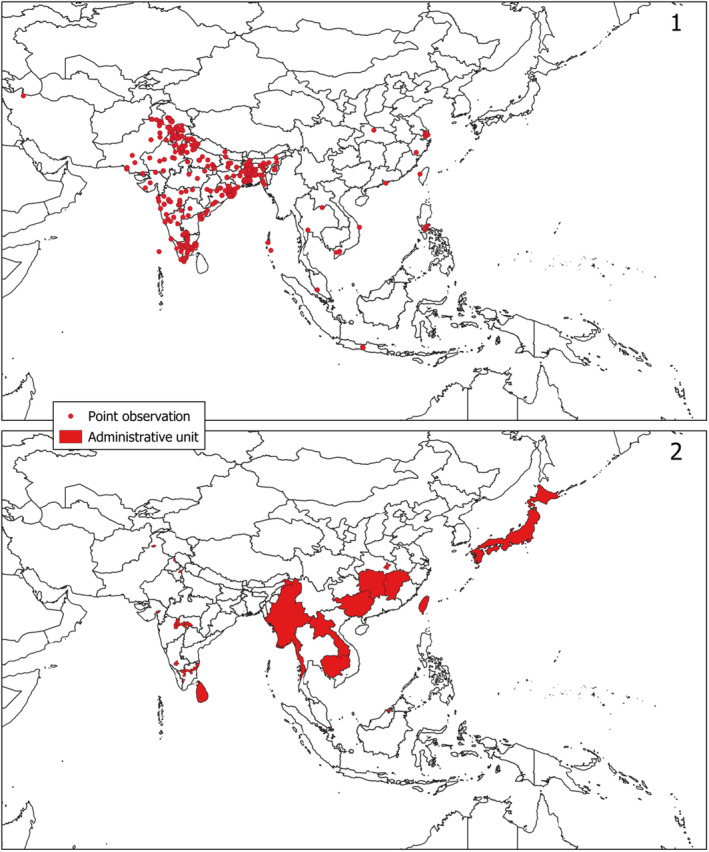
Detailed distribution of Leucinodes orbonalis in Asia based on a systematic literature search showing (1) points and (2) administrative regions where the organism occurs.
3.2.2. Degree days model
Data from Dhaliwal and Aggarwal (2021) were used to determine the base temperature of 15°C and a thermal constant of 438.6 degree days above the threshold to complete a generation. Based on this, between one and four generations could theoretically be possible in Europe each year. This compares to the 8–10 or more generations per year across large parts of India and southeast Asia (Figures 9 and 10) where L. orbonalis is a recognised pest.
FIGURE 9.
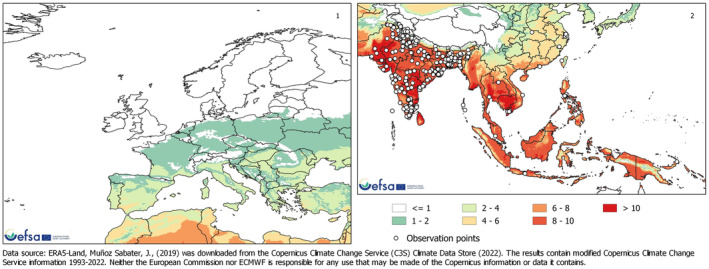
Estimated mean number of generations for Leucinodes orbonalis in Europe and the Mediterranean Basin (1) and southeast Asia and India (2). Number of generations were calculated considering a minimum number of accumulated degree days to complete one generation of 438.6 Degree days. Number of generations were calculated for each year in the period 1993–2022 and then averaged.
FIGURE 10.
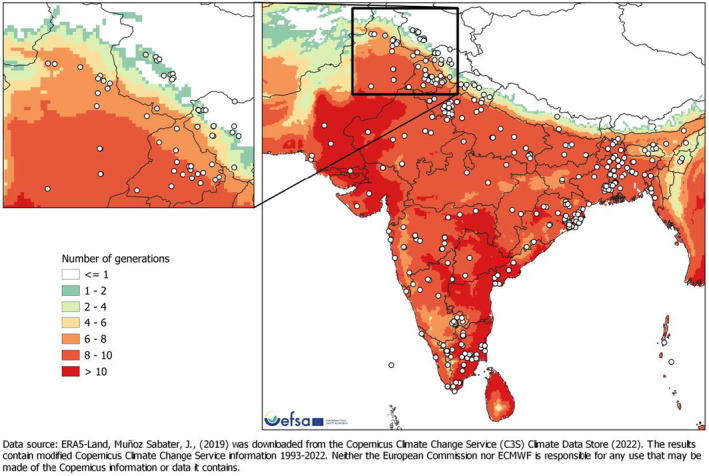
Estimated mean number of Leucinodes orbonalis generations in India and neighbouring regions.
Mountainous regions in northern India are much cooler than central and southern India. The accumulated heat (measured as DD) is therefore much less in the mountains leading to fewer generations. Note that L. orbonalis presence has been reported in areas where degree day mapping suggests a temperature sum supporting less than one is possible. Possible explanations for this include:
Findings could be from a transient population.
Mapping data are based on average temperatures over several years and reported findings could be from unusually warm periods.
L. orbonalis surviving in a ‘hot spot’ above the average of the grid cell of mapping resolution.
The reports are from trap catches of individuals moving from suitable neighbouring areas.
3.2.3. CLIMEX
A more sophisticated modelling approach to heat accumulation is the use of CLIMEX which takes organism ecophysiological requirements into account across a geographic region. Results from the CLIMEX model outputs for EI across India and southeast Asia are shown in Figure 11A,B. Green dots in Asia indicate point locations where L. orbonalis has been reported.
FIGURE 11.
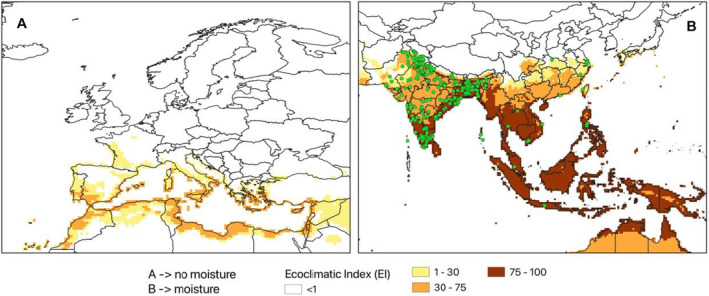
(A, B) CLIMEX Ecoclimatic Index for Leucinodes orbonalis (i) India and southeast Asia, and (ii) the Euro‐Mediterranean Basin.
CLIMEX modelling indicates EI values ≥ 30 across most of India and southeast Asia. In northern cooler areas, and at higher altitudes, EI values drop (Figure 11A). EI values that can support establishment are found in southern Europe, especially around the Mediterranean coast (Figure 11B).
The details about L. orbonalis climate suitability modelling are presented in Rossi et al. (2023) and available online on the ZENODO platform.
3.2.4. Establishment
Pest interceptions indicate that L. orbonalis can enter the EU (Appendix C). Modelling estimates that, depending on the EI threshold used for establishment, the number of transfer units infested with live L. orbonalis entering NUTS2 areas where establishment may be possible varies from a median of approximately 175 per year (EI ≥ 15) to approximately 105 per year (EI ≥ 30). To initiate a founder population, larvae entering the EU with infested S. melongena or S. torvum must complete development, locate a partner, mate and the subsequent progeny must survive, complete development and mate, thus initiating a founder population (Appendix D).
Table 6 provides key results from the EKE modelling. It shows the likelihood that a potential founder population will be initiated in the EU each year. Using an EI threshold of 15, the median number of founder populations establishing in the EU annually is 0.00014 (90% CR 0.00000–0.00264). This equates to a median estimate of one founder population approximately every 7000 years (90% CR approximately one every 380–355,600 years).
TABLE 6.
Model output results illustrating the range in estimates for key model steps from entry to initiation of founder population.
| Percentile (%) | 1 | 5 | 25 | 50 | 75 | 95 | 99 |
|---|---|---|---|---|---|---|---|
| Estimated mean annual number of total transfer units entering NUTS2 | 174,555 | 237,386 | 340,186 | 426,956 | 533,138 | 715,591 | 863,327 |
| Number of infested transfer units per 10,000 units imported | 0.10 | 0.13 | 0.98 | 4.20 | 11.13 | 23.73 | 30.07 |
| Estimated mean number of infested transfer units entering NUTS2 | 3.3 | 5.5 | 40.4 | 172.6 | 468.8 | 1100.9 | 1627.0 |
| Number of infested transfer units discarded in NUTS 2 by trade (pest may survive) | 0.3 | 0.6 | 4.6 | 19.9 | 55.5 | 145.0 | 237.3 |
| Number of infested transfer units discarded by consumer in NUTS2 (pest may survive) | 1.1 | 2.2 | 15.8 | 68.3 | 185.5 | 488.4 | 784.4 |
| Number of emerged adults surviving in NUTS2 areas | 0.03 | 0.09 | 0.72 | 3.16 | 9.87 | 32.29 | 61.25 |
| Number of founder populations in NUTS2 areas per year | 0.00000 | 0.00000 | 0.00003 | 0.00014 | 0.00056 | 0.00264 | 0.00626 |
| Expected number of years between founder populations | 160 | 379 | 1782 | 7006 | 35,590 | 355,600 | 1,760,100 |
Note: Scenario 1 considers an establishment index (EI) ≥ 15 and turkey berry and eggplant representing 25% and 75%, respectively, of total imports of the commodity code HS 0709 3000.(Scenario 1 in Table 2).
Using EI ≥ 30 as a threshold for establishment, Table 7 key results from the pathway modelling. It shows that the median likelihood that a founder population will be initiated in the EU is 0.00009 per year (90% CR 0.00000–0.00161). This equates to a median estimate of one founder population approximately every 11,500 years (90% CR approximately one every 620–583,300 years).
TABLE 7.
Model output results illustrating the range in estimates for each model step from entry to initiation of founder population.
| Percentile (%) | 1 | 5 | 25 | 50 | 75 | 95 | 99 |
|---|---|---|---|---|---|---|---|
| Number of total transfer units entering NUTS2 | 106,412 | 144,714 | 207,383 | 260,279 | 325,010 | 436,236 | 526,299 |
| Number of infested transfer units per 10,000 units imported | 0.10 | 0.13 | 0.98 | 4.20 | 11.13 | 23.73 | 30.07 |
| Number of infested transfer units entering NUTS2 | 2.0 | 3.4 | 24.6 | 105.2 | 285.8 | 671.2 | 991.8 |
| Number of infested transfer units discarded in NUTS 2 by trade (pest may survive) | 0.2 | 0.4 | 2.8 | 12.1 | 33.8 | 88.4 | 144.7 |
| Number of infested transfer units discarded by consumer in NUTS2 (pest may survive) | 0.7 | 1.3 | 9.7 | 41.6 | 113.1 | 297.8 | 478.2 |
| Number of emerged adults surviving in NUTS2 areas | 0.02 | 0.06 | 0.44 | 1.93 | 6.02 | 19.69 | 37.34 |
| Number of founder populations in NUTS2 areas per year | 0.00000 | 0.00000 | 0.00002 | 0.00009 | 0.00034 | 0.00161 | 0.00381 |
| Expected number of years between founder populations | 262 | 622 | 2923 | 11,493 | 58,380 | 583,316 | 2,887,223 |
Note: Scenario 4 considers an establishment index (EI) ≥ 30, and turkey berry and eggplant representing 25% and 75%, respectively, of total imports of the commodity code HS 0709 3000 (scenario 4 in Table 2).
3.2.5. Conclusions on identifying NUTS2 regions suitable for establishment
The fractions of CLIMEX grid cells in each NUTS2 region with EI ≥ 15 or ≥ 30 (Figure 12) were determined and the area where EI ≥ 15 or 30 calculated. Table 8 lists the NUTS2 regions in descending order of area.
FIGURE 12.
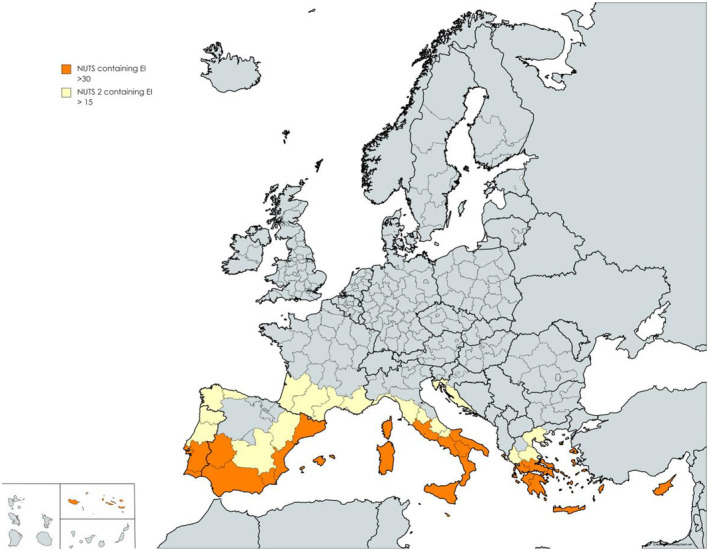
EU NUTS 2 regions that contain any areas where EI ≥ 15 (pale yellow) or EI ≥ 30 (orange). These areas account for 23% (EI ≥ 15) or 14% (EI ≥ 30) of EU human population and where we assume 23% and 14% of imported commodities are distributed.
TABLE 8.
Area of NUTS2 regions where establishment may be possible, based on EI thresholds of 15 or 30.
| NUTS2 code | Country (NUTS name) | % of EU population | Area EI ≥ 15 (thousands km2) | Area EI ≥ 30 (thousands km2) |
|---|---|---|---|---|
| ES61 | ES (Andalucía) | 1.93 | 67.50 | 47.50 |
| PT18 | PT (Alentejo) | 0.16 | 35.00 | 17.50 |
| ITG1 | IT (Sicilia) | 1.09 | 25.00 | 17.50 |
| ES52 | ES (Comunidad Valenciana) | 1.15 | 15.00 | 12.50 |
| EL43 | EL (Kriti) | 0.14 | 12.50 | 12.50 |
| CY00 | CY (Kýpros) | 0.20 | 10.00 | 10.00 |
| ES43 | ES (Extremadura) | 0.24 | 37.50 | 7.50 |
| ITF6 | IT (Calabria) | 0.42 | 15.00 | 7.50 |
| ITF4 | IT (Puglia) | 0.89 | 17.50 | 6.40 |
| EL42 | EL (Notio Aigaio) | 0.07 | 5.17 | 5.16 |
| ITG2 | IT (Sardegna) | 0.36 | 22.50 | 5.00 |
| EL41 | EL (Voreio Aigaio) | 0.04 | 5.00 | 5.00 |
| EL65 | EL (Peloponnisos) | 0.12 | 7.03 | 4.58 |
| ES53 | ES (Illes Balears) | 0.28 | 5.18 | 3.69 |
| ES62 | ES (Región de Murcia) | 0.34 | 10.00 | 3.31 |
| EL30 | EL (Attiki) | 0.86 | 3.88 | 3.02 |
| EL64 | EL (Sterea Elláda) | 0.11 | 5.00 | 1.96 |
| EL63 | EL (Dytiki Elláda) | 0.15 | 7.50 | 1.88 |
| ITI4 | IT (Lazio) | 1.29 | 12.50 | 1.43 |
| PT17 | PT (Área Metropolitana de Lisboa) | 0.65 | 5.00 | 1.36 |
| ES51 | ES (Cataluña) | 1.74 | 17.50 | 1.25 |
| ITF3 | IT (Campania) | 1.27 | 3.90 | 1.20 |
| ITF5 | IT (Basilicata) | 0.12 | 2.36 | 1.07 |
| EL62 | EL (Ionia Nisia) | 0.05 | 5.00 | 0.70 |
| PT20 | PT (Região Autónoma dos Açores) | 0.05 | 5.00 | 0.44 |
| MT00 | MT (Malta) | 0.12 | 0.31 | 0.31 |
| PT15 | PT (Algarve) | 0.11 | 7.50 | 0.25 |
| FRM0 | FR (Corse) | 0.08 | 5.00 | 0.22 |
| PT30 | PT (Região Autónoma da Madeira) | 0.06 | 0.76 | 0.04 |
| PT16 | PT (Centro (PT)) | 0.52 | 20.00 | – |
| FRI1 | FR (Aquitaine) | 0.81 | 12.50 | – |
| ES24 | ES (Aragón) | 0.30 | 10.00 | – |
| FRL0 | FR (Provence‐Alpes‐Côte d'Azur) | 1.18 | 7.50 | – |
| FRJ1 | FR (Languedoc‐Roussillon) | 0.67 | 5.00 | – |
| ITI1 | IT (Toscana) | 0.84 | 5.00 | – |
| PT11 | PT (Norte) | 0.82 | 3.73 | – |
| ES42 | ES (Castilla‐La Mancha) | 0.47 | 3.06 | – |
| EL54 | EL (Ipeiros) | 0.07 | 2.67 | – |
| HR03 | HR (Jadranska Hrvatska) | 0.29 | 2.25 | – |
| ES11 | ES (Galicia) | 0.63 | 2.17 | – |
| EL52 | EL (Kentriki Makedonia) | 0.41 | 1.45 | – |
| ITC3 | IT (Liguria) | 0.34 | 0.98 | – |
| FRJ2 | FR (Midi‐Pyrénées) | 0.73 | 0.87 | – |
| ITF1 | IT (Abruzzo) | 0.29 | 0.66 | – |
| ES12 | ES (Principado de Asturias) | 0.23 | 0.52 | – |
| ITF2 | IT (Molise) | 0.07 | 0.45 | – |
| EL61 | EL (Thessalia) | 0.17 | 0.21 | – |
| ITI2 | IT (Umbria) | 0.19 | 0.09 | – |
| Sum % EU population in regions containing EI ≥30 | 14.09 | |||
| Sum % EU population in regions containing EI ≥15 | 23.12 | |||
| Sum area (km 2 ) | 517.69 | 248.26 | ||
3.2.6. Introduction of L. orbonalis into the EU
For successful introduction, a pest that enters the EU must initiate a founder population. For a species that reproduces sexually and enters the EU in a virgin state, establishing a founder population requires mate finding and finding a host on which progeny can develop. There should be some spatial and temporal synchrony in emergence of mating partners and with hosts for success. The Panel used Monte Carlo simulations with a probabilistic pathway model to assess the number of eggplant fruit or bunches of turkey berry entering each year into those parts of the EU that are suitable for establishment. The model then quantifies the subsequent steps of waste production, escape of adult insects from waste, mating and initiation of a founder population by an egg‐laying female. Within the EU imported S. melongena and S. torvum are distributed in proportion to human population in NUTS2 areas.
3.2.7. Unquantified uncertainties affecting the assessment of introduction
What happens to organisms invading a new area is a field of invasion biology that is little known or understood (Puth and Post, 2005); hence, there is uncertainty regarding the effect of stochasticity of behavioural events (development, emergence, mating, host finding) leading to successful pest transfer to suitable hosts and initiation of a founder population that is sustainable. This is because such steps are largely unobserved and there is little empirical evidence around the processes involved although successful invasion is often attributed to propagule pressure (Leung et al., 2004; Simberloff, 2009). The following uncertainties were identified but are not reflected in the uncertainty of the introduction model:
Although the assessment focused on eggplants and turkey berry from Asia, other potential pathways exist, thus adding to the likelihood of entry, however, they were considered less relevant after detailed consideration of the small volumes entering; any additional entry would be marginal.
No literature could be found detailing the growing practices used in Asia to produce eggplant or turkey berry for export to the EU.
The annual quantity of imports is relatively small; some EU growers are already producing more exotic varieties of eggplants of the type currently sourced from Asia; imports from some sources have declined and imports might drop further in future or even cease.
Changes in pest management regime (e.g. ban of insecticides, ineffectiveness of pesticides due to development of resistance, increased use of biological control) can affect probability that export consignment is infested and future likelihood of establishment in opposing directions.
What happens to organisms invading a new area is a field of invasion biology that is little known or understood (Puth and Post, 2005); hence, there is uncertainty regarding the effect of stochasticity of behavioural events (development, emergence, mating, host finding) leading to successful pest transfer to suitable hosts and initiation of a founder population that is sustainable. This is because such steps are largely unobserved and there is little empirical evidence around the processes involved although successful invasion is often attributed to propagule pressure (Leung et al., 2004; Simberloff, 2009).
3.2.8. Conclusion on entry and establishment (Pest introduction)
Based on the evidence regarding host plants of L. orbonalis, and on interception data, the importation of eggplant (S. melongena) and turkey berry (S. torvum) from Asia were judged to be the most relevant pathways for entry of L. orbonalis into the EU. In a scenario where 25% of imports are turkey berry and 75% are eggplant, and with a threshold EI of 15, then using EKE and pathway modelling, the median number of infested transfer units (individual eggplant fruit or bunches of turkey berries) entering the EU annually, where climate is suitable for establishment, is estimated to be approximately 175 (90% CR approximately 6–1100). Each infested transfer unit is likely to contain only one larva (Appendix B). Assuming a minimum EI of 30 for successful establishment, the number of infested units entering into suitable regions would be 105 as a median estimate and 3–670 as a credible range.
Climatic conditions are most suitable for establishment in parts of the southern EU, especially in Spain (Andalucía, Comunidad Valenciana and Extremadura), Portugal (Alentejo), Italy (Sicilia, Calabria, Puglia), Greece (Kriti) and Malta and Cyprus. If imports are allocated in proportion to human population, between 14.09% and 23.12% of transfer units enter regions of the EU suitable for establishment (lower estimates based on EI ≥ 30, higher estimate based on EI ≥ 15). Of the infested units entering NUTS2 regions where EI ≥ 15 approximately 12% are discarded before reaching the final consumer and approximately 50% of infested units is discarded by the consumer (Appendix D). Furthermore, 1.0% (median; 90% CR, 0.2%–1.9%) of larvae are expected to survive to adulthood and escape from commercial waste while a median of 5.2% (90% CR 0.98%–12.2%) escape from consumer household waste. When the resulting numbers of adults emerge across NUTS2 regions the likelihood that a female will find a mate depends on the window of encounter in space and time. In combination with the likelihood that the subsequent progeny survives to initiate a founder population, the number of established founder populations was estimated to be 0.00014 per year (90% CR 0.00000–0.00264). Thus, the Panel would not expect new founder populations within the foreseeable future or the time horizon of 5 years of this assessment. The predicted waiting time between new founder populations is in the Panel's estimation at least approximately 380 years assuming an EI of 15 required for establishment (Table 6: 5 percentile) and 622 years (Table 7: 5 percentile) when an EI of 30 is assumed to be required for successful establishment. Table 9 gives quantiles of the average number of years between subsequent founder populations in different scenarios. Given the low frequency of new founder populations across all scenarios, the Panel did not proceed to quantitatively assess effectiveness of risk reduction options targeting L. orbonalis. However, such options exist, e.g. the production of eggplant and turkey berry fruit in pest‐free places of production. The Panel found four notifications mentioning that product infested with L. orbonalis was granted entry to the EU. This should no longer be the case after the instatement of temporary emergency measures against L. orbonalis in October 2022.
TABLE 9.
Range of estimates in expected number of years between establishment of founder populations according to scenario assumptions.
| Scenario | % of turkey berry: % eggplant | Threshold EI | Expected number of years between establishment of founder populations | ||||
|---|---|---|---|---|---|---|---|
| Percentile (%) | |||||||
| 5 | 25 | 50 | 75 | 95 | |||
| 1 | 25:75 | 15 | 379 | 1782 | 7006 | 35,590 | 355,600 |
| 2 | 100:0 | 15 | 144 | 686 | 2709 | 13,957 | 137,574 |
| 3 | 0:100 | 15 | 836 | 4007 | 15,845 | 81,222 | 831,525 |
| 4 | 25: 75 | 30 | 622 | 2923 | 11,493 | 58,380 | 583,316 |
| 5 | 100:0 | 30 | 236 | 1125 | 4444 | 22,895 | 225,673 |
| 6 | 0:100 | 30 | 1371 | 6573 | 25,991 | 133,235 | 1,364,012 |
Scenario 2 presents the greatest likelihood of L. orbonalis establishing in the EU although the median estimate is that a founder population would occur once in 2709 years (CR once in 144–137,574 years). However, this assumes only turkey berry is imported, which the Panel knows is not the case. The frequency of founder populations being introduced will be less than in scenario 2. The ratio turkey berry: eggplant of 25:75 (indicated with grey shading in the table and as scenario 4 in table 9) is considered the most plausible among the three tested compositions of the trade; the other two scenarios are extreme assumptions.
3.3. Spread
3.3.1. Assessment of spread
The spread of a species introduced into a new environment (provided that it finds an ecological niche to occupy) is characterised by a lag phase, a spread phase and a saturation phase (Figure 4). The lag period is the time from the first introduction and reproduction of the pest, i.e. the initiation of a founder population, to its establishment with a constant rate of range expansion into pest‐free areas.
To make an informed estimate of the presumed spread rate of L. orbonalis if it were to establish in suitable regions of the EU, the Panel investigated occurrence records of the related African species L. laisalis. This species was first reported in Europe from southern Spain in the 1950s and has been spreading in the southern Iberian Peninsula since then (Appendix E, Spread). L. orbonalis and L. laisalis are closely related (Mally et al., 2015) and share many similar life traits: They are native in the tropics and subtropics, the larvae of both species feed on Solanum spp. and the adults are of similar size, indicating similar flight‐ and therefore spread capacities.
The duration of the lag period in the regions where L. orbonalis could potentially establish was estimated to be approximately 34.5 years (90% CR 4.8–92.2 years). After the lag period, L. orbonalis is estimated to spread at a rate of 2.3 km/year (90% CR 0.6–7.0 km/year). More details are available in Appendix E (Spread).
3.3.2. Uncertainties affecting the assessment of spread
The African L. laisalis has different larval host plant preferences, with Solanum sodomeum (a synonym of Solanum linnaeanum) being its preferred host (Huertas‐Dionisio, 2000), although it has also been found to feed on S. melongena in Spain (Huertas‐Dionisio, 2000). S. linnaeanum appears to be of no agricultural interest, and its spatial availability will impact the spread rate of L. laisalis. Leucinodes orbonalis with its preferred host plant S. melongena – a widely cultivated crop in southern Spain – might therefore spread more easily, as its host plant is presumably much more abundant.
No studies were found investigating the flight capability of Leucinodes moths; Chang et al. (2014) state that the adults ‘only fly for short distances’.
It is unclear whether the spread of L. laisalis in the southern Iberian Peninsula originates from a single founder population, or whether spread from multiple founder populations.
Based on the occurrence records of L. laisalis, a mean spread rate of 1.6 km/year was calculated; in contrast, the second occurrence record in 1975 is at a distance of 196 km from the first record in 1958, resulting in a much higher spread rate of 11.5 km/year.
The duration of the lag period is mainly driven by the effect of EU agricultural practices and by the presence of natural enemies and control measures targeted at other Lepidopteran species, e.g. the Tomato leaf miner Tuta absoluta (Meyrick) (Gelechiidae), the potato tuber moth Phthorimaea operculella (Zeller) (Gelechiidae), and the Beet armyworm Spodoptera exigua (Hübner) (Noctuidae). These factors were not evaluated but are sources of uncertainty.
The expansion rate is driven by the dispersal ability of the insect, which is not well known, and by the effect of the host species communities in terms of species composition, patchiness and distance among suitable patches and availability in the EU environments compared to the observations collected from the area of origin. Information is lacking to assess this in detail.
3.3.3. Conclusions on spread
Were L. orbonalis to be introduced into the EU, the Panel estimates that it would take between 4.9 and 92.2 years (90% CR; median 34.5 years) for populations to grow sufficiently before a steady rate of spread of approximately 2.28 km/year (90% CR 0.65–7.02 km/year) was reached.
3.4. Impact
Leucinodes orbonalis larvae are oligophagous and feed on different species of Solanum. Its preferred host plant is eggplant, S. melongena, but it can also feed on other Solanum plants of economic relevance, such as tomato (S. lycopersicum) and potato (S. tuberosum). In tomato and potato, the larvae would mostly be confined to stem‐boring, as tomato fruit are reported as suboptimal plant tissue for feeding due to their high‐water content, and potato tubers are not fed on under natural conditions. With tubers developing underground, the larvae do not access the tubers.
3.4.1. Assessment of impact
Literature on impact of L. orbonalis in Asia is heavily skewed towards eggplant, the pest's primary host plant, with 800+ studies focussing on L. orbonalis feeding on this crop; in contrast, only very few papers with actual observations of L. orbonalis feeding on potato and tomato exist, with six papers and a single paper, respectively (see Appendix F.5).
In India, the cultivation area of eggplant in 2013 was 0.53 million hectares (Indian Council of Agricultural Research, 2016), whereas that of tomato amounts to 0.79 million hectares (Indian Council of Agricultural Research, undated). The area of potato cultivation was more than three times larger than that of eggplant, with 1.79 million hectares averaged over the years 2007–2009 (Scott & Suarez, 2011). These large cultivation areas of both tomato and potato in India vs. the low and infrequent infestations reported in the literature indicate that the expected impact on tomato and potato crops in the EU is similarly negligible. Nevertheless, the lack of data on yield losses in potato and tomato in the literature makes an assessment on the potential losses in the EU unfeasible at the moment.
The main impact of L. orbonalis on solanaceous crops in the EU is to be expected on eggplant. In India and Bangladesh, baseline infestation rates are 25%–30%, often due to poor agricultural sanitation where plant parts infested with L. orbonalis larvae are not immediately destroyed but instead collected at a corner of the field, enabling re‐infestation of the crop from adults emerging from the discarded plant parts.
Literature reporting eggplant yield losses in Asia was generally from regions with a CLIMEX Environmental Index (EI) > 30. Impacts in the EU were therefore considered to be largely limited to regions where EI > 30. Such locations coincide with the area for establishment, hence transient populations outside regions where EI < 30 were judged not to be able to cause measurable impacts. In southern European countries with EI > 30, the Panel estimates that approximately 4.5% (90% CR 2%–13%) of eggplant fruit grown outdoors may be lost to L. orbonalis infestation when no specific pest control measures are applied (see Appendix F for details and reasoning). In a scenario with specific pest control measures, the yield loss of eggplant was estimated to range from 0.28% to 1.9% (90% CR; median 0.54%).
3.4.2. Uncertainties affecting the assessment of impact
The main uncertainties affecting the impact assessment are related to the transferability of reports from Asia on impacts caused by L. orbonalis to the situation in the EU. Reports from Asia are predominantly on eggplant, which is grown in substantial quantities in Spain and Italy, the main sources of eggplant consumed within the EU.
The future likely loss of registered chemical insecticides in the EU and the increasing resistance of L. orbonalis against pesticides could constrain the effectiveness of control measures.
The development of non‐chemical alternatives (e.g. mating disruption, mass trapping and especially conservation biological control with generalist predators) could potentially improve control.
Climate change would be expected to influence the pest cycle, with higher temperatures increasing the population growth and the number of generations per year. How climate change will affect EU production of S. melongena is unknown.
3.4.3. Conclusions on impact
Were L. orbonalis to enter, establish and spread within the EU with the population reaching an approximate equilibrium such that EU farmers consider the organism a member of the general pest fauna, estimated median eggplant yield losses are estimated to be 4.47% (90% CR 0.67%–13.0%) when no specific control measures are in place, and 0.54% (90% CR 0.13%–1.94%) when growers apply targeted pest control against L. orbonalis.
3.5. Risk reduction options
Recognising the very low likelihood of establishment in the EU (0.00014 founder populations per year, 90% CR 0.00000–0.00264 per year), the Panel did not quantitatively evaluate the effectiveness of additional specific phytosanitary measures targeted at L. orbonalis. However, options for risk reduction exist, e.g. the production of eggplant and turkey berry fruit in pest‐free places of production in the countries of origin, and designation of L. orbonalis as a quarantine species. The Panel found four notifications mentioning that product infested with L. orbonalis was granted entry to the EU. L. orbonalis has a temporary EU emergency quarantine status since October 2022 (Article 30(1) of Regulation (EU) 2016/2031), which is an effective measure to stop this practice.
3.6. Consequences of climate change
The Panel did not conduct quantitative climate change scenarios for L. orbonalis, but considers that in the baseline scenario (EI ≥ 30; 25% turkey berry and 75% eggplant), 23% of the imported infested eggplant and turkey berry would be moved to areas suitable for establishment. With climate warming, the area suitable for establishment would enlarge, and hence, a greater proportion of the imported infested fruit would end up in areas suitable for establishment. If the number of infested fruits imported to areas suitable for establishment would increase by a certain percentage, then the number of moths emerging in those areas, the number of moths finding a mate, and the number of population founding events per year would increase by the same percentage in the pathway model used in this opinion. The average waiting time until a population founding event would then be decreased by this percentage. Based on this reasoning, the Panel concludes that climate warming would not qualitatively change the conclusions of the analysis under the assumption of current climate. Quantitatively, the risk area would be larger, and the time until the next founding event would be shorter, but founding events would still remain rare. In a worst‐case scenario where 100% of imports were distributed across areas with suitable climate, the frequency of founder populations would increase more, approximately four‐ to sevenfold (100/23 to 100/14). A sevenfold increase in likelihood of introduction for the baseline scenario (Table 7, Scenario 4) would give a median frequency of a founder population establishing in approximately 1600 years (90% CR 90–83,000 years).
3.7. Overall uncertainty
Entry pathways
This assessment focused on special and exotic varieties of eggplant (S. melongena) and on turkey berry (S. torvum) as pathways, imported from different tropical and subtropical Asian countries. Both commodities are part of a niche market and enter the EU in small, but largely uncertain quantities. The majority of eggplant consumed in the EU is produced within the EU, primarily in Spain and Italy. A wide range of varieties is grown in the EU, and niche products such as mini‐eggplants and white eggplants are added to this spectrum. Fruit of other Solanum plants may become popular and imported in larger quantities. This may change the composition and relative importance of pathways over time.
Transfer & establishment of founder populations
The imported consignments of eggplant and turkey berry from the countries where the pest occurs are generally small and in the order of a few kg to tens of kg per consignment (Appendix C). These consignments are too small to be further divided and distributed in the EU but will instead go as a whole to the customer. Where in the EU these final customers are is unknown, and the Panel therefore made the assumption that imported eggplants and turkey berry are distributed according to population in the EU (EFSA PLH Panel, 2021 Elasmopalpus lignosellus Quantitative Pest Risk Assessment) – an assumption that may be incorrect. However, the EFSA PLH Panel has no other simple and efficient basis on which to distribute imported produce that will aid the identification of points for pest introduction.
There is no specific information on production of waste of aubergines and turkey berry pre‐consumer (during distribution and retail) and by the consumer. The pathway model assumes that 10% of the product is turned into waste in the supply chain pre‐consumer, in line with an earlier assessment for Elasmopalpus lignosellus in asparagus for which empirical estimates were available from an American study (EFSA PLH Panel, 2021 Elasmopalpus lignosellus Quantitative Pest Risk Assessment; EFSA PLH Panel, 2023b). Waste by the consumer was based on expert knowledge elicitation, with a wide distribution of uncertainty. Escape from waste was also assessed by EKE, taking into account the experience gained during the earlier opinion on Taumatotibia leucotreta in cut roses, for which the escape from waste was analysed in detail (EFSA PLH Panel, 2023a Taumatotibia leucotreta Quantitative Pest Risk Assessment). Assessments of the escape probability of adult moths from discarded waste were made without the support from pest‐ or commodity‐specific empirical data, which contribute to wide ranges reflecting high uncertainty on the true values. There are no empirical data on the establishment of new persisting founder populations by single mated female moths, and this negatively affects the certainty of the estimates.
Climatic modelling of establishment
Establishment modelling results in maps of relative likelihood of establishment, and thresholds such as the threshold of EI ≥ 30 for establishment in the CLIMEX model should not be taken absolutely. In reality, there is an increase in suitability for establishment, number of generations and likely level of impact as EI increases. At which EI population densities are high enough for the insect to establish or to cause impact, is unknown. The threshold EI ≥ 30 is recommended in the CLIMEX documentation as a practically useful guide (Kritikos et al., 2015), but it cannot be used to distinguish areas where the insect can and cannot establish. Degrees of establishment potential exist. Maps can also not be translated 1:1 in expected population densities, but zones with higher EI are more likely to have higher population densities and higher impacts than zones with lower EI. Cross continents, zones with similar EI are expected to have similar pest pressure if the organism establishes outside the native range, but other factors than those accounted for in the model may affect pest pressure, e.g. natural enemies and cultivation practices. The Panel considered a threshold EI ≥ 15 to account for uncertainty about the precise value of the threshold.
Spread
Estimates of lag phase and constant rate of range expansion were made based on the spread of a related species, Leucinodes laisalis, in the southern Iberian Peninsula. It is not known whether the spread of L. laisalis was from one or multiple founder populations, which affects the certainty of the estimates.
Impact
Many studies have investigated the impact of L. orbonalis, especially in India. The majority of studies expressed impact as infestation rate (number of shoots and/or fruit affected), but yield loss is more related to severity than infestation rate. This requires estimation of yield losses and causes uncertainty.
Studies on impact are done under conditions that are conducive to impact to increase the power of comparing treatments in experiments, but this reduces the representativeness of the resulting data. Experiments that result in low impact may never be published because treatment comparisons would likely be inconclusive. Again, this lowers the representativeness of the studies.
Uncertainty decomposition
The decomposition of uncertainty with the pathway model (Table 10) indicates that the largest uncertainty is within the estimate of the amount of infested produce at the origin of the pathway (51% of model uncertainty). The level of pest infestation is often the largest uncertainty in quantitative pest risk assessments (e.g. EFSA PLH Panel, 2016a, 2016b, 2017a, 2017b, 2023). The next largest uncertainty in the model is the estimate of likelihood that larvae would escape from consumer waste and complete their development (15.0%), followed by the probability that eggs oviposited in the EU survive, hatch and the resulting larvae develop and complete development to reproduce and initiate a founder population (13%). Combining the factors involved in transfer, 45% of the model uncertainty is due to lack of information about transfer which is an area of invasion biology that typically lacks empirical evidence on the detailed steps involved (Leung et al., 2004; Simberloff, 2009).
TABLE 10.
Decomposition of explained variance in the pathway model for introduction of Leucinodes orbonalis. R2 gives the partial R2 of each regressor in a linear regression meta‐model of pathway model results in which the number of founder populations is the response variable and the parameter values in the model are regressors. The final column indicates the relative contribution of each parameter to explained variance. Here, variance represents the uncertainty in pathway model calculations, and the contribution of each parameter is the contribution to uncertainty.
| Rank | Model parameter | R 2 decomposition | Relative sensitivity (%) |
|---|---|---|---|
| #1 | Infestation rate on pathway on arrival into EU | 0.24 | 51 |
| #2 | Escape from consumer waste | 0.07 | 15 |
| #3 | Survival and development of eggs to initiate founder population | 0.06 | 13 |
| #4 | Mating in EU | 0.06 | 13 |
| #5 | % discarded by consumers | 0.02 | 4 |
| #6 | Quantity of imports | 0.01 | 2 |
| #7 | Weight of turkey berry cluster | 0.01 | 1 |
| #8 | Weight of exotic eggplant varieties | 0.00 | 1 |
| #9 | % discarded by trade | 0.00 | 0 |
| Sum | 0.46 | 100 |
4. CONCLUSIONS
Following a request from the European Commission, the EFSA Panel on Plant Health performed a pest risk assessment of Leucinodes orbonalis for the EU. The quantitative assessment focused on pathways and likelihood of entry, climatic conditions allowing establishment, the distribution of imported material within the EU after entry, the likelihood of establishment, the rate of spread following a lag period and potential impacts to Solanaceous crops in the EU.
L. orbonalis is an oligophagous pest feeding mainly on Solanaceae family crops (such as eggplants, potatoes and tomatoes). The main pathway (S. melongena and S. torvum from Asian countries where L. orbonalis presence is recorded) was deduced from the potential combinations between crops and countries of the origin. L. orbonalis is not known to occur outside of Asia and Oceania and the majority of records in Africa is most probably invalid. This species has been intercepted in the EU over 300 times since 2004. Based on the size and frequency of imports, and with evidence of interceptions in the EU, the interceptions were mainly on S. torvum and S. melongena fruits. The CN code for eggplants HS 070930 unifies the two above mentioned species in the category called ‘eggplants’ or ‘aubergines’.
The import data for those goods were downloaded from the Eurostat database for the years 2010–2019 from the Asian countries with the records of L. orbonalis extracted from the scientific literature. Based on the size and frequency of imports, and with evidence of interceptions in Europe, the importation of eggplants from Asian countries was identified as the most likely pathways for entry. Tomato and potato are hosts of L. orbonalis, but were not considered as pathways as the larvae cannot develop to maturity in tomato fruit while import of potato from Asia is forbidden and the insects do not enter the tuber under field conditions.
Climate matching and CLIMEX modelling indicate that conditions are most suitable for establishment of L. orbonalis in parts of the southern EU, especially around the Mediterranean Sea. Two possible scenarios for establishment were considered based on two EI thresholds. Using EI ≥ 15 approximately 23% of imported S. melongena and S. torvum are distributed to NUTS2 regions in which climatic conditions are suitable for establishment. Using EI ≥ 30 approximately 14% of imports reach such areas.
Each infested eggplant entering the EU is likely to contain only one larva, as such an important limiting factor in establishing a founder population is the likelihood of a male and a female emerging in temporal and spatial proximity to locate each other and mate. With respect to the need of larval development to adulthood from discarded infested produce, then mating and the progeny surviving, the number of newly established founder populations developing was estimated to be 0.00014 per year (90% CR 0.00000–0.00264). Thus, the Panel would not expect new founder populations within the foreseeable future. Nevertheless, if a founder population were to establish, it would likely remain local for a number of years and the lag period before sustained spread was estimated to be 34.5 years (90% CR 5–92 years) following the establishment of a founder population. L. orbonalis is not considered to be a strong flyer. Were L. orbonalis to establish, the median rate of natural spread was estimated to be 2.3 km/year (90% CR 0.65–7.0 km per year).
Impact assessment focused on Solanaceae host plant species, mainly eggplants. In a scenario where L. orbonalis has spread and is managed by farmers as part of the general pest fauna, i.e. no specific official phytosanitary measures are in place against it, and growers apply targeted pest control against L. orbonalis, median yield losses in eggplant were estimated to be 0.54% (90% CR 0.13%–1.94%). The Panel found insufficient evidence to consider tomato and potato production to be at risk from infestation by L. orbonalis if the insect were to establish in the EU because tomato and potato are unpreferred hosts. In the presence of eggplant, L. orbonalis will feed on eggplant. The Panel did not consider the possibility of shifts to previously unreported host plants because it falls outside the scope of a PRA (EFSA PLH Panel, 2018).
Concluding overall, this opinion shows that the EU encompasses regions with climate suitable for the establishment of L. orbonalis, and that this species could cause damage if it established. However, it is unlikely to be introduced in the foreseeable future because of the relatively low volume of commodity providing a pathway, and the low likelihood that adults emerging in the EU will successfully mate and initiate a founder population.
ABBREVIATIONS
- CN
Combined nomenclature (8‐digit code building on HS codes to provide greater resolution)
- CR
certainty range
- DD
degree days
- DNA
Deoxyribonucleic acid
- EI
ecoclimatic index (an index of climatic suitability used by CLIMEX)
- EKE
Expert Knowledge Elicitation
- EPPO
European and Mediterranean Plant Protection Organization
- HRP
High Risk Plants
- HS
Harmonised System (6‐digit World Customs Organization system to categorise goods)
- IPM
Integrated Pest Management
- ISPM
International Standard for Phytosanitary Measures
- MS
Member state (of the EU)
- NUTS
Nomenclature Units for Territorial Statistics
- RRO
risk reduction option
- SDM
Species Distribution Model
- ToR
Terms of Reference
CONFLICT OF INTEREST
If you wish to access the declaration of interests of any expert contributing to an EFSA scientific assessment, please contact interestmanagement@efsa.europa.eu.
REQUESTOR
European Commission
QUESTION NUMBER
EFSA‐Q‐2023‐00069
PANEL MEMBERS
Claude Bragard, Paula Baptista, Elisavet Chatzivassiliou, Francesco Di Serio, Paolo Gonthier, Josep Anton Jaques Miret, Annemarie Fejer Justesen, Alan MacLeod, Christer Sven Magnusson, Panagiotis Milonas, Juan A. Navas‐Cortes, Stephen Parnell, Roel Potting, Philippe L. Reignault, Emilio Stefani, Hans‐Hermann Thulke, Wopke Van der Werf, Antonio Vicent, Jonathan Yuen, and Lucia Zappalà.
COPYRIGHT FOR NON‐EFSA CONTENT
EFSA may include images or other content for which it does not hold copyright. In such cases, EFSA indicates the copyright holder and users should seek permission to reproduce the content from the original source.
MAP DISCLAIMER
The designations employed and the presentation of material on any maps included in this scientific output do not imply the expression of any opinion whatsoever on the part of the European Food Safety Authority concerning the legal status of any country, territory, city or area or of its authorities, or concerning the delimitation of its frontiers or boundaries.
Supporting information
Supplementary material to the 'Pest risk assessment of Leucinodes orbonalis for the European Union' ‐ Entry model
ACKNOWLEDGEMENTS
The EFSA PLH Panel wishes to thank the following for the support provided to this scientific output: Irene Pilar Munoz Guajardo and Marika De Santis from the EFSA library for providing an easy access to the crucial references; ISA Expert: Ivana Majić (University of Osijek, Faculty of Agrobiotechnical Sciences) for the first draft of the Expert Knowledge Elicitation evidence dossier for the Spread and Impact session. The EFSA PLH Panel wishes to thank Hearing Experts: Cherubino Leonardi (University of Catania, Italy), Aleksandar Jovanovic (Export Marketing Consultant in ‘Autentika’), Srinivasan Ramasamy (the Flagship Program Leader for Safe and Sustainable Value Chains and Lead Entomologist of the World Vegetable Center (Taiwan), Luke Chinaru Nwsou (University of Port Harcourt, Nigeria).
APPENDIX A. Identity of Asian Leucinodes
A.1.
There is the possibility of currently undiscovered Asian Leucinodes species similar to the situation uncovered by Mally et al. (2015) in Sub‐Saharan Africa. The vast majority of reviewed Asian literature does not report any efforts or methods of identifying the investigated specimens to species. In order to investigate this possibility of undiscovered Leucinodes species, the available literature for published DNA sequence data were screened, that could help answering this question. A part of the mitochondrial cytochrome oxidase I (COI) gene was proposed to serve as ‘DNA Barcode’ to identify animal species based on their unique group of DNA Barcode sequences (Hebert et al., 2003). Therefore, ‘DNA Barcode’ sequences were compiled from several publications on the Austral‐Asian L. orbonalis (Chang et al., 2014; Mally et al., 2015; Sagarbarria et al., 2018; Shashank et al., 2015; Murali et al., 2017; Palraju et al., 2018; Karthika et al., 2019; Natikar et al., 2022; Padwal et al., 2022), DNA Barcode sequences available in the Barcode of Life Database and sequences not published in the literature but submitted as ‘unpublished’ to GenBank. We assembled a total of 398 sequences into a DNA alignment (Data S1) and used the R packages APE (Paradis et al., 2004) and SPIDER (Brown et al., 2012) as well as the FigTree v1.4.4 software (Rambaut, 2006–2018) to analyse the data and illustrate a neighbour‐joining tree (Figure A.1).
FIGURE A.1.

Neighbour‐joining tree of Austral‐Asian Leucinodes partial cytochrome oxidase I (COI) sequences. The coloured groups represent putative undescribed species from Vietnam (in blue) and Australia (in red and orange); the large remainder of sequences (marked in black) refers to L. orbonalis and exclusively originates from Asia. The scale bar represents 0.02 nucleotide substitutions per site (Creator and Copyright©: R. Mally, R packages ape and spider, FigTree v1.4.4).
In the Barcode of Life Database (https://v4.boldsystems.org), L. orbonalis is represented by the Barcode BIN BOLD:AAE7334. However, based on examination of DNA sequences of the mitochondrial COI gene, there appear to be three potentially undescribed Leucinodes species: one from Hanoi (Vietnam) (Barcode BIN BOLD:ACH6349; Chang et al., 2014), and two from Queensland (Australia; Barcode BINs BOLD:AAO2620, BOLD:AAO9130), potentially occurring sympatrically with L. orbonalis.
APPENDIX B. Biology of Leucinodes orbonalis
B.1.
Leucinodes orbonalis is a species of Lepidoptera. The life cycle comprises an egg stage, six larval stages, a pupal stage and an adult stage.
Biology. The biology of L. orbonalis has been described in more detail from populations in India (Lall & Ahmad, 1965; Saxena, 1965), the Philippines (Ardez et al., 2008; Navasero & Calilung, 1990) and China (Li et al., 2014). The pre‐imaginal development of L. orbonalis is temperature‐dependent (Koundinya et al., 2019; Li et al., 2014; Mannan et al., 2015).
Egg. Eggs are usually laid in four subsequent nights, with 48–53 eggs per night totalling to 192–212 eggs (Saxena, 1965); in the Philippines, Navasero and Calilung (1990) found the females to lay 8–295 eggs, with an average fecundity of 118 eggs per female. Eggs are laid during night, singly or in small clusters of two to four on the lower leaf surfaces of the topmost and middle leaves of S. melongena (Ardez et al., 2008); under lab conditions, eggs are laid in larger egg masses of 40–60 eggs (Mannan et al., 2015). The eggs are oval and creamy white, turning orange towards larval hatching, with the black head capsule becoming visible through the eggshell (Ardez et al., 2008; Lall & Ahmad, 1965). The diameter of a single egg is on average 0.48 mm (Lall & Ahmad, 1965). Egg development time is inversely correlated with temperature, with higher temperatures shortening the time for development (Li et al., 2014). Larval hatching tends to occur primarily in early mornings (Mannan et al., 2015) and late afternoons (Li et al., 2014).
Larva. Freshly hatched larvae wander around on their host plant in search of a suitable place to penetrate the plant tissue; this searching lasts from a few minutes (Das & Patnaik, 1971) to about an hour (Saxena, 1965). The entry hole into the fruit/shoot is plugged with excreta, presumably to keep the plant part from drying out, and to keep pathogens, parasitoids and predators out. However, Navasero and Calilung (1990) report that the entry holes and excreta facilitate the development of secondary infections by microorganisms and insect scavengers (Staphylinidae, Phoridae). The entry holes may also partly heal while the fruit is further growing in size (Navasero & Calilung, 1990). Usually, a single fruit is inhabited by one or more rarely two larvae, but sometimes up to four larvae can be found in one fruit (Akulov et al., 2015; Shukla, 1986).
While Saxena (1965) states that the larva does not leave the inside of the plant shoot or fruit until it starts building the cocoon for pupation, Navasero and Calilung (1990) assume that larvae from dried‐up, fallen leaves, flowers and shoots climb back up the plant and infest other plant parts, which is evidenced by exit holes (which lacked a plug of excreta) and entry holes larger than those made by first‐ and second‐instar larvae. According to Li et al. (2014), the larvae feed in as many shoots as necessary for development, but once they enter fruit, they remain inside until mature enough for pupation. In host plants such as Solanum nigrum with fruits too small to harbour the growing larva, it exits the fruit and webs together three to four fruits to feed on them externally from inside the web (Das & Patnaik, 1971).
Information about larvae feeding on leaves are contradictory: Saxena (1965) expressed doubts about earlier reports of the caterpillar boring into the leaves' petioles and mid‐ribs (Banerjee & Basu, 1955), ‘because the entry of the caterpillar inside the plant tissue brings about a withering effect on the tender parts of plants which dry up and fall down’. Li et al. (2014) also state that L. orbonalis does not damage the leaves of its host plant. However, Navasero and Calilung (1990) confirmed that larvae bore into the petioles and mid‐ribs, causing the leaves to dry and eventually fall off. Especially the young instars feed on the petals of ovaries of flowers, causing the flower to fall off or to develop into an abnormally shaped fruit that may or may not reach maturity (Li et al., 2014; Navasero & Calilung, 1990).
When feeding in shoots, the larva enters this part of the host plant through tender parts, e.g. where the leaf buds are located. Stem‐boring leads to the drooping of the topmost leaves due to the disruption of the water‐conducting tissues in the stem. Fruits are entered directly and not through the shoots (Saxena, 1965), and apparently usually from below the calyx where the neonate larvae hide (Das & Patnaik, 1971; Akulov et al., 2015). When fruits are present on the host plant, they are preferred by the larvae over the plant's shoots (Navasero & Calilung, 1990), potentially due to the shoots becoming tough and hard in older plants (Lall & Ahmad, 1965).
From the fifth instar on, the caterpillar feeds less voraciously than in earlier instars (Saxena, 1965). If disturbed in the fourth instar, or if the fruit starts to rot, the caterpillar will construct a silken cocoon and for the remainder of its larval development stay in there, passing the remaining instars and eventually pupation in there (‘cocooning behaviour’; Saxena, 1965). This has important implications for the potential accidental introduction of L. orbonalis to other parts of the world: The larva only requires a time window of about 15–16 days of undisturbed feeding from hatching from the egg to the fourth larval instar in order to get to a stage of development where no more food is required and the life cycle can be completed. This cocooning behaviour, with the larva residing in its pupation cocoon from the fifth or sixth larval instar on, might explain why some authors (e.g. Atwal, 1976; Boopal et al., 2013; Mathur & Jain, 2006) report only four or five larval instars.
Three days after the fifth moult, the larva leaves the fruit/shoot through a relatively large exit hole to find a suitable place on the lower branches of the plant among dry leaves for pupation (Saxena, 1965).
In tropical climate, the development of second‐ to sixth‐instar larvae is faster compared to that in subtropical climate (Figure B.1). Egg, pre‐pupal and pupal stages, on the other hand, appear to hardly vary in their duration between different climates (Figure B.1).
FIGURE B.1.
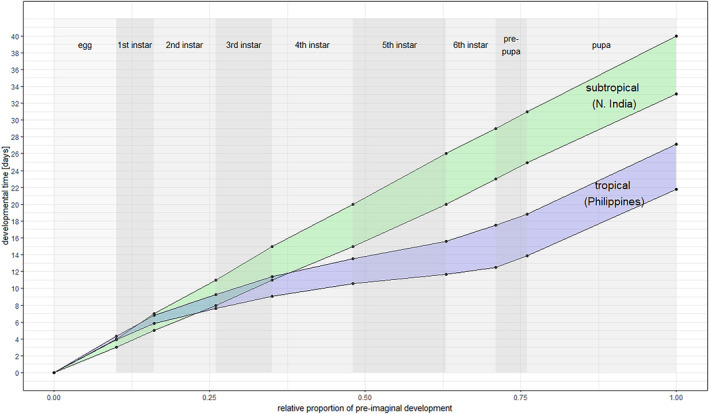
Relative (x‐axis) and absolute (y‐axis) duration of the phases of development of L. orbonalis from egg to pupa for Northern India (green area, from Saxena 1965) and the Philippines (blue area, from Navasero & Calilung, 1990); the shaded areas between the point lines indicate the range between minimum and maximum developmental time for each stage. Baseline for relative durations of developmental phases based on Saxena (1965) (Creator and Copyright©: R. Mally, data from Table B.1, created with R package collection ‘tidyverse’).
TABLE B.1.
Pre‐imaginal stages, their durations and body measures; from (a): Saxena (1965; Patna, Bihar state, India), (b): Navasero and Calilung (1990; Los Baños, Laguna province, Philippines).
| Stage | Duration of instar (days) | Length | Width |
|---|---|---|---|
| Egg |
(a): 3 in summer, 3–4 days in winter (b): 4.12 ± 0.22 days |
(a): 0.8–0.85 mm (b): 0.82 mm |
(a): 0.5–0.6 mm (b): 0.59 mm |
| 1st larval |
(a): 2–3 days (b): 2.19 ± 0.25 days |
(a): 0.7–3.2 mm (b): 1.3 ± 0.094 mm |
(a): 0.2–0.5 mm (b): 0.26 ± 0.02 mm |
| 2nd larval |
(a): 3–4 days (b): 2.16 ± 0.35 days |
(a): 4.5–6.2 mm (b): 2.92 ± 0.15 mm |
(a): 0.8–1.3 mm (b): 0.41 ± 0.05 mm |
| 3rd larval |
(a): not stated (b): 1.77 ± 0.34 days |
(a): 6.5–8.7 mm (b): 5.32 ± 0.61 mm |
(a): 1.4–1.7 mm (b): 0.61 ± 0.05 mm |
| 4th larval |
(a): 4–5 days (b): 1.8 ± 0.33 days |
(a): 9.3 mm (b): 8.91–9.79 mm |
(a): 1.8 mm (b): 0.93–1.19 mm |
| 5th larval |
(a): 5–6 days (b): 1.61 ± 0.45 days |
(a): 16.8 mm (b): 13.41 ± 0.82 mm |
(a): 2.9 mm (b): 1.8 ± 0.2 mm |
| 6th larval |
(a): 3 days (b): 1.36 ± 0.54 days |
(a): 19.3 mm (b): 18.2 ± 1.51 mm |
(a): 3.6 mm (b): 3.62 ± 0.37 mm |
| Prepupa |
(a): 46–48 h (b): 1.34 days |
(a): 11.3 mm (b): Not stated |
(a): Not stated (b): Not stated |
| Pupa |
(a): 8–9 days, regardless of the season (b): 8.1 ± 0.2 days |
(a): 11.6 mm (b): 12.21 ± 0.63 mm |
(a): 3.8 mm (b): 4.2 ± 0.25 mm |
| Adult |
(a): ~ 7 days (females), 4–5 days (males) (b): 4–8 days (mean 5.84 days) |
(a): Not stated (b): 13 mm |
(a): 22 mm wingspan (b): 23.5 ± 0.5 mm wingspan |
Larval host plants. The species is part of the tribe Lineodini, a lineage of Crambidae moths that has its highest diversity in the American tropics, and that has specialised on Solanaceae (nightshades) as larval food plants (Mally et al., 2019). The preferred host plant is eggplant/aubergine (Solanum melongena), where usually 1–3 (but up to 20) larvae feed inside a single fruit, which primarily depends on the number of seeds in the fruit (the fewer seeds, the more larvae), but also on the fruit's size (personal communication dr S. Ramasamy). Despite numerous citations of non‐Solanaceae larval host plants in the literature, L. orbonalis appears to be unable to complete its life cycle on plants other than nightshades (Alam et al., 2003). Ardez et al. (2008) studied the host plant preference in no‐choice trials in the laboratory and found that L. orbonalis larvae were unable to develop on fruits of okra (Abelmoschus esculentus, Malvaceae) or tubers of sweet potato (Ipomoea batatas, Convolvulaceae). Larvae were feeding on cowpea fruits (Vigna unguiculata, Fabaceae), but formed abnormal pupae, from which no adults emerged (Ardez et al., 2008).
In free‐choice trials, L. orbonalis only laid eggs on Solanum melongena, but on none of the other six available plants (three Solanaceae: S. tuberosum, S. nigrum, S. lycopersicum, and three non‐Solanaceae: Vigna unguiculata, Abelmoschus esculentus, Ipomoea batatas), indicating that eggplant is the preferred larval host plant (Ardez et al., 2008). However, no‐choice trials showed that the life cycle can also be completed on potato tubers (S. tuberosum) and fruits of black nightshade (S. nigrum), although the moths reared from potato tubers had a shorter adult lifespan compared to moths reared from fruits of eggplant and black nightshade (Ardez et al., 2008). Under natural conditions (i.e. not in laboratory rearings), L. orbonalis larvae appear to only attack the shoots of potato but not the tubers (Nair, 1967; Natikar & Balikai, 2021). The larvae can successfully complete their life cycle in the shoots of potato plants (Navasero & Calilung, 1990), and the species has become the most harmful organism for potato crops in the Indian state of Karnataka (Natikar & Balikai, 2018a). In plants with fruits being too small for the growing caterpillar to effectively feed inside (as is the case e.g. in S. nigrum), the larva in later instars exits the fruit and spins several fruits together with a silken web, lives in the fruit cluster and feeds on the fruits externally (Das & Patnaik, 1971; Navasero & Calilung, 1990). Outside the Indian growing season, when eggplant is not planted, L. orbonalis uses the black nightshade, S. nigrum, and the yellow‐fruit nightshade, S. virginianum (= S. xanthocarpum) as larval host plants, feeding in shoots and fruits (Lall & Ahmad, 1965).
Ardez et al. (2008) also report that larvae successfully fed on tomato fruits (Solanum lycopersicum, Solanaceae) and reached the pupal stage, but as in cowpea, no adults emerged from the pupae. Contrary to the observation of Ardez et al. (2008), Das and Patnaik (1971) found the species boring into tomato fruits and shoots in the wild, and, like Boopal et al. (2013), successfully reared larvae to adults on tomato fruits in the lab. Lab‐reared L. orbonalis also successfully completed their life cycle in the study of Jethva and Vyas (2009), but it is not clear from the study on which part of the tomato plant (stem/fruits) the larvae were feeding. Due to the paucity of literature reporting L. orbonalis infestations on tomato in Asia as well as the absence of interceptions of L. orbonalis from tomato fruit imported from Asia, tomato is not considered as a pathway for entry (Table B.2).
TABLE B.2.
Leucinodes orbonalis larval host plants: summary of information from literature about biology.
| Hosts described in the literature | Number of references (refIDs) | Life stage of L. orbonalis | Where on the plant detected | Detailed description from the cited reference |
|---|---|---|---|---|
| Solanum aethiopicum (Ethiopian eggplant), gilo cultivar group (Scarlet eggplant); synonym S. integrifolium | 2 (Kariyanna et al., 2020a, Kariyanna et al., 2020b) | Egg, larva, pupa, adult | Shoots, fruits | |
| Solanum incanum × Solanum melongena F1 hybrid variety | 1 (Baksh, 1979) | Larva | Fruits | |
| Solanum lycopersicum (tomato) | 5 (Ardez et al., 2008; Boopal et al., 2013; Das & Patnaik, 1971; Jethva & Vyas, 2009; Maureal et al., 1982) | Egg, larva, pupa | Leaves, fruits |
‘Bores into tender shoots’ (Shrestha & Lee, 2020) ‘unclear where the eggs where laid, and on which plant tissue the larvae fed’ (Boopal et al., 2013) |
| Solanum macrocarpon (African eggplant) | 2 (Dhaliwal & Aggarwal, 2021; Hanur et al., 2011) | Larva, pupa (in the soil) | Fruits, shoots |
‘BSFB was able to infect both the shoot meristem tips as well as developing fruits, suggesting that S. macrocarpon is vulnerable to BSFB in all stages. The extent of susceptibility, however, was more in the latter case. Also, larvae of BSFB were found to search for the fruit rather than the tougher calyx region when challenged onto the fruits per se.’ (Hanur et al., 2011) ‘Larvae of L. orbonalis have an internal feeding habit, which results in withered shoots and destruction of fruit tissues.’ (Dhaliwal & Aggarwal, 2021) |
| Solanum melongena (eggplant, aubergine, brinjal) | 147 references | Egg, larva, pupa, adult | Flowers, shoots, stems, fruits, leaves |
‘The pest overwinters in larval stage under a hardy, leathery and dirty coloured cocoon attached to the host plant usually at about 1–3 cm below the soil surface’ (Lal, 1975) ‘larvae bore into tender shoots during vegetative growth, subsequently attacking flowers and fruits’ (Islam et al., 2020) ‘SFB larvae enter the internal tissue of the plant and become inaccessible to the external environment by sealing their entry points with their silk and excreta’ (Talukder et al., 2021) ‘Larvae grow to 2 cm in length with segmented legs, mandibles and the body is divided into three parts – head, thorax and abdomen. It moults 4 times and passes through 5 instars (Singla et al., 2018).’ (Talukder et al., 2021)’ ‘Female can lay up to 250 white‐coloured with 0.5 mm diameter and flat eggs singly on lower surface of leaves, shoots, flower buds and sometimes on calyces of fruits. Incubation period is 3–6 days; larva becomes mature in 10–15 days. Larva is stout, pinkish in colour with brown head (Plate 7.2L). Pupation takes place in greyish boat‐shaped tough cocoons (Plate 7.2 m) among fallen leaves or on plant itself. Pupal period lasts for 6–10 days, after which adult emerges. The whole life cycle from egg to adult emergence takes 21–45 days with up to five overlapping generations in a year’ (Kunjwal & Srivastava, 2018) ‘Being an internal borer the early instar larvae of this pest feed exclusively on the tender shoots, petiole and flower buds, while the later instars bore into the fruits’ (Nayak, 2014) ‘Third‐ and fourth‐instar ESFB larvae are most responsible for the damage to eggplant, according to reports. The larvae of the eggplant shoot and fruit borer eat the vulnerable shoots of infected plants’ (Islam et al., 2022) ‘Because the larvae hide in fruits and shoots, insecticides used to get rid of them are typically unsuccessful’ (Islam et al., 2022) ‘lay its eggs on the lower surface of fresh leaves in clusters of 80–253 eggs on average’ (Ullah et al., 2022) ‘Larvae also feed on flowers, which results in flower drop or misshapen fruits’ (Shelton et al., 2019) ‘Numbers of larvae per leaf ranged among the five compared cultivars from 0.27 to 0.43 larvae per leaf’ (Kassi et al., 2019) ‘Once the larva bores into petiole and midrib of leaves and tender shoots, it causes dead hearts. In later stages, it also bores into the flower and fruits’ (Singh et al., 2014) |
| Solanum nigrum (black nightshade) | 2 (Ardez et al., 2008; Das & Patnaik, 1971) | Egg, larva | Fruits, leaves |
‘The eggs of EFSB [...] reared on black nightshade which laid their eggs on the underside of the leaflets, hatched in 4 days. The EFSB eggs are usually laid singly or in a cluster of 2–4.’ (Ardez et al., 2008) ‘As the larvae grows, it cannot accommodate itself in the fruit of Solanum nigrum. Therefore, it comes out and webs 3–4 such fruits together and feeds on them by remaining inside the web.’ (Das & Patnaik, 1971) |
| Solanum sp. | 2 (Mannan et al., 2015; Vinod et al., 2001) | Larva | Fruits | |
| Solanum tuberosum (potato) | 11 (Lall & Ahmad, 1965; Nair, 1967; Maureal et al., 1982; Hanapur & Nandihalli, 2004; Ardez et al., 2008; Tribikram & Patnaik, 2010; Boopal et al., 2013; Kariyanna et al., 2020a; Kariyanna et al., 2020b; Natikar & Balikai, 2018b, 2022) | Egg, larva, pupa | Shoots, leaves, roots |
‘The larva of shoot borer, L. orbonalis, attacks the shoots of potato, causing withering and wilting of the stem ultimately resulting in retardation of the plant growth’ (Natikar & Balikai, 2022) ‘The newly hatched larvae entered the lamina of the leaf, fed on the parenchyma and then entered into the leaf stalk, as a result of which the leaf withered. It has been learnt (in litt.) from Dr. K. K. Nirula, Nematologist, Central Potato Research Institute, Shimla (Punjab) that L. orbonalis is found boring the shoots of potato in Bihar, but the incidence of the borer is very low.’ (Nair, 1967) ‘Up to 4 larvae were observed boring the shoots and leaf stalks on each plant of this variety [Alpha]’ (Nair, 1967) |
| Solanum viarum (tropical soda apple) | 1 (Udayagiri & Mohan, 1985) | Larva | Fruits | ‘The caterpillars were noticed feeding on the contents of the developing berries. The entry holes were plugged with excreta.’ (Udayagiri & Mohan, 1985) |
Pupa. The sixth‐instar larva (or fourth‐ to fifth‐instar larvae in the case of ‘cocooning behaviour’, see under ‘Larva’) leaves the plant part it was feeding in and searches for a dry place for pupation, usually near the stem base (Lal, 1975), or on twigs or leaves (Mathur & Jain, 2006), where it constructs a boat‐shaped pupation cocoon, leaving a small exit hole in the cocoon from which it will later emerge as an adult (Saxena, 1965). Construction of the cocoon takes about a day (Navasero & Calilung, 1990). The fresh cocoon is completely white and turns into a dark brown in the course of a few hours (Saxena, 1965), matching the surroundings and making it difficult to spot the cocoons (Alam et al., 2003). According to Ardez et al. (2008), the pupal cases' colour depends on the larval host plant, ranging from (light) brown (reared on eggplant) over light to yellowish brown (tomato) to pinkish brown (on black nightshade). The size of the cocoon is determined by the nutrition status of the larva (Patel & Basu, 1948; Saxena, 1965). The actual pupation is, as all other stages of the insect, temperature‐dependent (Alam et al., 2003; Mannan et al., 2015) and takes 8–9 days on average (Ardez et al., 2008; Lall & Ahmad, 1965; Navasero & Calilung, 1990; Saxena, 1965). The maculation of the adult becomes slightly visible about 2 days before the moth emerges (Navasero & Calilung, 1990).
Adult. After emergence form the pupal cocoon, the adult rests for about 15 min upon the cocoon in order to dry, and to pump haemolymph into the wings, thus completely unfolding them (Saxena, 1965). Adult emergence takes primarily place in the first part of the night (Alam et al., 2003; Mannan et al., 2015).
Female moths can be distinguished from males by their somewhat larger appearance due to a thicker abdomen and a larger wingspan. Furthermore, the female bents its thick abdomen upwards above the body in resting position (Figure B.2), releasing sex pheromones from the abdomen tip to attract males for mating (‘calling behaviour’), whereas males keep their slender abdomen straight at rest. The proboscis is well‐developed, but according to Saxena (1965) the adults do not appear to feed; nonetheless, in lab rearing studies, a sugary liquid is generally provided for the adults to feed on (Lall & Ahmad, 1965; Navasero & Calilung, 1990).
FIGURE B.2.

Adult Leucinodes orbonalis female in its typical resting position with the abdomen bent up, releasing sex pheromones from the abdomen tip to attract males for mating. Photo: © Md Jusri, under CC‐BY‐NC 4.0 licence (https://creativecommons.org/licenses/by‐nc/4.0/), iNaturalist observation https://www.inaturalist.org/observations/15009501.
The adults are purely nocturnal, with emergence from the pupal cocoon, mating and egg laying all taking place at night, while remaining inactive during the day, resting on the underside of the host plants' leaves (Mannan et al., 2015). They are attracted to light and can be caught and monitored using light traps (Yousafi et al., 2016a, 2016b).
The adult stage is short‐lived: approximately 3 days after its emergence form the pupal cocoon, and after mating, the female lays eggs for four consecutive nights and then dies; the males live for 4–5 days and die shortly after copulation (Saxena, 1965). Other sources state that mating occurs already in the night of emergence from the cocoon or the following night (Kavitha et al., 2008; Mannan et al., 2015). The entire copula lasts approximately 30–45 min (Mannan et al., 2015). Females usually mate more than once (Mathur & Jain, 2006). An artificial sex pheromone mimicking that of the females has been developed to mass‐trap males and disrupt mating (Cork et al., 2001; Gunawardena et al., 1989). Females produce their sex pheromone mainly during a time window from 10 p.m. to midnight, and male response is highest during the first night (Gunawardena et al., 1989), potentially due to the female still being virgin. The female sex pheromone has a maximum effective range of 10 m, and moths released at 15 or 20 m from a pheromone source did not reach the source in a mark‐and‐recapture study (Prasad et al., 2005). The female starts laying eggs in the night after the (first) copulation, although egg laying in the night of copulation has also been reported (Mannan et al., 2015 and references therein). The number of eggs laid decreases with each night, and the entire duration of oviposition is 2–4 days (Mannan et al., 2015 and references therein).
Generations per year. In tropical climate (Laguna province, the Philippines) under lab rearing conditions, the life cycle of L. orbonalis takes about 26–36 days (Navasero & Calilung, 1990), so that 10–14 generations per year can be completed. The species also reproduces year‐round under subtropical conditions in regions such as Bihar state (India), with the entire life cycle ranging from 39 days in January/February to 26 days in May, with an average of 30.61 days per generation (Lall & Ahmad, 1965). In such a climate, 11–12 generations per year can be completed. Jethva and Vyas (2009) extrapolated from lab studies of L. orbonalis reared on tomato that the population would multiply by a factor of 2.358 per week.
Hibernation. In the Kullu Valley of Himachal Pradesh, India, the species occurs up to the middle part of the valley (1200–2000 m above sea level), where cold winter temperatures (down to −6.5°C) and snow cover (up to 200 cm) require L. orbonalis to hibernate (Lal, 1975). Here, the species overwinters as fully grown larva in a cocoon attached to the base of the main shoot, just above the roots between 3 cm beneath to 1.5 cm above the soil. There are usually 1–2, and up to 5 cocoons per plant (Lal, 1975). In lower altitude regions of India, where hostplants are available year‐round, L. orbonalis supposedly does not hibernate (Lal, 1975).
Natural enemies. Srinivasan (2008) lists the lacewing Chrysopa kulingensis (Neuroptera: Chrysopidae), an unidentified species of the true bug genus Campyloneura (Hemiptera: Miridae) and three species of Coccinellidae (Cheilomenes sexmaculata, Coccinella septempunctata, Brumoides suturalis) as predators of L. orbonalis, presumably especially of the eggs and pupae, and Navasero and Calilung (1990) report an unidentified earwig (Dermaptera) present in about 15% of damaged fruits that likely preys on L. orbonalis larvae, as these were either missing or dead in those fruits. Fathi (2022) found three predatory insect species on eggplant in Northern Iran: Chrysoperla carnea (Neuroptera: Chrysopidae), Orius niger (Heteroptera: Anthocoridae) and Coccinella septempunctata.
Srinivasan (2008) lists six parasitic wasp species of Ichneumonidae, three of Braconidae, one Chalcididae and one Eulophidae species, as well as one species of Tachinidae (Diptera) as parasitoids of immature stages of L. orbonalis. In the Philippines, five species of parasitic wasps are reported by Navasero and Calilung (1990), with an unidentified Apanteles species (Braconidae) being the most abundant parasitoid. Parasitisation rates of L. orbonalis larvae by Apanteles sp. were found to be higher in shoots (11.11%) as compared to fruits (3.12%), presumably because host larvae can be detected and reached more easily by the parasitoid in shoots than in the more voluminous fruits (Navasero & Calilung, 1990). In Northern Iran, two main parasitoids are reported: Trichogramma brassicae (Hymenoptera: Trichogrammatidae) and Bracon hebetor (Hymenoptera: Braconidae) (Fathi, 2022).
The fungus Curvularia spicifera (Ascomycota: Pleosporaceae) is a plant pathogen but is reported (as synonym Bipolaris tetramera) as an L. orbonalis entomopathogen by Srinivasan (2008), citing Tripathi and Singh (1991).
APPENDIX C. Pest Introduction: Identification of main pathway
C.1.
To be considered a genuine host, a phytophagous insect must be able to complete its development and produce viable progeny following mating by feeding only on the plant regarded as a host. Such information is seldom found in literature, which often only summarises the information on pests and plants they are associated with. Reports of different life stages being found on the plant can often be a good indicator of a plant being a true host. Table C.1 lists the plant species on which L. orbonalis has been intercepted, as well as plants recorded in the literature and amount of information available on that (sometimes none, or scarce).
TABLE C.1.
Compilation of plant commodities with L. orbonalis interceptions (2nd column) and host plants reported in the literature (3rd column), including confirmed feeding (4th column) and confirmed completion of the life cycle (5th column). Potential pathways (plant species) of introduction requiring action are marked in bold, potential pathways without action required are marked with an asterisk.*
| Plant species | Number of interceptions on that commodity | Plant part imported – Any life stage associated? | Number of literature references reporting as host | Is the larva feeding on this host? (only confirmed cases) | Does it complete the life cycle? (only confirmed cases) |
|---|---|---|---|---|---|
| Solanaceae | |||||
| Capsicum annuum (bell and chilli peppers) | 1 | Fruit – no | 3 | No | No |
| Physalis angulata (= P. minima; angular winter cherry) | 0 | Fruit – no | 2 | No information | No information |
| Physalis peruviana (cape gooseberry) | 0 | Fruit – no | 3 | No information | No information |
| Solanum aculeatissimum (love‐apple) | 2 | Fruit – yes | 2 | No information | No information |
| S. aethiopicum (= S. integrifolium; gilo/garden egg(plant)) | 2 | Fruit – yes | 9 | Yes | No information |
| S. aethiopicum × S. melongena | 0 | Fruit – yes | 3 | No information | No information |
| S. anguivi | 2 | Fruit – yes | No information | No information | |
| S. anomalum | 0 | Fruit – yes | 2 | Yes | No information |
| S. donianum (mullein nightshade) | 0 | Fruit – yes | 1 | No information | No information |
| S. incanum | 0 | Fruit – yes | 3 | Yes | No information |
| S. incanum × S. melongena | 0 | Fruit – yes | 2 | Yes | No information |
| S. linnaeanum (= S. undatum; devil's apple) | 1 | Fruit – yes | No information | No information | |
| S. lycopersicum (tomato) | 0 | Fruit – yes | 16 | Yes | Yes |
| S. macrocarpon (African eggplant) | 2 | Fruit – yes | 5 | Yes | Yes |
| S. macrocarpon × S. melongena | 0 | Fruit – yes | 1 | No information | No information |
| S. melongena (eggplant/brinjal/aubergine) | 222 | Fruit – yes | 467 | Yes | Yes |
| S. melongena × S. viarum | 0 | Fruit – yes | 1 | No information | No information |
| S. melongena × S. violaceum (‘indicum’) | 0 | Fruit – yes | 1 | No information | No information |
| S. myriacanthum (Himalayan nightshade) | 0 | Fruit – yes | 2 | Yes | No information |
| S. nigrum (black nightshade) | 0 | Fruit – yes | 14 | Yes | Yes |
| S. stramoniifolium | 1 | Fruit – yes | No information | No information | |
| S. torvum (turkey berry) | 66 | Fruit – yes | 8 | Yes | No information |
| S. tuberosum (potato) | 0 | Tubers – no | 28 | Yes | Yes |
| S. viarum (tropical soda apple) | 0 | Fruit – yes | 1 | Yes | No information |
| S. violaceum | 0 | Fruit – yes | 7 | No information | No information |
| S. virginianum (= S. xanthocarpum schrad.; yellow‐fruit nightshade) | 8 | Fruit – yes | 5 | Yes | Yes |
| Solanum sp. | 26 | Fruit – yes | 2 | Yes | No information |
| Anacardiaceae | |||||
| Mangifera indica/sp. (mango) | 3 | Fruit – no | 3 | No information | No information |
| Spondias dulcis (golden apple) | 1 | Fruit – no | 0 | No information | No information |
| Asteraceae | |||||
| Helianthus sp. (sunflower) | 0 | No information | 1 | No information | No information |
| Convolvulaceae | |||||
| Ipomoea batatas (sweet potato)* | 1 (ipomoea aquatica) | Tubers – no | 1 | Yes | No |
| Cucurbitaceae | |||||
| Momordica sp. | 4 | Fruit – no | 0 | No (m. Charantia) | No |
| Fabaceae | |||||
| Cajanus cajan (pigeon pea) | 0 | Pods – no information | 1 | No information | No information |
| Cicer arietinum (chickpea) | 0 | Pods – no information | 1 | No information | No information |
| Pisum sativum (pea) | 0 | Pods – no information | 2 | No information | No information |
| Vigna unguiculata (= V. sinensis; cowpea) | 1 (Vigna sp.) | Pods – yes | 1 | Yes | No |
| Lamiaceae | |||||
| Ocimum sp. (basil) | 2 | No information | 0 | No information | No information |
| Malvaceae | |||||
| Abelmoschus esculentus (okra) | 0 | Pods – yes | 1 |
Yes (Ardez et al., 2008); No (Maureal et al., 1982) |
No |
| Gossypium sp. (cotton) | 0 | No information | 1 | No information | No information |
| Myrtaceae | |||||
| Psidium guajava (guava) | 2 | Fruit – no | 0 | No information | No information |
| Poaceae | |||||
| Zea mays (corn) | 0 | No information | 1 | No information | No information |
| Sorghum sp. (broomcorn) | 0 | No information | 1 | No information | No information |
| Rosaceae | |||||
| Rosa sp. (rose) | 0 | No information | 1 | No information | No information |
| Rutaceae | |||||
| Citrus hystrix (kafir lime) | 1 | Fruit – no | 0 | No information | No information |
| Murraya paniculata (orange jasmine) | 1 | Fruit – no | 0 | No information | No information |
| Total | 350 | 604 | |||
As Table C.1 shows, the majority of non‐Solanaceae plant species listed as host are based either on a few interceptions, many of which were in mixed consignments with eggplant, or on a few reports in the literature that usually refer to older publications or don't even provide a reference at all. These plant associations were deemed incidental, and thus dismissed as principal pathways for introduction. However, two cases stand out in Table C.1: Momordica sp. with four interceptions, and mango (Mangifera indica) with three interceptions and three reports in the literature.
The claim of mango as host plant can be traced back to Hutson (1931), which is solely based on an abstract of a 17‐page typescript that apparently never has been published, and that states ‘the shoot‐borer, Leucinodes orbonalis, Gn., and the Pentatomid, Coptosoma siamica, Wlk., on mango’. The association with mango therefore remains dubious, especially since the pentatomid true bug Coptosoma siamica is known to only feed on Desmodium novaehollandiae latifolium (Fabaceae) (Rider, 2015), and not on mango as stated in the Hutson (1931) abstract. In the case of Momordica sp., there is no information in the literature reporting bitter melon as a host. In 24‐h no‐choice tests, Maureal et al. (1982) found that third‐instar larvae did not feed on Momordica charantia (bitter melon) fruit. Both mango and bitter melon are therefore also excluded as principal pathways for introduction.
Among Solanaceae, Capsicum was demonstrated to not be eaten by the larvae (Maureal et al., 1982); Physalis was only cited in the literature, but never intercepted in association with L. orbonalis, nor shown to be fed on by the larvae.
Isahaque and Chadhuri (1983) report S. torvum (turkey berry) to be an alternative host. Turkey berry is the second‐most common interception commodity of L. orbonalis in the EU, after eggplant. In Solanum species with small fruits, like S. nigrum and potentially also S. torvum, the individual fruits do not provide enough space for the developing larva to feed inside; instead, when the fruit becomes too small to sustain the caterpillar, it exits the fruit, spins together several fruits with a web and feeds from within this web externally on the fruits (Das & Patnaik, 1971). Leucinodes orbonalis infestations on imported fruits of such Solanum species should therefore be easily recognised, at least when infested with older larvae too large to feed inside the fruits. Turkey berry fruits have a diameter of 10–13 mm (Vorontsova & Knapp, 2023a).
Ardez et al. (2008) observed in their lab experiments that moths reared from S. nigrum (black nightshade) fruits did not differ in their longevity from those reared from eggplant, and they conclude that S. nigrum is the most suitable alternative host plant for L. orbonalis. At the same time, Das & Patnaik (1971) report that adults reared from S. nigrum were smaller in size, and females laid far fewer eggs than compared to eggplant‐ or tomato‐reared specimens; nonetheless, all eggs were viable, and the insect can complete its life cycle on this plant. Das & Patnaik (1971) report that the fruits are too small for the developing larvae, and they therefore exit the fruit they were feeding in as small larva, and web three to four fruits together, feeding on them from inside the web. Navasero and Calilung (1990) confirmed this larval behaviour of feeding on spun‐together fruit clusters. The diameter of black nightshade fruits is 5–13 mm (Coleman et al., 2020). Although no study investigated whether L. orbonalis larvae can also feed on other parts of the plant, Panel assumes that they also bore into the shoots of black nightshade.
In India, L. orbonalis larvae were reported to occasionally bore into the shoots and leaf stems of potato (S. tuberosum) plants, and infestation can reach up to 40% (Nair, 1967). In the lab, L. orbonalis larvae can be reared with potato tubers (Ardez et al., 2008; Boopal et al., 2013; Kariyanna et al., 2020a, 2020b; Maureal et al., 1982). The longevity of L. orbonalis adults reared on potato tubers was significantly shorter than of those reared on fruits of S. melongena and S. nigrum (Ardez et al., 2008). Maureal et al. (1982) observed a longer developmental period, higher mortalities especially in later larval instars, and a shorter adult lifespan in specimens reared on potato tubers as compared to specimens reared on eggplant. There are no published reports of L. orbonalis larvae feeding on tubers in the wild, and the species appears to feed solely on plant parts above ground.
Das & Patnaik (1971) successfully reared L. orbonalis on the fruits of S. lycopersicum (tomato), with a somewhat lower fecundity of the resulting females as compared to S. melongena; they furthermore state that in Bhubaneswar (India), the larvae were observed to also bore into the shoots of tomato plants. Jethva and Vyas (2009) conducted laboratory rearings on tomato, but they did not specify the plant tissue they provided the larvae for feeding. Maureal et al. (1982) observed a longer developmental period, higher mortalities especially in later larval instars, and a shorter adult lifespan in specimens reared on tomato fruits as compared to specimens reared on eggplant.
Tejavathu et al. (1991) find S. anomalum to be ‘resistant’ to L. orbonalis, despite 5.9% shoot‐ and 11.3% fruit infestation. No information is available on whether the species can complete its life cycle on this plant. Solanum anomalum has comparably small fruits, with diameters of 5.5–9 mm (Vorontsova & Knapp, 2023b). Solanum anomalum is endemic to Africa, and the species reported by Tejavathu et al. (1991) might be a misidentification of S. aethiopicum or S. anguivi, the wild progenitor of S. aethiopicum (Vorontsova & Knapp, 2023b). Hanur et al. (2011) report that L. orbonalis can successfully develop all larval stages on S. macrocarpon before pupating in the soil, and that ‘Patterns and extent of infestation and damage on the affected parts were comparable in both S. melongena and S. macrocarpon […], indicating that S. macrocarpon is equally susceptible to infestation by BSFB’. Srinivasappa et al. (1998) report an infestation rate of 10.16% of fruits, whereas according to Kumar and Sadashiva (1991), fruit damage by L. orbonalis larvae is less than 1%. Isahaque and Chadhuri (1983) report feeding on S. myriacanthum (Himalayan nightshade), who observed the larvae boring into the shoots. It is unknown whether the small fruits (diameter 20–30 mm; Nee, 2023) are also consumed. Udayagiri and Mohan (1985) report L. orbonalis from S. viarum (tropical soda apple), stating ‘The caterpillars were noticed feeding on the contents of the developing berries. The entry holes were plugged with excreta. L. orbonalis infestation was observed to be low’. Since the fruits are fairly small (22–25 mm in diameter according to Nee et al., 2023), the later‐stage larvae may also feed from the outside, as reported for S. nigrum by Das & Patnaik (1971). Similarly small fruits (15–25 mm; Knapp, 2023) are present in S. virginianum (= S. xanthocarpum Schrad.; yellow‐fruit nightshade), which Lall and Ahmad (1965) report (as S. xanthocarpum) to be a wild host of L. orbonalis ‘during off season (April to August)’, i.e. when S. melongena is not grown.
Baksh (1979), Behera and Gyanendra (2002), Shinde (2007) and Pugalendhi et al. (2010) report shoot‐ and/or fruit infestation of hybrids of eggplant (S. melongena) with S. aethiopicum, S. incanum, S. viarum and S. violaceum (as syn. S. indicum), but it remains unclear whether the insect's life cycle can be successfully completed on these hybrids.
In conclusion, the principal pathways for the potential entry of L. orbonalis into the EU are judged to be with fruit of Solanum melogena (eggplant) and of Solanum torvum (turkey berry).
Introduction model: Quantity of imports
As noted above, many potential pathways for entry were considered although the commodities judged most feasible were the import of exotic and specialist varieties of eggplant fruit (Solanum melongena) and turkey berry fruit (S. torvum).
Solanum melongena is a widely consumed solanaceous crop with many common names including aubergine, brinjal, eggplant, garden egg, melanzana and patlican. The related Solanum aethiopicum has a similar common name of Ethiopian eggplant, while Solanum torvum is known as turkey berry or pea eggplant. There is a wide range of cultivars of S. melongena with differences in growth habits, fruit size, shape and colour. Depending on cultivar, the length of the fruits varies from approximately 5 cm to more than 30 cm. Fruit may be elongated, cylindrical, ovoid, slender or spherical. Fruit can be purple, green, white, yellow and variegated. The most familiar varieties grown and consumed in Europe are deep purple with a very high gloss.
The market for eggplant can be divided into three sectors: common, special and exotic varieties (Table C.2). A relatively small amount of eggplants are imported into the EU from countries where L. orbonalis occurs. Asian eggplants are niche varieties imported by specialist buyers (CBI, 2020).
TABLE C.2.
Market segmentation of fresh eggplant in the EU.
| Eggplant category | Target outlets | Examples |
|---|---|---|
| Common varieties | Supermarkets, General greengrocers | Typical and common EU grown eggplant varieties |
| Special varieties | Supermarkets, Specialist shops, Street markets | Graffiti eggplant, Japanese eggplant |
| Exotic varieties | Ethnic shops, Restaurants | White eggplant, Thai eggplant, Chinese eggplant |
Exotic and special varieties of eggplants are typically imported in standard 5 kg boxes (Re:fresh Directory, 2018).
Overview of model input estimates: Import quantity
Based on EU imports of eggplants over the period of 2010–2019 from countries where L. orbonalis occurs (Table C.3), the mean annual import was calculated. These countries are Austria (AT), Belgium (BE), Bulgaria (BG), Cyprus (CY), Czech Republic (CZ), Germany (DE), Denmark (DK), Estonia (EE), Spain (ES), Finland (FI), France (FR), Greece (GR), Croatia (HR), Hungary (HU), Ireland (IE), Italy (IT), Latvia (LT), Luxembourg (LU), Latvia (LV), Malta (MT), the Netherlands (NL), Poland (PL), Portugal (PT), Romania (RO), Slovenia (SI), Slovakia (SK) and Sweden (SE).
From 2010 to 2014 imports declined then recovered (Figure C.1); between 2016 and 2019 they were relatively stable, ranging from a minimum of 157 tonnes in 2018 to a maximum of 182 tonnes in 2016. An import ban was established on Indian eggplant from 2014 to 2016 but was not extended due to improved phytosanitary measurements (https://www.freshfruitportal.com/news/2017/01/04/eu‐lifts‐restrictions‐on‐indian‐eggplant‐and‐other‐vegetables/). Trade during 2020 and shortly afterwards was not considered as the Covid‐19 pandemic had disrupted trade flows (Table C.3).
TABLE C.3.
Direct annual import of aubergines (CN 070930) from countries with reported presence of Leucinodes orbonalis (Bangladesh, Brunei Darussalam, China, Indonesia, India, Japan, Cambodia, Lao People's Democratic Republic, Sri Lanka, Myanmar [Burma], Malaysia, Nepal, Philippines, Pakistan, Singapore, Thailand, Taiwan, Viet Nam) to EU member states from 2010 to 2022 (in t, calculated from EUROSTAT table DS‐045409, downloaded on 13th June 2023, status 16th May 2023). The years 2020–2022 (grey shading) were excluded from the calculation of the mean annual import volume, as these were affected by the Covid‐19 pandemic.
| MS | 2010 | 2011 | 2012 | 2013 | 2014 | 2015 | 2016 | 2017 | 2018 | 2019 | 2020 | 2021 | 2022 | Mean | CV (%) | Prop. (%) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Tot.(EU) | 243.426 | 179.251 | 175.625 | 140.893 | 98.81 | 126.386 | 178.601 | 163.744 | 155.773 | 173.81 | 123.685 | 130.912 | 121.698 | 163.6 | 23% | 100% |
| AT | 1.506 | 3.032 | 1.943 | 1.644 | 3.272 | 4.267 | 2.605 | 2.405 | 1.212 | 1.340 | 0.757 | 0.851 | 0.557 | 2.3 | 42% | 1.4% |
| BE | 28.300 | 30.900 | 44.600 | 35.500 | 6.789 | 0.018 | 0.927 | 1.383 | 0.049 | 0.447 | 0.040 | 0.374 | 0.000 | 14.9 | 119% | 9.1% |
| BG | 0.0 | 0.0% | ||||||||||||||
| CY | 0.0 | 0.0% | ||||||||||||||
| CZ | 0.050 | 0.092 | 0.049 | 1.006 | 0.141 | 3.030 | 48.631 | 41.791 | 42.358 | 49.365 | 29.880 | 47.146 | 46.610 | 18.7 | 125% | 11.4% |
| DE | 129.686 | 73.579 | 76.056 | 48.083 | 30.439 | 24.318 | 27.137 | 28.422 | 27.050 | 27.101 | 24.987 | 13.251 | 6.573 | 49.2 | 70% | 30.1% |
| DK | 29.016 | 11.843 | 5.317 | 9.204 | 12.218 | 14.226 | 12.979 | 11.451 | 10.799 | 11.351 | 11.243 | 10.751 | 6.036 | 12.8 | 48% | 7.8% |
| EE | 0.0 | 0.0% | ||||||||||||||
| ES | 0.000 | 0.000 | 0.040 | 0.000 | 0.000 | 0.002 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.0 | 300% | 0.0% |
| FI | 1.180 | 1.044 | 0.000 | 0.339 | 0.315 | 0.000 | 0.319 | 1.187 | 0.351 | 0.000 | 0.000 | 0.000 | 0.000 | 0.5 | 102% | 0.3% |
| FR | 13.166 | 17.178 | 10.070 | 12.065 | 4.703 | 37.032 | 51.835 | 47.357 | 44.785 | 57.912 | 43.585 | 43.298 | 37.161 | 29.6 | 68% | 18.1% |
| GR | 0.0 | 0.0% | ||||||||||||||
| HR | 0.0 | 0.0% | ||||||||||||||
| HU | 0.0 | 0.0% | ||||||||||||||
| IE | 0.216 | 0.000 | 0.435 | 1.049 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.304 | 0.045 | 1.146 | 1.266 | 0.2 | 169% | 0.1% |
| IT | 0.0 | 0.0% | ||||||||||||||
| LT | 0.0 | 0.0% | ||||||||||||||
| LU | 0.0 | 0.0% | ||||||||||||||
| LV | 0.0 | 0.0% | ||||||||||||||
| MT | 0.0 | 0.0% | ||||||||||||||
| NL | 7.687 | 40.470 | 35.920 | 30.713 | 39.832 | 37.296 | 29.973 | 26.787 | 22.309 | 15.722 | 8.090 | 9.314 | 19.611 | 28.7 | 38% | 17.5% |
| PL | 0.150 | 0.000 | 0.000 | 0.000 | 0.017 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.0 | 282% | 0.0% |
| PT | 0.015 | 0.000 | 0.000 | 0.155 | 0.000 | 0.000 | 0.015 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.0 | 261% | 0.0% |
| RO | 0.0 | 0.0% | ||||||||||||||
| SE | 32.454 | 1.113 | 1.195 | 1.135 | 1.084 | 6.197 | 4.180 | 2.961 | 6.860 | 10.268 | 5.058 | 4.781 | 3.884 | 6.7 | 142% | 4.1% |
| SI | 0.0 | 0.0% | ||||||||||||||
| SK | 0.0 | 0.0% |
FIGURE C.1.
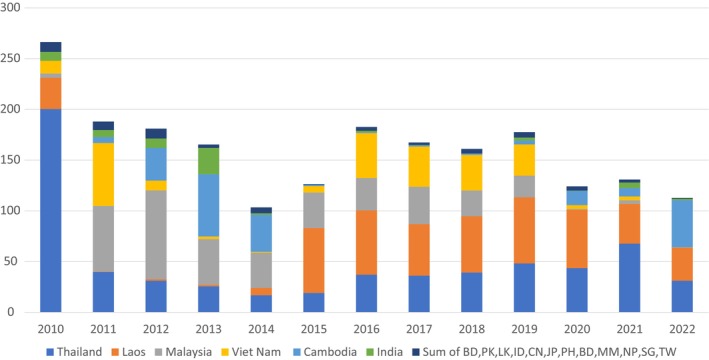
Tonnes of eggplants imported into EU 27 in 2010–2022 from countries where Leucinodes orbonalis is known to occur. (EUROSTAT, EU trade (ds‐059322) by CN, status 15th November 2023).
Based on a normal distribution and the annual imports from 2010–2019, a mean import volume of 163.6 tonnes per year was calculated, with a standard derivation of 38.37 t/year and a coefficient of variation of 23%.
Uncertainties
The Panel assumed that trade over the next 5 years would be similar to trade in the recent past (2010–2019) (Table C.3 and Figure C.2). In reality, trade may increase or decrease quickly in response to markets and consumer preferences. This uncertainty was not accounted for but was considered small compared to uncertainty in other parameters of the pathway model.
FIGURE C.2.
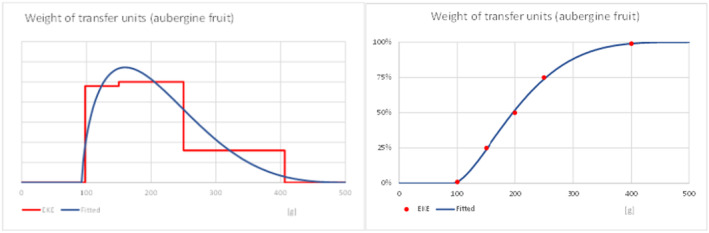
Distribution of weight of exotic/special eggplant varieties fitted to EKE estimates. (Left hand chart shows probability density function density function to describe the remaining uncertainties of the parameter; right hand chart shows cumulative distribution function (CDF) of the likelihood of the parameter).
Introduction model: Weight of an eggplant fruit
There is variation in the size and weight of exotic and special eggplant cultivars imported into the EU. Examples are provided in Table C.4. Eurostat data do not distinguish between eggplant cultivars and therefore an EKE was performed to estimate the mean weight of an exotic eggplant fruit. Eggplants from Bangladesh, India, and Thailand are typically exported in boxes of 5 kg; exports from Pakistan are more variable and do not have a standardised box weight (FPJ, 2011).
TABLE C.4.
Example dimensions and weights of exotic eggplant varieties (Source: https://plantura.garden/uk/vegetables/aubergine/aubergine‐varieties).
| Type | Shape | Colour | Radius (cm) | Length (cm) | Approximate volume (cm3) | Weight (g) | Fruits per 5 kg box |
|---|---|---|---|---|---|---|---|
| Thai eggplant | Spherical | Green variegated | 5.7 | – | 249.5 | 78 | 65 |
| Japanese eggplant | Elongated | Very dark purple | 1.9 | 20.3 | 231.7 | 142 | 35 |
| White eggplant | Elongated | White | 2.4 | 18.4 | 327.8 | 163 | 30 |
| Chinese eggplant | Elongated | Light purple | 2.5 | 39.4 | 797.9 | 312 | 16 |
| Philippine eggplant | Elongated | Purple and green | 3.7 | 24.1 | 1009.9 | 397 | 12 |
TABLE C.5. Estimated mean weight of a fresh exotic/special eggplant fruit (grams).
| Question | What is the mean weight of a fresh exotic/special eggplant variety (in grams) imported into the EU from countries where Leucinodes orbonalis occurs? | ||||||
|---|---|---|---|---|---|---|---|
| Results | Estimated mean weight of exotic/special fresh eggplant fruit imported into EU (kg) | ||||||
| Percentiles % | 1% | 5% | 25% | 50% | 75% | 95% | 99% |
| EKE estimates | 100 | – | 150 | 200 | 250 | – | 400 |
| Fitted results (g) | 100 | 113 | 152 | 197 | 253 | 343 | 401 |
| Fitted distribution | BetaGeneral (1.6549, 4.6322, 93,530) | ||||||
TABLE C.6. Estimated mean weight of a fresh turkey berry fruit cluster (grams).
| Question | What is the mean weight of a fresh turkey berry fruit cluster (in grams) imported into the EU from countries where Leucinodes orbonalis occurs? | ||||||
|---|---|---|---|---|---|---|---|
| Results | Estimated mean weight of fresh turkey berry cluster imported into EU (kg) | ||||||
| Percentiles % | 1% | 5% | 25% | 50% | 75% | 95% | 99% |
| EKE estimates | 18 | – | 27 | 35 | 42 | – | 55 |
| Fitted results (g) | 18.0 | 20.3 | 27.3 | 34.5 | 42.4 | 51.4 | 55.0 |
| Fitted distribution | BetaGeneral (1.5622, 1.9533, 16.8, 57.8) | ||||||
TABLE C.7. Uncertainties associated with low and high fruit weight scenarios.
| Uncertainty | Low weight scenario | High weight scenario |
|---|---|---|
| Fruit size | Mainly small eggplant and turkey berries | Mainly large eggplant fruit and turkey berries |
| Cluster size | Small number of turkey berries per cluster (infructescence) | Large number of turkey berries per cluster (infructescence) |
TABLE C.8.
Imports of eggplant from non‐EU countries to the Netherlands in 2022, by country (kg).
| Country | Minimum | P25 | P50 | Mean | P75 | Maximum | Number | Sum |
|---|---|---|---|---|---|---|---|---|
| Egypt | 140 | 150 | 160 | 200.0 | 230 | 300 | 3 | 600 |
| Ghana | 20 | 32.5 | 50 | 127.7 | 177.5 | 404 | 7 | 894 |
| Jordan | 104 | 557 | 803 | 1081.0 | 1549 | 2587 | 38 | 41,079 |
| Kenia | 12 | 14 | 16 | 16.0 | 18 | 20 | 2 | 32 |
| Laos | 1 | 19.75 | 28.75 | 32.1 | 32.1 | 33.9 | 4 | 92 |
| Malaysia | 30 | 33.45 | 36.9 | 36.9 | 40.35 | 43.8 | 2 | 74 |
| Morocco | 20 | 1006 | 2156 | 2626.0 | 3520 | 14,520 | 58 | 152,304 |
| Mexico | 15 | 390 | 629.5 | 725.0 | 910 | 3510 | 84 | 60,900 |
| Uganda | 7 | 54 | 699 | 682.4 | 975 | 1890 | 8 | 5459 |
| Spain | 126 | 136.5 | 154 | 207.0 | 294 | 308 | 6 | 1449 |
| Surinam | 10 | 90 | 150 | 190.2 | 252.2 | 1708 | 420 | 79,873 |
| Thailand | 0.291 | 21.2 | 24.7 | 23.4 | 27.9 | 32.55 | 31 | 724 |
| Turkey | 60 | 450 | 900 | 1364.0 | 1350 | 11,170 | 42 | 57,297 |
| South Africa | 1.2 | 19.2 | 42 | 54.2 | 72 | 1250 | 499 | 27,045 |
| Total | 0.291 | 37.2 | 96 | 355.0 | 275 | 14,520 | 1205 | 427,822 |
TABLE C.9. Estimated mean number of eggplant fruits infested with L. orbonalis when entering the EU (per 10,000 fruit).
| Question: | How many out of 10,000 transfer units (fresh eggplant fruit) will be on average infested with L.orbonalis when entering the EU from countries where the pest occurs? | ||||||
|---|---|---|---|---|---|---|---|
| Results | Infestation rate of eggplant fruit when entering the EU (per 10,000 fruit) | ||||||
| Percentiles: % | 1% | 5% | 25% | 50% | 75% | 95% | 99% |
| EKE estimates: | 0.1 | – | 1.5 | 3 | 12 | – | 30 |
| Fitted values: (fruit infested per 10,000) | 0.100 | 0.127 | 0.983 | 4.20 | 11.1 | 23.7 | 30.1 |
| Fitted distribution | BetaGeneral (0.46872, 1.8525, 0.099, 35) | ||||||
TABLE C.10. Scenario calculations on the likely level of infestation with L. orbonalis in eggplant fruit.
| Step | Detail | Data (source) | Scenario | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | |||
| x1 | Import into EU of aubergines (CN code 070930) from countries with L. orbonalis 2010–2022 (kg) | From Eurostat | 160,000 | 160,000 | 160,000 | 160,000 | 160,000 | 160,000 | 160,000 | 160,000 | 160,000 |
| x2 | Weight of a single consignment (kg) | Three scenarios: 12, 24, 48 | 12 | 24 | 48 | 12 | 24 | 48 | 12 | 24 | 48 |
| x3 | Number of consignments | Calculated: x1/x2 | 13,333 | 6667 | 3333 | 13,333 | 6667 | 3333 | 13,333 | 6667 | 3333 |
| x4 | Percentage of inspected consignments | Constant: 100% | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 |
| x5 | Number of consignments inspected | Calculated: x3 * x4/100 | 13,333 | 6667 | 3333 | 13,333 | 6667 | 3333 | 13,333 | 6667 | 3333 |
| x6 | Number of consignments found contaminated with L. orbonalis per year 2010–2022 | Reported to Europhyt | 11.5 | 11.5 | 11.5 | 11.5 | 11.5 | 11.5 | 11.5 | 11.5 | 11.5 |
| x7 | Number of fruit/bunches inspected per consignment | Dutch NVWA info and scenarios | 60 | 60 | 60 | 30 | 30 | 30 | 10 | 10 | 10 |
| x8 | Estimated proportion of infested fruit/bunches | Calculated: ‐ln(1‐x6/x5)/x7 | 1.44E‐05 | 2.88E‐05 | 5.76E‐05 | 2.88E‐05 | 5.75E‐05 | 0.000115 | 8.63E‐05 | 0.000173 | 0.000346 |
| x9 | Infested fruit/bunches per 10,000 | Calculated: x8 * 10,000 | 0.144 | 0.288 | 0.576 | 0.288 | 0.575 | 1.152 | 0.863 | 1.726 | 3.456 |
TABLE C.11. Uncertainties associated with low and high infestation rate scenarios.
| Uncertainty | Low infestation rate scenario | High infestation rate scenario |
|---|---|---|
| Infestation in field | Low pest pressure | High pest pressure |
| Sorting | High quality for export | Inadequate sorting |
| Control at border | Good border control: large sample size, visual control | Poor border control: small sample size, only identity check |
| Efficiency of visual control | Clearly visible entry holes | Difficult detection of eggs, old entry holes |
The median weight of an exotic/special eggplant variety is 197 g; (90% CR is from 113 to 343 g).
Uncertainties
There are more varieties than those given as examples in Table C.4.
There is more variation in the dimensions and weights of exotic varieties than shown in Table C.4.
Eggplants from Bangladesh, India and Thailand are typically exported in boxes of 5 kg; exports from Pakistan are more variable and do not have a standardised box weight (re:Fresh Directory, 2018).
Reasoning
Weights of exotic eggplant varieties range from 78 to 397 g (Table C.4).
Turkey berries have a diameter of 10–13 mm (Vorontsova & Knapp, 2023a).
5–40+ turkey berries per infructescence (Vorontsova & Knapp, 2023a).
Lower limit:
Predominantly small‐fruited eggplants with light weight imported;
Predominantly small clusters of turkey berry with small berries imported.
Upper limit:
Predominantly large varieties of eggplant imported;
Predominantly large clusters of turkey berry with large berries imported.
Median:
Imported eggplant fruit represents a mix of large and small varieties.
Imported turkey berries represent a mix of small and large berry clusters.
Inter‐quartile range:
Relatively high uncertainties below and above the median for both eggplant and turkey berry.
Introduction model: Infestation rate of eggplant fruit when entering the EU
According to hearing expert Dr Ramasamy, in subtropical and tropical Asia where eggplant is grown, fruit infestation with L. orbonalis usually ranges from 25% to 30% at the lower end and can reach up to 100%; extremely well‐managed fields can have as little as 10% infestation. The current practice in many Asian eggplant‐producing countries is to apply large amounts of (cheap) pesticides during the entire growth season to keep L. orbonalis infestations rates as low as possible, resulting in increasing pesticide resistance (e.g. Shirale et al., 2017). Pheromone mass trapping of males is very common and is part of a (still poorly adapted) integrated pest management (IPM) package that further includes healthy seedling production, the removal and prompt destruction of infested shoots and fruits, and the withholding of chemical pesticides to promote natural enemies of L. orbonalis. These IPM measures can reduce the infestation to 20%–30% and lower the intensity of pesticide application from 90 sprays to 15–20 sprays per growth cycle (Dr S. Ramasamy, personal communication).
No methods are currently known to physically, chemically or spectrometrically detect L. orbonalis infestations of eggplant fruit in a non‐invasive way.
Dossier findings are summarised below.
Eggplant fruit can be harvested 3–4 months after propagation by seed.
Under favourable conditions, flowering and fruit production is continuous, consequently eggplants are repeatedly harvested by hand.
Fruits grow rapidly under optimum conditions and plants are harvested frequently.
Cotton gloves are often worn during harvest to protect fruit against physical injury.
Fruits are handled carefully to avoid bruising and compression injury.
Fruits are graded then individually wrapped with paper or plastic film.
Careful handling of each individual fruit provides opportunity to detect infested fruit and to reject these from export.
Fruits are packed in bags, crates or baskets.
Fruits undergo rapid (pre‐)cooling to 10°C immediately after harvest to inhibit discolouration, weight loss, drying of calyx, and decay.
Eggplant fruit are sensitive to chilling (temperatures below 10°C). Damage such as surface pitting, surface bronzing, and seed and tissue browning will result if stored for a few days below 10°C.
Storage: Fruit are stored at 10–12°C with 90–95% RH. (WG interpretation: insufficient to cause larval mortality).
Eggplant can be stored for 2–3 weeks but are generally held for less than 14 days as visual and sensor qualities deteriorate rapidly. Decay is likely to increase after storage > 2 weeks, especially after removal to typical retail conditions.
Eggplants must have a phytosanitary certificate before being brought into the EU. Eggplants intended for the EU therefore have to be inspected and found free from quarantine pests, specifically Spodoptera frugiperda and Thrips palmi. Eggplant fruit should also be practically free from other pests (EU 2019/2072, Annex VII).
However, there have been interceptions of L.orbonalis from a number of Asian countries.
Pest issues
The Panel analysed Dutch NVWA data for 2022 on the size of incoming shipments of Solanum fruit with CN code 070930, comprising a.o. S. melongena and S. torvum, here referred to as eggplant sensu lato (s.l.). Fourteen extra‐EU countries imported eggplant s.l. to EU. Shipment sizes differed greatly between countries of origin, indicating that some countries send product in bulk, whereas other send small shipment of, probably, a specialty product, e.g. Solanum torvum berries. Shipments from Asian countries were on average small compared to shipments from most other countries of origin (Table C.8, Figure C.3).
FIGURE C.3.
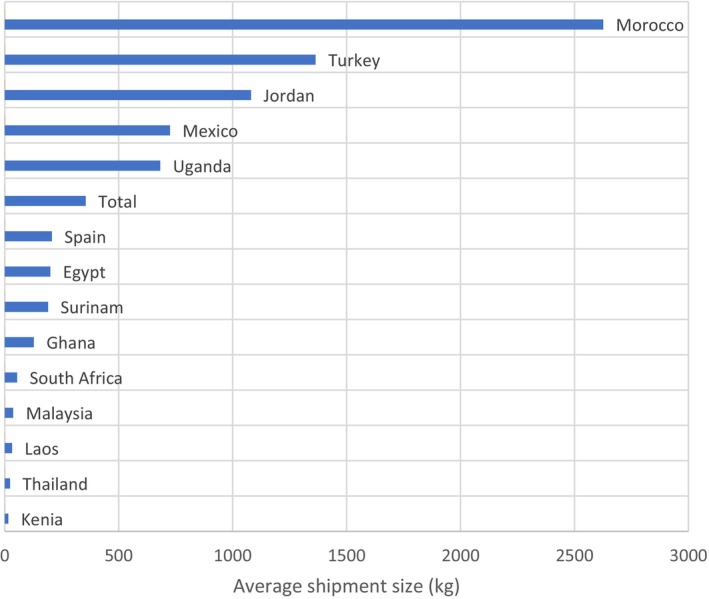
Average estimated shipment size of eggplants shipments of countries (based on imports by NL in 2022).
The figure below (based on the same data) ranks the countries by the size of average shipments, showing that shipment sizes of aubergine s.l. from Asia are among the smallest of countries of origin of eggplant.
Usually, the number of interceptions per 100 kg of product is quite low. The Panel estimated a level of infestation in product coming from Asia from 0.13 to 24 per 10,000 fruits (approximately 2000 kg for eggplant or 450 kg for turkey berry). However, occasionally, from a country of origin, this number can be higher, as is the case for Sri Lanka and Pakistan in Figure C.4. In such cases, the number of infestations will diminish over time as stricter phytosanitary measurements are implemented (dashed lines in Figure C.4), to reach the required level of pest freedom, i.e. the absence of the pest from the exported commodity.
FIGURE C.4.
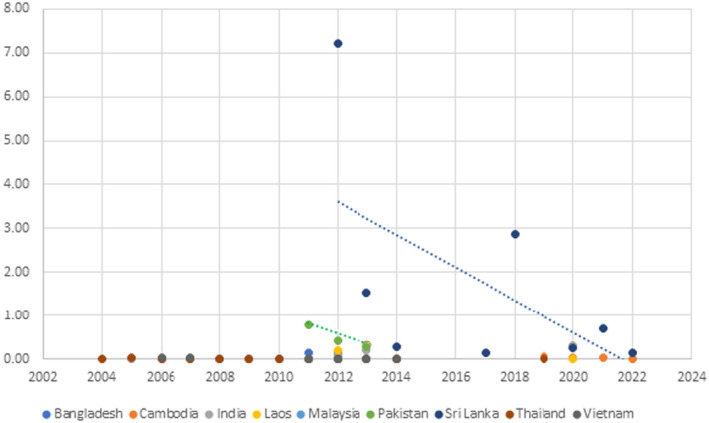
Number of Leucinodes orbonalis interceptions per 100 kg of eggplant (Solanum melongena) imported from Asian countries. Dashed lines indicate changes of interception proportions over time in imports from Pakistan (green) and Sri Lanka (dark blue).
There are no data on the number of inspections conducted to produce the number of interceptions reported in Europhyt and TRACES. This severely weakens how Figures C.1 and C.4 (Interceptions plotted against time) can be interpreted.
Post‐harvest issues
After harvest, eggplant fruits are handled very carefully to avoid physical damage.
Eggplants are carefully cleaned in the field.
Fruit are gently rubbed to remove debris and soil particles.
Harvested fruit should be protected from environmental conditions and kept in shaded areas.
Plastic crates are recommended when transporting the eggplants from the field to minimise damage.
In cleaning, sorting, grading pack houses fruit can be washed and dried.
Eggplants are stored at 10°C or more (chill sensitive when stored below 10°C).
Eggplant is stored separately from tomatoes, bananas and melons – their high ethylene content causes eggplants to spoil quickly.
Sources: Cargo Handbook.com; Majlis Ilmu, 2018.
Overview of model input estimates: infestation rate on arrival into EU
Elicited mean annual rates of infestation per 10,000 eggplant fruits are given in the table below with the probability distribution under the table. EKE estimates are values proposed by the expert working group as consensus estimates. Model inputs are derived from the distribution fitted to the EKE estimates.
Figure C.5
FIGURE C.5.
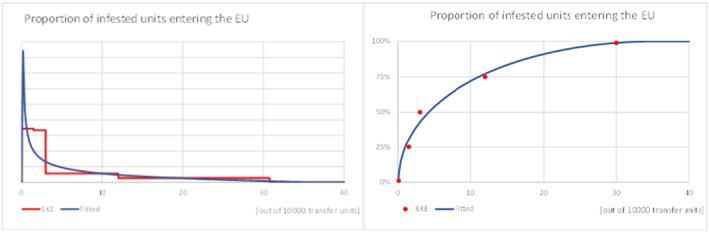
Distribution of infestation rate of eggplant fruit fitted to EKE estimates (left hand chart shows probability density function density function to describe the remaining uncertainties of the parameter; right hand chart shows cumulative distribution function (CDF) of the likelihood of the parameter).
The median rate of eggplant fruit infestation is 4.20 per 10,000 (= 42 per 100,000); (90% CR is from 12.7 per million to 2370 per million).
Reasoning
The Panel used Europhyt and TRACES interception data to infer a likely level of infestation of the imported eggplant fruit.
The weights of consignments inspected are recorded as net weight (Europhyt) or commodity weight (TRACES) (kg).
Leucinodes orbonalis was most frequently intercepted on consignments of between 20.1 and 30 kg (Figure C.6).
Assuming that the consignment sizes on which interceptions occur are representative of all consignments, the number of consignments each year can be estimated.
FIGURE C.6.
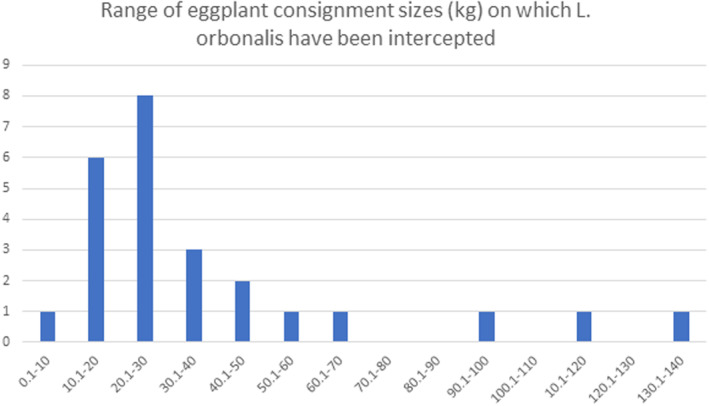
Consignment sizes vary from less than 10 kg to over 130 kg. (NL data).
Analysis
The Dutch NVWA import inspection data for 2022 include 37 records of imports of eggplant s.l. from Asian countries with presence of L. orbonalis, 31 shipments from Thailand, 2 from Malaysia and 4 from Laos.
The mean weight of these 37 shipments was 24 kg, and the mean number of boxes (‘colli’) per shipment was also 24. Thus, the average contents of a box would be approximately 1 kg of fruit. This suggests that these must be small fruit.
According to the inspection guidelines, inspectors should inspect 60 fruit per shipment as a minimum. With a shipment of 24 kg, if one fruit is 200 g, there would be 120 fruits in a shipment. If the commodity is S. torvum, assuming 35 g per bunch, there would be 24/0.035 = 685 bunches in a shipment.
The average import of eggplant (CN code 709300) to EU is 160 t/year. With shipment sizes in the order of 24 kg, in means that about 7000 consignments with aubergine are imported each year to EU from countries with L. orbonalis. Each consignment should be inspected, and the prescribed sample size for inspection is 60 fruits, but Panel might take into account that in the actual practice, this number is not reached. On average, out of these approximately 7000 consignments per year, about 11.5 per year are found infested. The corresponding proportion of infested fruit can be worked out from the formula
where P is the proportion of consignments found infested (median estimate 11.5/6667 = 0.17%. It could be a larger or smaller proportion depending upon whether consignments tend to be smaller or larger than the mean estimate of 24 kg. N is the sample size per consignment which is prescribed to be 60 fruit or bunches but might be smaller in practice (not larger).
The solution is:
Different scenarios are worked out in the table below. The conclusion is reached that the number of infested fruit/fruit clusters per 10,000 is in the order of 1 (range: 0.14–3.5). That is a similar order of magnitude (perhaps a bit less) compared to what was found for Elasmopalpus lignosellus, in asparagus from Peru (EFSA PLH Panel, 2023) but given the much smaller trade of eggplant compared to asparagus, the number of infested fruit/units imported to the EU will be much smaller for eggplant than for asparagus.
Lower limit:
Estimation from the interception data with optimistic assumptions.
Upper limit:
Estimation from interception data with low sample size;
High infestation level at country of origin, with no indications of proper measures.
Median:
General low infestation level, as shown in the interception data;
Limitation of the detection at border control.
Inter‐quartile range:
High uncertainties below and above the median.
APPENDIX D. Establishment
D.1.
This appendix summarises background information that was used to estimate those parameters relevant to assess the likelihood of pest establishment following entry. The establishment component of the overall model for introduction aimed to calculate the number of adult L. orbonalis emerging from imported infested fruit that go on to mate and transfer to hosts and start a founder population.
A detailed description of how environmental parameters were used to inform the assessment of establishment is provided by Rossi et al. (2023). Following a thorough literature review, 573 references were used to provide data to assess establishment. Data from 334 papers were extracted regarding pest distribution for mapping. The assessment focussed on mapping the distribution of L. orbonalis in south Asia and projecting potential distribution in Europe and the around the Mediterranean. Output maps for the four methods used to inform assessment of the area of potential establishment are available on Zenodo, i.e. (i) Köppen–Geiger zones in Europe and in the area where the pest occurs, (ii) mapping of accumulated degree days and number of generations for Europe and Asia, (iii) maps produced using CLIMEX and (iv) the Species Distribution Modelling results.
Degree days and number of generations
Degree days were calculated by accumulating daily mean temperature above a base temperature of 15°C (Base T). A linear regression model using data from Dhaliwal and Aggarwal (2021) was used to estimate that 438.6 accumulated degree days above BaseT were necessary for one generation of L. orbonalis to be completed.
Across large parts of Europe, the maximum degree days possible each year is 1500. Degree days in excess of 4000 per year occur widely across southern Asia (Figure D.1). See also Rossi et al. (2023).
FIGURE D.1.
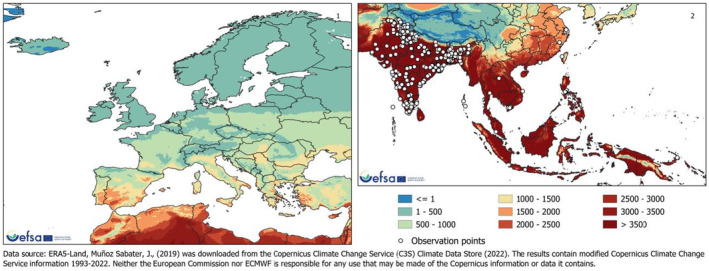
Mean degree days accumulation for Leucinodes orbonalis in Europe and the Mediterranean Basin (1) and South‐East Asia and India (2) based on a base temperature of 15°C. Degree days were calculated for each year in the period 1993–2022 and then averaged.
Figure D.2 indicates large parts of India accumulate sufficient degree days such that there can be 10 or more generations per year. In the EU degree days accumulations would theoretically sustain three to four life cycles of L. orbonalis per year.
FIGURE D.2.
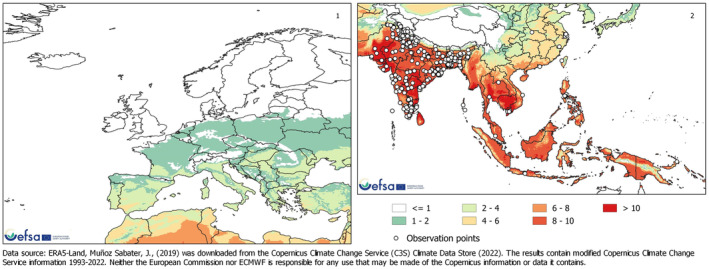
Estimated mean number of generations for Leucinodes orbonalis in Europe and the Mediterranean Basin (1) and South‐East Asia and India (2). Number of generations were calculated considering a minimum number of accumulated degree days to complete one generation of 438.6 degree days. Number of generations were calculated for each year in the period 1993–2022 and then averaged.
CLIMEX projection
Results from the literature review (2.2.1) provided 64 papers that were used to inform the selection of CLIMEX parameters (Rossi et al., 2023). Parameters used in CLIMEX are shown in Table D.1.
TABLE D.1.
CLIMEX parameters for Leucinodes orbonalis. Parameters highlighted in grey were not adjusted and values already included in the CLIMEX ‘semi‐arid’ template were used.
| Class | Parameter | Description | No moisture index | Moisture index | Source |
|---|---|---|---|---|---|
| Moisture index | SM0 | Lower soil moisture threshold | – | 0.01 | Iteration |
| SM1 | Lower optimal soil moisture | – | 0.1 | Iteration | |
| SM2 | Upper optimal soil moisture | – | 3 | Iteration | |
| SM3 | Upper soil moisture threshold | – | 4 | Iteration | |
| Temperature index | DV0 | Lower temperature threshold | 15°C | 15°C | 315 |
| DV1 | Lower optimal temperature | 26°C | 26°C | 24, 315, 1861 | |
| DV2 | Upper optimal temperature | 38°C | 38°C | 24, 315, 1833 | |
| DV3 | Upper temperature threshold | 39°C | 39°C | 1683 | |
| Cold stress | TTCS a | Cold stress temperature threshold | 5.13°C | 5.13°C | a |
| THCS a | Cold stress accumulation rate | −0.00237 week−1 | −0.00237 week−1 | a | |
| DTCS | Cold stress will begin to accumulate when this threshold number of degree days above DVCS is not achieved | 2°C | 2°C | Iteration | |
| DHCS | Rate at which Cold Stress accumulates once the threshold number of degree days above DVCS (DTCS) is not achieved | −0.001 week−1 | −0.001 week−1 | ||
| TTCSA | Average weekly temperature below which Cold Stress accumulates | 0°C | 0°C | ||
| THCSA | Rate at which Cold Stress accumulates once temperatures drop below the threshold value of TTCS1 | 0°C | 0°C | ||
| Heat stress | TTHS | Heat stress temperature threshold | 39°C | 39°C | 1683 |
| THHS | Heat stress accumulation rate | 0.001 week−1 | 0.001 week−1 | ||
| DTHS | Heat stress will begin to accumulate when this threshold number of degree days above DV3 is exceeded | 0°C | 0°C | ||
| DHHS | This is the rate at which Heat Stress accumulates once the threshold number of degree days above DV3 (DTHS) is exceeded | 0°C | 0°C | ||
| Dry stress | SMDS a | Dry stress soil moisture value | 0.09797 | 0.09797 | a |
| HDS a | Dry stress soil moisture rate | −0.00782 week−1 | −0.00782 week−1 | a | |
| Wet stress | SMWS | Soil moisture wet stress threshold | 4 | 4 | Iteration |
| HDS | Wet stress accumulation rate | 0.001 week−1 | 0.001 week−1 | ||
| DD above DV0 | DV0 | Lower temperature threshold | 15°C | 15°C | 315 |
| DV3 | Upper temperature threshold | 39°C | 39°C | 1683 | |
| DD above DVCS | DVCS a | Degree days based cold stress | 5.13°C | 5.13°C | a |
| DD above DVHS | DVHS | Degree days based heat stress | 39°C | 39°C | 1683 |
| Annual Heat sum | PDD b | Minimum degree day sum needed to complete a generation | 438.6 | 438.6 | b |
Values retrieved using the built‐in CLIMEX genetic algorithm (version 4.1.0.0, Kriticos et al., 2015).
PDD was calculated with a linear regression model using life table studies data (see section: Degree days and number of generations maps).
TABLE D.4. Estimated proportion of eggplant fruit infested with Leucinodes orbonalis discarded as household waste, i.e. by the end consumer.
| Question: | A transfer unit (aubergine fruits/grapes) is infested with L. orbonalis. Some transfer units will be discarded at the consumer level (private waste). What is the proportion of transfer units (aubergine fruits/grapes) that will be disposed at the consumer level? | ||||||
|---|---|---|---|---|---|---|---|
| Results | Proportion of infested transfer units discarded as household (private) waste | ||||||
| Percentiles: % | 1% | 5% | 25% | 50% | 75% | 95% | 99% |
| EKE estimates: | 20% | – | 35% | 50% | 60% | – | 75% |
| Fitted values: | 20.0% | 23.5% | 35.8% | 48.6% | 61.0% | 72.1% | 74.9% |
| Fitted distribution | BetaGeneral (1.3262, 1.2334, 0.185, 0.76) | ||||||
Information from CLIMEX informed estimates of the lag period (Appendix E) and impacts (Appendix F).
The majority of locations where L. orbonalis has been recorded from south Asia are in regions where EI exceeds 30 and a substantial number of records are from regions where EI is above 75 (Figure D.3.2). EI does not exceed 75 anywhere in the Euro‐Mediterranean region (Figure D.3.1) Light orange regions in Figure D.3.1. indicate potential areas for establishment EI ≥ 15 but < 30). However, such regions would be sub‐optimal for L. orbonalis. Darker orange regions (EI ≥ 30, < 75) indicate more favourable regions for establishment.
FIGURE D.3.
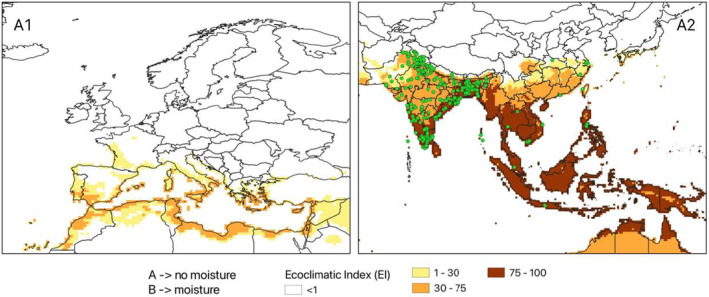
CLIMEX Ecoclimatic Index for Leucinodes orbonalis. 1. Euro‐Mediterranean region: 2 South Asia. EI range shown in five colour categories.
Area of establishment for Leucinodes orbonalis
Using the CLIMEX modelling system, Rossi et al. (2023) show that the majority of locations where L. orbonalis has been recorded from south Asia are in regions where EI exceeds 30 and a substantial number of records are from regions where EI is above 75. EI does not exceed 75 anywhere in the Euro‐Mediterranean region. There is uncertainty about the minimum threshold for establishment so two thresholds were used; EI 30 and EI 15. The table below indicate the area where EI ≥ 30 and EI ≥ 15 in NUTS 2 regions of the EU.
A total area of approximately 180,770 km2 of the EU has an EI ≥ 30 (Table D.2). Of this area, over 50% occurs in four NUTS 2 areas Andalucia (ES), Alentejo (PT) Sicily (IT) and Valencia (ES). Recognising the uncertainty of using an EI of 30 as a threshold, Table D.2 also shows the area of the EU where abiotic environmental factors considered within CLIMEX indicate where establishment may be possible if an EI threshold of 15 is used. In this case the total area suitable for establishment increases by a factor of approximately 2.5 to approximately 450,200 km2.
TABLE D.2.
Approximate area (thousands of km2) of NUTS2 regions of the EU where EI > 30 (where impacts thought most likely to occur) and where EI > 15 where establishment may be possible.
| A | B | C | D | E | F | G | H |
|---|---|---|---|---|---|---|---|
| NUTS2 code | NUTS 2 name | Area |
% of sum EI ≥ 30 |
Cumulative % | Area |
% of sum EI ≥ 15 |
Cumulative % |
| MS (name) | EI ≥30 | EI ≥ 15 | |||||
| ES61 | ES (Andalucía) | 47.5 | 26.28 | 26.28 | 67.5 | 14.99 | 14.99 |
| PT18 | PT (Alentejo) | 17.5 | 9.68 | 35.96 | 35 | 7.77 | 22.77 |
| ITG1 | IT (Sicilia) | 17.5 | 9.68 | 45.64 | 25 | 5.55 | 28.32 |
| ES52 | ES (Comunitat Valenciana) | 12.5 | 6.91 | 52.55 | 15 | 3.33 | 31.65 |
| EL43 | EL (Kriti) | 12.5 | 6.91 | 59.47 | 12.5 | 2.78 | 34.43 |
| CY00 | CY (Kýpros) | 10 | 5.53 | 65.00 | 10 | 2.22 | 36.65 |
| ES43 | ES (Extremadura) | 7.5 | 4.15 | 69.15 | 37.5 | 8.33 | 44.98 |
| ITF6 | IT (Calabria) | 7.5 | 4.15 | 73.30 | 15 | 3.33 | 48.31 |
| ITF4 | IT (Puglia) | 6.4 | 3.54 | 76.84 | 17.5 | 3.89 | 52.20 |
| EL42 | EL (Notio Aigaio) | 5.16 | 2.85 | 79.69 | 5.17 | 1.15 | 53.35 |
| ITG2 | IT (Sardegna) | 5 | 2.77 | 82.46 | 22.5 | 5.00 | 58.35 |
| EL41 | EL (Voreio Aigaio) | 5 | 2.77 | 85.22 | 5 | 1.11 | 59.46 |
| EL65 | EL (Peloponnisos) | 4.58 | 2.53 | 87.76 | 7.03 | 1.56 | 61.02 |
| ES53 | ES (Illes Balears) | 3.69 | 2.04 | 89.80 | 5.18 | 1.15 | 62.17 |
| ES62 | ES (Región de Murcia) | 3.31 | 1.83 | 91.63 | 10 | 2.22 | 64.39 |
| EL30 | EL (Attiki) | 3.02 | 1.67 | 93.30 | 3.88 | 0.86 | 65.25 |
| EL64 | EL (Sterea Elláda) | 1.96 | 1.08 | 94.39 | 5 | 1.11 | 66.36 |
| EL63 | EL (Dytiki Elláda) | 1.88 | 1.04 | 95.43 | 7.5 | 1.67 | 68.03 |
| ITI4 | IT (Lazio) | 1.43 | 0.79 | 96.22 | 12.5 | 2.78 | 70.80 |
| PT17 | PT (Área Metropolitana de Lisboa) | 1.36 | 0.75 | 96.97 | 5 | 1.11 | 71.91 |
| ES51 | ES (Cataluña) | 1.25 | 0.69 | 97.66 | 17.5 | 3.89 | 75.80 |
| ITF3 | IT (Campania) | 1.2 | 0.66 | 98.32 | 3.9 | 0.87 | 76.67 |
| ITF5 | IT (Basilicata) | 1.07 | 0.59 | 98.92 | 2.36 | 0.52 | 77.19 |
| EL62 | EL (Ionia Nisia) | 0.7 | 0.39 | 99.30 | 5 | 1.11 | 78.30 |
| PT20 | PT (Região Autónoma dos Açores) | 0.44 | 0.24 | 99.55 | 5 | 1.11 | 79.41 |
| MT00 | MT (Malta) | 0.31 | 0.17 | 99.72 | 0.31 | 0.07 | 79.48 |
| PT15 | PT (Algarve) | 0.25 | 0.14 | 99.86 | 7.5 | 1.67 | 81.15 |
| FRM0 | FR (Corse) | 0.22 | 0.12 | 99.98 | 5 | 1.11 | 82.26 |
| PT30 | PT (Região Autónoma da Madeira) | 0.04 | 0.02 | 100.00 | 0.76 | 0.17 | 82.43 |
| PT16 | PT (Centro (PT)) | – | – | 20 | 4.44 | 86.87 | |
| FRI1 | FR (Aquitaine) | – | – | 12.5 | 2.78 | 89.65 | |
| ES24 | ES (Aragón) | – | – | 10 | 2.22 | 91.87 | |
| FRL0 | FR (Provence‐Alpes‐Côte d'Azur) | – | – | 7.5 | 1.67 | 93.53 | |
| FRJ1 | FR (Languedoc‐Roussillon) | – | – | 5 | 1.11 | 94.64 | |
| ITI1 | IT (Toscana) | – | – | 5 | 1.11 | 95.76 | |
| PT11 | PT (Norte) | – | – | 3.73 | 0.83 | 96.58 | |
| ES42 | ES (Castilla‐La Mancha) | – | – | 3.06 | 0.68 | 97.26 | |
| EL54 | EL (Ipeiros) | – | – | 2.67 | 0.59 | 97.86 | |
| HR03 | HR (Jadranska Hrvatska) | – | – | 2.25 | 0.50 | 98.36 | |
| ES11 | ES (Galicia) | – | – | 2.17 | 0.48 | 98.84 | |
| EL52 | EL (Kentriki Makedonia) | – | – | 1.45 | 0.32 | 99.16 | |
| ITC3 | IT (Liguria) | – | – | 0.98 | 0.22 | 99.38 | |
| FRJ2 | FR (Midi‐Pyrénées) | – | – | 0.87 | 0.19 | 99.57 | |
| ITF1 | IT (Abruzzo) | – | – | 0.66 | 0.15 | 99.72 | |
| ES12 | ES (Principado de Asturias) | – | – | 0.52 | 0.12 | 99.83 | |
| ITF2 | IT (Molise) | – | – | 0.45 | 0.10 | 99.93 | |
| EL61 | EL (Thessalia) | – | – | 0.21 | 0.05 | 99.98 | |
| ITI2 | IT (Umbria) | – | – | 0.09 | 0.02 | 100.00 | |
| Sum | 180.77 | 100 | 450.2 | 100.00 |
Introduction model: Allocation to NUTS regions
Based on the results of CLIMEX shown in Rossi et al. (2023), the pathway model for entry detailed in Appendix C was followed up with a step for the within‐EU distribution of infested host fruit, the allocation to areas with Eco‐climatic Index (EI) ≥ 30, and ≥ 15 to reflect the uncertainty regarding the EI threshold for establishment. Following arrival in the EU, it is assumed that the produce is distributed across EU NUTS regions in proportion to the human population. Based on Eurostat reports of population in each NUTS2 area, 23.12% of the EU population are in NUTS2 regions in which EI ≥ 15 and 14.09% are in regions where EI ≥ 30.
Introduction model: Proportion of fruit discarded
An important factor to consider is that for larvae to complete their development, the contaminated eggplant fruit they arrive in should be discarded before it is processed, e.g. cooked and consumed.
At this point in the pathway for introduction, the density of infested eggplant fruit is thought to be very low and only eggplants discarded together by importers, wholesalers or retailers will be of sufficient quantity for there to be a reasonable chance that a male and female may emerge in sufficiently close proximity for them to find each other and mate to potentially initiated a founder population. Eggplant is a sensitive fruit, and discards could be due to several reasons, such as damage during handling and transport, physical quality problems, market conditions and pest finds.
Commercial waste of infested eggplant
TABLE D.3. Estimated proportion of eggplant fruit infested with Leucinodes orbonalis discarded as commercial waste, i.e. between import and consumer.
| Question: | A transfer unit (aubergine fruits/grapes) is infested with L. orbonalis. Some transfer units will be discarded between import and consumer (commercial waste). What is the proportion of transfer units (aubergine fruits/grapes) that will be disposed between import and consumer? | ||||||
|---|---|---|---|---|---|---|---|
| Results | Proportion of infested transfer units discarded as commercial waste | ||||||
| Percentiles: % | 1% | 5% | 25% | 50% | 75% | 95% | 99% |
| EKE estimates: | 5% | – | 9% | 12% | 15% | – | 30% |
| Fitted values: | 4.99% | 6.28% | 9.10% | 11.8% | 15.2% | 21.1% | 26.0% |
| Fitted distribution | BetaGeneral (3.5888, 33.705, 0.032, 1) | ||||||
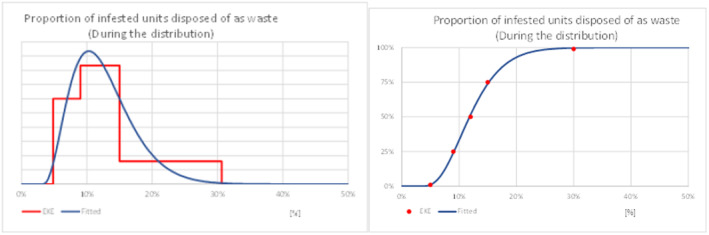
FIGURE D.4. Distribution of proportion of infested eggplants disposed of as waste between import and consumer fitted to EKE estimates (left hand chart shows probability density function to describe the remaining uncertainties of the parameter; right hand chart shows cumulative distribution function (CDF) of the likelihood of the parameter).
The median proportion of transfer units (eggplant fruits/grapes) infested with L. orbonalis and discarded after import but before reaching the consumer was estimated to be 11.8% (90% CR from 6.28% to 21.1%).
Household waste of infested eggplant
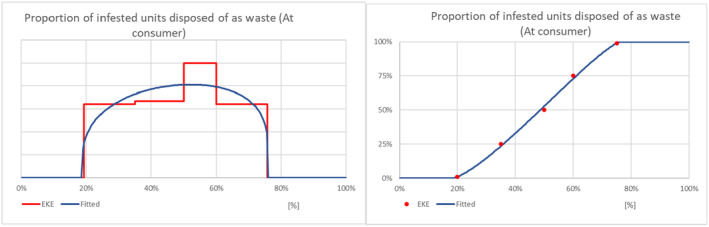
FIGURE D.5. Distribution of proportion of infested eggplants disposed of at the consumer level as household waste fitted to EKE estimates (left hand chart shows probability density function to describe the remaining uncertainties of the parameter; right hand chart shows cumulative distribution function (CDF) of the likelihood of the parameter).
The median proportion of transfer units (eggplant fruits/grapes) infested with L. orbonalis and discarded after import but before reaching the consumer was estimated to be 48.6% (90% CR from 23.5% to 72.1%).
Reasoning
Infested eggplants are very likely detected by the end consumer.
TABLE D.5. Uncertainties associated with low and high discard proportion scenarios.
| Uncertainty | Low discard proportion scenario | High discard proportion scenario |
|---|---|---|
| Detection of infestation | Importers, wholesalers and retailers are less likely to detect infestations from inspecting the fruit externally | End consumers process the fruit by cutting them open, resulting in a high detection and disposal rate |
| Quality standards | Low standards, businesses want to sell as many fruit as possible | High standards, businesses want to sell as high a quality product as possible, resulting in the sorting and disposal of unsatisfactory fruit |
| Processing of fruit | Small infested fruit may be cooked uncut | Large fruit are cut open, and infestation will be detected |
Lower limit:
Eggplant is an expensive fruit, and importers, wholesalers and retailers want to sell as many fruit as possible.
Households will want to use as much of the expensive fruits as possible.
Infestation of turkey berries may be overlooked when cooking the berries uncut.
Upper limit:
High proportion of disposal by importers, wholesalers and retailers to remove unsatisfactory fruit due to transport and handling damage or problems with physical quality;
The majority of infested fruit are detected by the end consumer and discarded.
Median:
Infestation of fruit is difficult to detect between import and end consumer, i.e. before the fruit are processed for cooking.
While cutting the fruit for cooking, end consumers will detect infestation.
Cutting the fruit for cooking may kill larvae feeding inside.
Inter‐quartile range:
High uncertainties below and above the median
Introduction model: Probability of larvae surviving to develop to adulthood then escape from discarded waste
L. orbonalis larvae exit the fruit prior to pupation (Navasero & Calilung, 1990; Saxena, 1965).
Eggs are laid on leaves and fruit, but larvae hatch within 3–4 days after egg deposition (Navasero & Calilung, 1990; Saxena, 1965), which is assumed to take place before a consignment of eggplant arrives in the EU; by the time of arrival, larvae hatching from eggs laid on fruit will have hatched and bored into the fruit.
Larval development (incl. egg phase) takes about 11–29 days, pupal development (incl. prepupal phase) 10–11 days at 21 C (Table B.1, Figure B.1; Navasero & Calilung, 1990; Saxena, 1965).
From the fourth larval instar on, when disturbed or the fruit is rotting, the larva can prematurely enter the prepupal phase by constructing the pupation cocoon, in which it passes the remaining larval instars without feeding until it pupates (‘cocooning behaviour’; Saxena, 1965).
TABLE D.6. Estimated probability of an adult Leucinodes orbonalis emerging and escaping from eggplant fruit discarded with commercial waste (excluding household waste).
| Question: | What is the probability that a larva in the discard waste will result in an adult emerging from the commercial waste? | ||||||
|---|---|---|---|---|---|---|---|
| Results | Likelihood that an adult will emerge from discarded eggplant commercial waste | ||||||
| Percentiles: % | 1% | 5% | 25% | 50% | 75% | 95% | 99% |
| EKE estimates: | 0.1% | – | 0.6% | 1% | 1.5% | – | 2% |
| Fitted values: | 0.100% | 0.195% | 0.582% | 1.03% | 1.48% | 1.90% | 2.01% |
| Fitted distribution | BetaGeneral (1.1703, 1.2288, 0.00068, 0.0205) | ||||||
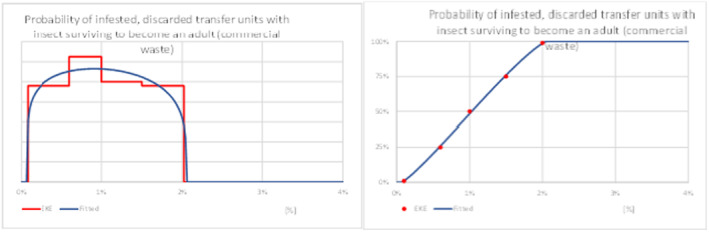
FIGURE D.6. Distribution of probability of an adult emerging from infested eggplant fruit discarded with commercial waste fitted to EKE estimates (left hand chart shows probability density function to describe the remaining uncertainties of the parameter; right hand chart shows cumulative distribution function (CDF) of the likelihood of the parameter).
The median likelihood that a larva discarded in commercial waste (i.e. prior to reaching the end consumer) will continue its development to emerge as an adult that escapes from waste was estimated to be 1.03% (90% CR from 0.195% to 1.90%).
The median likelihood that a larva discarded in household waste (i.e. after the supply chain when the fruit have reached the end consumer) will continue its development to emerge as an adult that escapes from waste was estimated to be 5.13% (90% CR from 0.98% to 12.22%).
TABLE D.7. Estimated probability of an adult Leucinodes orbonalis emerging and escaping from eggplant fruit discarded with household waste (excluding commercial waste).
| Question: | What is probability that a larva in the discard waste will result in an adult emerging from the household waste? | ||||||
|---|---|---|---|---|---|---|---|
| Results | Likelihood that an adult will emerge from discarded eggplant household waste | ||||||
| Percentiles: % | 1% | 5% | 25% | 50% | 75% | 95% | 99% |
| EKE estimates: | 0% | – | 3% | 5% | 8% | – | 15% |
| Fitted values: | 0.361% | 0.98% | 2.92% | 5.13% | 7.89% | 12.2% | 15.05% |
| Fitted distribution | BetaGeneral (1.6704, 4.5179, 0, 0.21) | ||||||
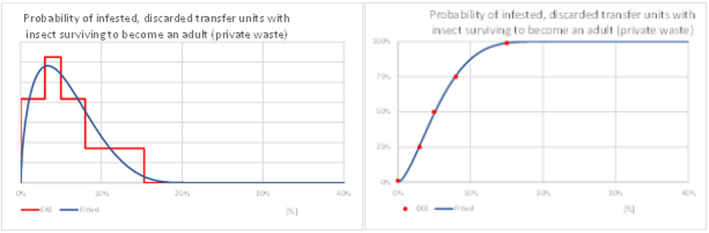
FIGURE D.7. Distribution of probability of an adult emerging from infested eggplant fruit discarded with household waste fitted to EKE estimates (left hand chart shows probability density function to describe the remaining uncertainties of the parameter; right hand chart shows cumulative distribution function (CDF) of the likelihood of the parameter).
Reasoning
Eggplant fruit discarded between import and consumer (commercial waste) likely end up in landfills or organic waste processing plants, where survival of the larva is unlikely.
A considerable amount of eggplant fruit discarded after arriving at the end consumer (household waste) is discarded on composts, where survival of the larva to adulthood is favourable.
From the fourth instar on, larvae can complete their life cycle in the prematurely constructed pupation cocoon without feeding (Saxena, 1965).
TABLE D.8. Uncertainties associated with low and high emergence probability scenarios.
| Uncertainty | Low emergence probability scenario | High emergence probability scenario |
|---|---|---|
| Waste handling | Low proportion of composting, primarily waste processing and/or landfills | High proportion of composting |
| Developmental stage of larva | After disturbance, early instar larvae (1st to 3rd instar) may leave the fruit and starve in lack of alternative fruit to feed on | Late instar larvae (from 4th instar on) can pass the remaining larval stages in the pupal cocoon without feeding |
Lower limit:
Low proportion of compost disposal as household waste in urban areas, where composting options are limited;
Majority of discarded fruit go to landfills, where they are quickly covered by additional waste, trapping or killing the developing larva.
Upper limit:
High probability of disposal by end consumer on compost;
Household waste primarily containing late instar larvae that can survive to adulthood without food.
Median:
Mix of waste disposal (landfills, waste processing, composting).
Inter‐quartile range:
High uncertainties below and above the median
Introduction model: Proportion of adults mating
An important limiting factor in establishing a founder population is the likelihood of a male and a female emerging in temporal and spatial proximity to locate each other and mate. Recall that border interceptions of insects are a poor predictor of successful establishment (Kenis et al., 2007; Caley et al., 2015).
Panel adopted the probabilities of adult mating that were elicited for Elasmopalpus lignosellus (EFSA PLH Panel, 2023), but the actual probabilities are presumably even lower. One reason for this is the shorter adult lifespan of L. orbonalis, with males living 4–5 days and quickly dying after copulation, whereas females live 7 to maximally 8 days (Navasero & Calilung, 1990; Saxena, 1965). The time window for females and males to meet and mate is therefore narrower compared to E. lignosellus.
TABLE D.9. Estimated probability that an adult female of Leucinodes orbonalis will mate, as elicited for Elasmopalpus lignosellus (EFSA PLH Panel, 2023).
| Question: | What is the probability that an escaped adult female will find a mate either before flying off to find a host or at a host? | ||||||
|---|---|---|---|---|---|---|---|
| Results | Proportion of females successfully mating | ||||||
| Percentiles: % | 1% | 5% | 25% | 50% | 75% | 95% | 99% |
| EKE estimates | 0.00% | – | 0.05% | 0.08% | 0.12% | – | 0.20% |
| Fitted values | 0.0071% | 0.018% | 0.049% | 0.081% | 0.12% | 0.17% | 0.20% |
| Fitted distribution | BetaGeneral (1.8097, 3.2291, 0, 0.0024) | ||||||
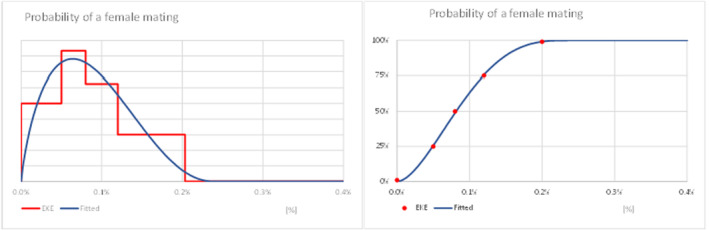
FIGURE D.8. Distribution of probability of a female successfully mating fitted to EKE estimates (left hand chart shows probability density function to describe the remaining uncertainties of the parameter; right hand chart shows cumulative distribution function (CDF) of the likelihood of the parameter).
The median likelihood that an escaped adult female will successfully mate was estimated to be 0.081% (i.e. 8 in 10,000) (90% CR from 0.018% to 0.17%, approximately 2 in 10,000 to 17 in 10,000).
Reasoning
Short range of attraction of the female sex pheromone, with a maximum range of 10 m (Prasad et al., 2005);
Shorter adult life span of L. orbonalis (males 4–5 days, females 7–8 days; Saxena, 1965; Navasero & Calilung, 1990) compared to E. lignosellus (9–13 days, up to 20 days under lab conditions; EFSA PLH Panel, 2023), creating a shorter time window for mate finding and mating.
TABLE D.10. Uncertainties associated with low and high mating success scenarios.
| Uncertainty | Low mating success scenario | High mating success scenario |
|---|---|---|
| Adult life span | L. orbonalis adults more short‐lived than those of E. lignosellus | |
| Mate attraction | Short range of sex pheromone attraction | Female sex pheromone is strongly attracting males |
| Synchronisation of adult emergence | Few or no adults of opposite sex emerge in temporal and spatial proximity | Adult emergence from pupae is well synchronised |
Lower limit:
Only 3–4 days after adult emergence available for mate finding and mating;
Female sex pheromone is strongest in young female moths, loses attractiveness/potency with time.
Upper limit:
Female sex pheromone is very attractive to males (an artificial version is used for mass trapping of L. orbonalis males in pest management).
Median:
Effective range and potency of the sex pheromones of L. orbonalis and E. lignosellus are comparable.
Inter‐quartile range:
High uncertainties below and above the median.
Introduction model: Likelihood of founder population initiation
Leucinodes orbonalis is an oligophagous pest and feeds in the shoots and fruits of different species of Solanum in the plant family of nightshades (Solanaceae). The Mediterranean region is home to 48 species of Solanum (incl. lycopersicum; Valdés, 2012) (see table below with the presence recorded at the MS level from Valdes 2012 database online accessed on 28/11/2023), all serving as potential host plants for L. orbonalis larvae. Several European Solanum species are widely distributed weeds, such as S. nigrum, ‘a common weed of cultivated land, open spaces in forests and roadsides [that is] found in disturbed areas between 0 and 2200 (3500) m elevation;’ (Särkinen et al., 2023). As such, L. orbonalis has a good chance of finding a suitable host within areas suitable for establishment.
| AT | BE and LU | BG | HR | CY | CZ | DK | EE | FI | FR | DE | GR | NL | ES | HU | IE | IT | LV | LT | PL | PT | RO | MT | SK | SI | SE | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| S. abutiloides | x | |||||||||||||||||||||||||
| S. alatum | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | |||||||
| S. americanum | x | x | x | x | ||||||||||||||||||||||
| S. aviculare | x | |||||||||||||||||||||||||
| S. bonariense | x | x | x | |||||||||||||||||||||||
| S. capsicoides | x | |||||||||||||||||||||||||
| S. carolinense | x | x | x | |||||||||||||||||||||||
| S. chenopodioides | x | x | x | x | x | |||||||||||||||||||||
| S. chrysotrichum | x | |||||||||||||||||||||||||
| S. citrullifolium | x | x | x | |||||||||||||||||||||||
| S. dulcamara | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | ||
| S. eleagnifolium | x | x | x | x | x | x | x | |||||||||||||||||||
| S. fastigiatum | x | |||||||||||||||||||||||||
| S. giganteum | x | |||||||||||||||||||||||||
| S. gracile | x | |||||||||||||||||||||||||
| S. heterodoxum | x | x | x | |||||||||||||||||||||||
| S. hispidum | x | |||||||||||||||||||||||||
| S. laciniatum | x | x | ||||||||||||||||||||||||
| S. laxum | x | x | x | |||||||||||||||||||||||
| S. lidii | x | |||||||||||||||||||||||||
| S. linnaeanum | x | x | x | x | x | x | x | x | x | |||||||||||||||||
| S. lycopersicum | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | ||||
| S. marginatum | x | x | ||||||||||||||||||||||||
| S. mauritanium | x | x | ||||||||||||||||||||||||
| S. melanocerasum | x | x | x | |||||||||||||||||||||||
| S. melongena | x | x | x | x | x | x | x | x | x | |||||||||||||||||
| S. microcarpum | x | |||||||||||||||||||||||||
| S. nava | x | |||||||||||||||||||||||||
| S. nigrum | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x |
| S. patens | x | |||||||||||||||||||||||||
| S. physalifolium | x | x | x | x | x | x | ||||||||||||||||||||
| S. pseudocapsicum | x | x | x | x | x | x | ||||||||||||||||||||
| S. pygmaeum | x | |||||||||||||||||||||||||
| S. retroflexum | x | |||||||||||||||||||||||||
| S. robustum | x | |||||||||||||||||||||||||
| S. rostratum | x | x | x | x | x | x | x | x | x | x | x | x | x | |||||||||||||
| S. sarachoides | x | x | x | x | x | x | x | x | ||||||||||||||||||
| S. scabrum | x | x | ||||||||||||||||||||||||
| S. seaforthianum | x | |||||||||||||||||||||||||
| S. sisymbrifolium | x | x | x | x | x | x | x | x | ||||||||||||||||||
| S. torvum | x | |||||||||||||||||||||||||
| S. triflorum | x | x | x | x | x | x | x | |||||||||||||||||||
| S. trisectum | x | |||||||||||||||||||||||||
| S. tuberosum | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | ||||||
| S. vespertilio | x | |||||||||||||||||||||||||
| S. viarum | x | |||||||||||||||||||||||||
| S. villosum | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | x | ||
| S. virgatum | x |
The female is carrying up to 300 eggs that are laid singly on the leaves and fruit of host plants (Navasero & Calilung, 1990). Specialised predators and parasitoids are expected to be absent in a potential founder population. Due to the internal feeding of the larvae and the closing of the entry hole into the plant with a plug of excreta, L. orbonalis is not very susceptible to generalist predators.
Once the next generation emerges, males and females are likely to be present at the same location at a similar time, although the adult stage is short‐lived, with 4–5 days for males and about 7 days for females (Navasero & Calilung, 1990; Saxena, 1965). A potent (though apparently short‐ranged) female sex pheromone increases the likelihood of males and females meeting to mate (Prasad et al., 2005). Host plants in suitable stages are also likely to be found. In unfavourable conditions during winter, L. orbonalis is able to hibernate in the pupal stage (Lal, 1975). These factors promote the probability of a mated female founding a population that would persist. This probability was therefore estimated to not be small, with an elicited lower bound of 2.41% of mated females founding a persistent population and the upper bound 31.6% (Table D.11).
TABLE D.11.
Estimated probability that a founder population of Leucinodes orbonalis will be initiated following successful mating.
| Question: | What is the proportion of mated females of Leucinodes orbonalis that will successfully establish a founder population? | ||||||
|---|---|---|---|---|---|---|---|
| Results | Likelihood that a founder population will be initiated following successful mating | ||||||
| Percentiles: % | 1% | 5% | 25% | 50% | 75% | 95% | 99% |
| EKE estimates | 1% | – | 9% | 17% | 25% | – | 33% |
| Fitted values | 1.00% | 2.41% | 9.00% | 17.0% | 25.0% | 31.6% | 33.0% |
| Fitted distribution | BetaGeneral (1.0456, 1.0456, 0.0062, 0.3338) | ||||||
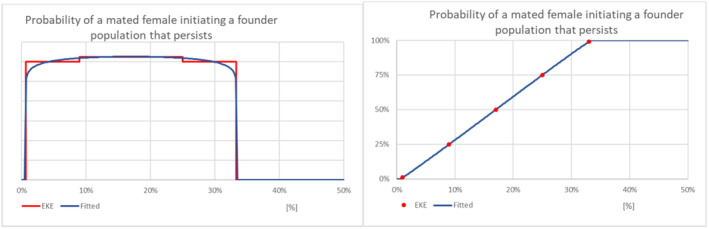
FIGURE D.9. Distribution of probability of a founder population being initiated following successful mating fitted to EKE estimates (left hand chart shows probability density function to describe the remaining uncertainties of the parameter; right hand chart shows cumulative distribution function (CDF) of the likelihood of the parameter).
The median likelihood that a founder population will be initiated following successful mating was estimated to be 17.0% (90% CR from 2.41% to 31.6%).
Reasoning
Gravid females of L. orbonalis and E. lignosellus lay similar amounts of eggs singly or in small clusters on or near the host plant.
Larvae of both species are internal borers and are thus fairly well protected from parasitoids and predators.
TABLE D.12. Uncertainties associated with low and high founder population establishment scenarios.
| Uncertainty | Low establishment scenario | High establishment scenario |
|---|---|---|
| Host plant availability | Lower host plant availability compared to E. lignosellus | Several Solanum weed species present in regions suitable for L. orbonalis establishment |
| Natural enemies | Higher biological control through natural enemies compared to E. lignosellus; eggs and especially young larvae more exposed to parasitation and predation as compared to E. lignosellus, whose larvae build a shelter in the upper soil level | Similar natural enemy pressure as E. lignosellus; within a few minutes after hatching, young larvae bore into the host plant and plug the entry hole with excreta |
Lower limit:
L. orbonalis only has Solanum spp. available as potential host plants, compared to the polyphagous E. lignosellus that feeds on plants of several plant families.
Upper limit:
L. orbonalis is oligophagous on several Solanum species, several of which are widely distributed weed species on cultivated and disturbed land.
Median:
As a newly established species, biological control of L. orbonalis through natural enemies (parasitoids, predators) is likely comparable to that of E. lignosellus in the same situation.
The females of both L. orbonalis and E. lignosellus produce similar amounts of eggs, and both species lay their eggs singly or in small clusters, thus spreading the risk of parasitisation and mortality.
Inter‐quartile range:
High uncertainties below and above the median
APPENDIX E. Spread
E.1.
To inform the assessment of spread, the Panel first estimated the duration of the lag phase before estimating the linear rate of range expansion during the phase in which spread is at its fastest.
No information is available on the natural spread capacities or flight capabilities of L. orbonalis. Therefore, to aid our estimation of spread of L. orbonalis after its establishment in the EU, we analysed spread data of a related African species, L. laisalis (Guenée, 1854), which was first reported from the southern Iberian Peninsula in 1958 and has since then been spreading in this region (Figure E.1). Leucinodes laisalis adults (Figure E.1) are similar in size to L. orbonalis, with a forewing length of 7.0–11.5 mm and the females being somewhat larger than the males (Mally et al., 2015). From this, we assume that both Leucinodes species have similar spread rates based on their presumably comparable flight capability.
FIGURE E.1.
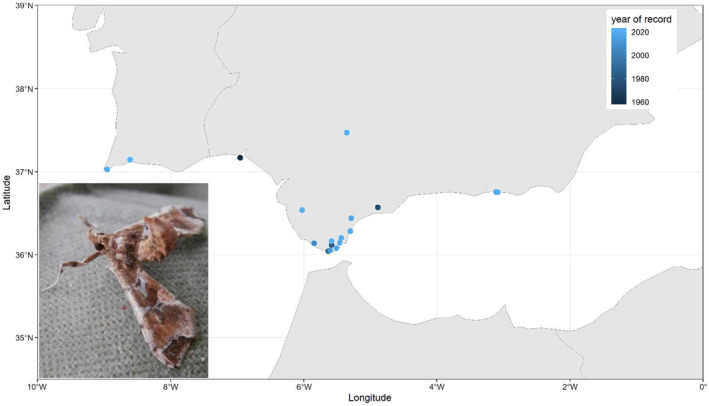
Map of reported occurrences of Leucinodes laisalis (Guenée, 1854) on the southern Iberian Peninsula between 1958 (first record) and 2023; data compiled from GBIF, Lepiforum, iNaturalist and Mally et al. (2015). Photo: Female adult L. laisalis, Spain, Andalusia, 5 km N of Terifa, 2014‐09‐22, det. & phot. © Stephen McAvoy, map © Richard Mally.
Occurrence data of L. laisalis on the Iberian Peninsula were compiled from different sources (GBIF.org, 2023; Huertas‐Dionisio, 2000; iNaturalist, 2023; Lepiforum, 2023; Mally et al., 2015) and mapped in R. The data show that L. laisalis has been repeatedly reported in southern Spain and Portugal since 1958 (Figure E.1). Leucinodes laisalis appears to be established in the southernmost Iberian Peninsula and is slowly spreading at an average rate of approximately 1.6 km/year based on a linear regression of the available data (Figure E.2). Taking only the records into account that are consecutively furthest away from the first record, and assuming linear spread, an annual spread rate of 5.51–11.51 km/year is assessed for L. laisalis (Table E.1).
FIGURE E.2.
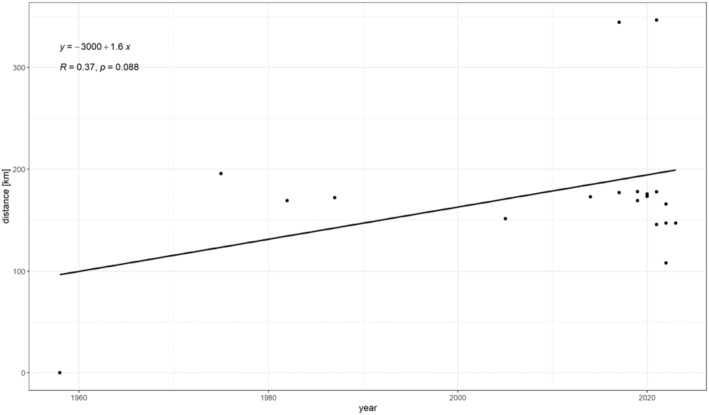
Scatterplot of the spread of Leucinodes laisalis in Europe between 1958 and 2023.
TABLE E.1.
Years of record of Leucinodes laisalis in Spain and Portugal, geographic distance from the first record point in 1958 and annual spread rate calculated for the three furthest consecutive distances.
| Year of record | Distance from first record (km) | Annual spread based on furthest consecutive record (km/a) |
|---|---|---|
| 1958 | 0 | |
| 1975 | 195.63 | 195.63/(1975–1958) = 11.51 |
| 1982 | 169.18 | |
| 1987 | 172.00 | |
| 2005 | 151.42 | |
| 2014 | 172.79 | |
| 2017 | 176.97 | |
| 2017 | 344.82 | 344.82/(2017–1958) = 5.84 |
| 2019 | 178.02 | |
| 2019 | 169.15 | |
| 2020 | 175.61 | |
| 2020 | 173.17 | |
| 2021 | 145.74 | |
| 2021 | 347.06 | 347.06/(2021–1958) = 5.51 |
| 2021 | 347.06 | |
| 2021 | 177.83 | |
| 2022 | 165.79 | |
| 2022 | 165.79 | |
| 2022 | 147.15 | |
| 2022 | 147.15 | |
| 2023 | 195.63 | |
| 2023 | 169.18 |
E.1. Estimated duration of the lag phase
Table E.2 shows EKE estimates for the duration of the lag phase based on the evidence summarised as bullet points below.
TABLE E.2.
Estimated duration of the lag phase of Leucinodes orbonalis (in years).
| Question | What is the duration of the lag phase, this means the time between first establishment and the continuous expansion of the infested area (linear phase)? | ||||||
|---|---|---|---|---|---|---|---|
| Results | How long is the average duration of the lag phase? (years) | ||||||
| Percentiles: % | 1% | 5% | 25% | 50% | 75% | 95% | 99% |
| EKE estimates | 4 | – | 17 | 30 | 65 | – | 100 |
| Fitted values (years) | 4.00 | 4.85 | 14.49 | 34.5 | 62.3 | 92.2 | 100.00 |
| Fitted distribution | BetaGeneral (0.666, 1.1588, 3.92, 103) | ||||||
FIGURE E.3. Distribution of the estimated mean duration of the lag phase of L. orbonalis (years) fitted to EKE estimates (left hand chart shows probability density function to describe the remaining uncertainties of the parameter; right hand chart shows cumulative distribution function (CDF) of the likelihood of the parameter).
FIGURE E.4. Distribution of the estimated mean spread rate of L. orbonalis (km/year) fitted to EKE estimates (Left hand chart shows probability density function to describe the remaining uncertainties of the parameter; right hand chart shows cumulative distribution function (CDF) of the likelihood of the parameter).
The median estimate for the duration of the lag phase was 4.5 years (90% CR from 4.85 to 92.2 years).
Reasoning
The occurrence and spread of the related species L. laisalis in southern Spain and Portugal were reviewed regarding the specific development at the beginning of the spread;
The spread behaviour of L. orbonalis is frequently described as stationary;
The lag phase is largely driven by the rate of population growth;
Fecundity: about 300 eggs per mated female;
Population development will depend on the availability and condition (e.g. state of nutrition) of host plants;
Population development will be faster on Solanum crops;
Host plant range includes wild plants such as Solanum torvum, S. nigrum, S. aethiopicum;
Mate attraction based on female sex pheromones with a short maximum range (10 m).
TABLE E.3. Uncertainties associated with short and long lag phase scenarios.
| Uncertainty | Short lag phase scenario | Long lag phase scenario |
|---|---|---|
| Life cycles | 4–6 generations per year | 1 generation per year |
| Host | Continuous host availability; Solanum crops | Host plants die during winter; Solanum weeds |
| Population dynamics | Good conditions for development, incl. Crop monoculture, bad sanitary habits, staggered planting | Poor conditions of development, esp. less preferred hosts available, wild environment, fewer fruit/biomass due to fewer nutrients |
Lower limit:
Favourable climatic conditions with up to six generations per year and continuous host plant availability;
Outbreaks in crop monocultures with poor phytosanitary measures, hygienic conditions and staggered planting.
Upper limit:
Unfavourable climatic conditions with only one generation per year and host plants dying off during winter;
Outbreaks in natural environment, e.g. on Solanum weeds (Solanum nigrum) with poor nutritional conditions;
Pest is described as stationary.
Median:
Good developmental conditions (agricultural areas with suitable crops) are more likely in the climatic suitable areas of the EU.
Over the years, Leucinodes orbonalis will adapt to poor conditions, which allows for better development.
Inter‐quartile range:
The lack of specific evidence indicates maximum uncertainties below/above the median.
E.2. Estimated linear rate of range expansion (spread)
Table E.4 shows the EKE estimates for the linear rate of spread based on the evidence summarised as bullet points below.
TABLE E.4.
Estimated rate of linear spread of Leucinodes orbonalis (km/year).
| Question | What is the annual median rate of spread when the established population is spreading at a constant rate? | ||||||
|---|---|---|---|---|---|---|---|
| Results | |||||||
| Percentiles: % | 1% | 5% | 25% | 50% | 75% | 95% | 99% |
| EKE estimates | 0.5 | – | 1.5 | 2.0 | 4.0 | – | 10.0 |
| Fitted values (km/year) | 0.50 | 0.652 | 1.31 | 2.28 | 3.80 | 7.02 | 10.0 |
| Fitted distribution | BetaGeneral (1.2452, 34.666, 0.445, 70) | ||||||
The median annual rate of spread for Leucinodes orbonalis was estimated to be 2.28 km per year (90% CR from 0.652 to 7.02 km per year) during the period of constant rate of spread.
Reasoning
The spread rate is the outcome of the contribution of natural dispersal together with local human‐assisted spread;
Spread due to post‐harvest movement, such as the trade in commodities, was not included in the estimation;
There is no quantitative data on the flight capability of L. orbonalis;
Adults are weak fliers (Chang et al., 2014; Dr Ramasamy, personal communication);
The adults can move from field to field and hence can spread over an area over the course of several generations; maximum estimated field to field spread is 10 km in a single year (comprising several generations; see next bullet point) (Dr Ramasamy, personal communication);
To complete its life cycle (i.e. one generation), L. orbonalis requires 26–36 days in the tropics (Navasero & Calilung, 1990) and 26–39 days in the subtropics (Lall & Ahmad, 1965), which allows for 10–14 generations per year; under Mediterranean European conditions, the number of expected generations per year is significantly lower;
Jethva and Vyas (2009) extrapolated from lab studies of L. orbonalis reared on tomato that the population would multiply by a factor of 2.358 per week.
TABLE E.5. Uncertainties associated with slow and faster spread rate scenarios.
| Uncertainty | Slower spread rate scenario | Faster spread rate scenario |
|---|---|---|
| Flight capability extrapolated from the related L. laisalis | Similar flight capacity of the two Leucinodes spp. assumed | |
| Range expansion extrapolation for L. laisalis | Average spread rate (linear model): 1.6 km/year | Maximum spread rate: 5.5–11.5 km/year; calculated average rate might include the lag phase, with the actual average rate being higher |
| Larval host plants | L. orbonalis spreads as fast as L. laisalis due to similar hosts | L. orbonalis spreads faster than L. laisalis due to broader host range and higher host plant density, with S. melongena as primary host |
| Plant production cycle | Eggplants not grown year‐round, therefore lower number of L. orbonalis generations per year in EU as compared to India (where a spread of 10 km/year is estimated) | Host plants grown/available throughout the year |
| Plant production conditions | Low availability of host plant (eggplant) due to managed cultivation, especially in protected conditions | Unmanaged, unprotected wild hosts available |
| Climatic suitability | The primarily tropical L. orbonalis is less well adapted to Mediterranean climate than L. laisalis, thus slower life cycle, reproduction and consequently spread | L. orbonalis also occurs in regions with occasional winter frost (Lal, 1975), indicating a relatively broad climatic tolerance |
Lower limit:
Spread rate for L. laisalis is average not lower limit.
L. laisalis is more adapted to the Mediterranean climate, whereas L. orbonalis is more adapted to the tropics and subtropics.
Main crop production is on small, scattered plots.
Upper limit:
L. orbonalis has larger host range with higher plant density in EU.
Lower host plant availability due to managed cultivation reduces the spread, esp. in protected conditions, but hosts in natural environment exist.
Spread is expected to be reduced compared to Asia.
Median:
Lower suitability for EU climate than L. laisalis.
Inter‐quartile range:
High uncertainty below the median;
Medium uncertainty above the median.
Singh et al. (2007) made catches of L. orbonalis in a landscape with only a single plot of brinjal (Solanum melongena) at different distances from the brinjal plot. Traps located at 50 and 100 m from brinjal field attracted fewer male L. orbonalis moths than those at 0, 150 and 350 m, indicating the feasibility of trapping male Leucinodes orbonalis moths even in a non‐brinjal area. Trap direction did not significantly influence trap catch. Nearly 60% of male L. orbonalis moths were observed in traps placed against direction of the wind.
APPENDIX F. Impact
F.1.
The Panel focused on the impact on eggplant, as it is evident from the large body of literature (467 studies; Table C.1) that this crop is heavily impacted by L. orbonalis in Asia, and vice versa that L. orbonalis prefers eggplant as host above all other solanaceous crops (Ardez et al., 2008). The Panel based its assessment on several Asian studies investigating impact of L. orbonalis on eggplant, which were compiled in meta‐analyses (see F.3–F.5 below).
F.1. Estimated impact with no pest management practice in place
This is an artificial scenario for a commercial farmer but can be seen in experimental trials where pest management practices are excluded.
Table F.1 shows EKE estimates for yield loss without specific control measurements against L. orbonalis, informed by a meta‐analysis of losses (below) reported from control plots in pesticide trials and on the evidence summarised as bullet points below.
TABLE F.1.
Estimated mean reduction in yield (yield loss) caused by L. orbonalis in the absence of pest control.
| Question | What is the likely mean reduction of annual yield of eggplant, grown outdoors in a southern European country, with EI higher than 30 without specific pest control measures? | ||||||
|---|---|---|---|---|---|---|---|
| Results | Annual eggplant yield loss without control measures | ||||||
| Percentiles: % | 1% | 5% | 25% | 50% | 75% | 95% | 99% |
| EKE estimates | 0.5% | – | 2.0% | 4.5% | 8.0% | – | 15.0% |
| Fitted values (% losses) | 0.504% | 0.670% | 2.01% | 4.47% | 8.03% | 13.0% | 15.1% |
| Fitted distribution | BetaGeneral (0.78643, 1.8133, 0.0048, 0.166) | ||||||
FIGURE F.1. Distribution of mean reduction of annual eggplant yield without control measures fitted to EKE estimates. (Left hand chart shows probability density function to describe the remaining uncertainties of the parameter; right hand chart shows cumulative distribution function (CDF) of the likelihood of the parameter).
The median annual yield loss in eggplant crops was estimated to be 4.47% (90% CR from 0.67% to 13.0%) in a scenario where no pest management is in place.
Reasoning
No specific measures taken for L. orbonalis but insecticides might be (but not necessarily) used against other Lepidoptera.
TABLE F.2. Uncertainties associated with low and high yield loss scenarios.
| Uncertainty | Low yield loss scenario | High yield loss scenario |
|---|---|---|
| Agricultural practices | Good agricultural practices, e.g. crop rotation, sanitation, clean planting material, regular inspections by grower | Amateur gardens. Small scale. No pesticides. Staggered planting, some greenhouses, but not sanitated, so near‐Indian opportunities for field‐to‐field movement of the insect and perpetuation. Lack of expertise and attention to pests. Improper waste disposal |
| Climatic conditions | More moderate summer temperatures (Mediterranean) | High summer temperatures (above Mediterranean) |
| Cultivars | Varieties less susceptible; integrated nutrient management | Susceptible varieties, high fertiliser input |
Lower limit:
Low‐level persistent population that cannot be entirely eradicated.
Upper limit:
Poor agricultural practices that allow for re‐infestation from discarded infested material
Median:
Overall, well under control in an ideal scenario.
Inter‐quartile range:
High uncertainties below and above the median.
F.2. Impact with pest management practices in place
Table F.3 shows EKE estimates for yield loss with specific pest control measurements against L. orbonalis, informed by a meta‐analysis of losses (Appendix F, below) reported from control plots in pesticide trials and on the evidence summarised as bullet points below.
TABLE F.3.
Estimated mean reduction in yield (yield loss) caused by L. orbonalis with pest control in place.
| Question | What is the likely mean reduction of annual yield of eggplant, grown outdoors in a southern European country, with EI higher than 30 with specific pest control measures? | ||||||
|---|---|---|---|---|---|---|---|
| Results | Annual eggplant yield loss with specific control measures | ||||||
| Percentiles: % | 1% | 5% | 25% | 50% | 75% | 95% | 99% |
| EKE estimates | 0.1% | – | 0.3% | 0.5% | 1% | – | 3% |
| Fitted values (% yield loss) | 0.102% | 0.130% | 0.281% | 0.536% | 0.964% | 1.94% | 2.91% |
| Fitted distribution | BetaGeneral (1.0297, 163.33, 0,00095, 1) | ||||||
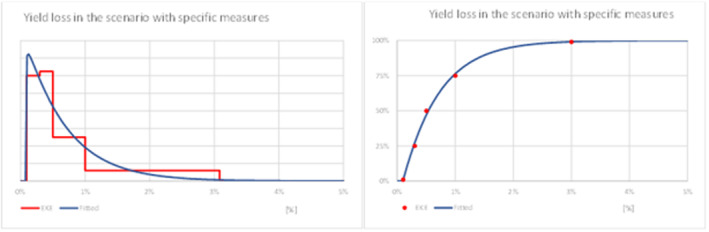
FIGURE F.2. Distribution of mean reduction of annual eggplant yield with specific control measures fitted to EKE estimates. (Left hand chart shows probability density function to describe the remaining uncertainties of the parameter; right hand chart shows cumulative distribution function (CDF) of the likelihood of the parameter).
The median annual yield loss in eggplant crops was estimated to be 0.536% (90% CR from 0.13% to 2.91%) in a scenario with specific pest management measurements is in place.
Reasoning
Dense pheromone trapping;
Public awareness campaigns.
TABLE F.4. Uncertainties associated with low and high yield loss scenarios.
| Uncertainty | Low yield loss scenario | High yield loss scenario |
|---|---|---|
| Agricultural practices | Good agricultural practices, e.g. crop rotation, sanitation, clean planting material, regular inspections by grower, pheromone trapping and mating disruption, spraying when insects are caught in traps | Amateur gardens; small scale; no pesticides. Staggered planting, some greenhouses, but not sanitated, so near‐Indian opportunities for field‐to‐field movement of the insect and perpetuation; lack of expertise and attention to pests; improper waste disposal; resistance to pesticides lowers efficacy of control |
| Climatic conditions | More moderate summer temperatures (Mediterranean) | High summer temperatures (above Mediterranean) |
| Cultivars | Resistant cultivars; integrated nutrient management | Susceptible varieties, high fertiliser input |
Lower limit:
Low‐level persistent population that cannot be entirely eradicated;
Effective mass trapping of males with dense network of pheromone traps;
Wide public awareness of effective integrated pest management.
Upper limit:
Poor agricultural practices that allow for re‐infestation from discarded infested material.
Poor public awareness of effective control measurements.
Median:
Overall, well under control in an ideal scenario.
Inter‐quartile range:
High uncertainties below and above the median.
F.3. Meta‐analysis of Leucinodes orbonalis infestation reported in Solanum melongena in the literature (with control)
The full texts of the publications on Leucinodes orbonalis, identified during the systematic search, were screened for information on L. orbonalis infestation. This meta‐analysis is based on the results of 17 independent experiments. Publications reported L. orbonalis shoot and fruit infestation in Solanum melongena. Data on infestation were extracted and analysed. The datafile consisted of 453 records (observations) (Table F.5 and Figures F.3 and F.4).
TABLE F.5.
Number of records used to analyse the impact of Leucinodes orbonalis infestation on eggplant (Solanum melongena).
| Number of records (N = 453) | |
|---|---|
| Fruit | 248 |
| Shoot | 178 |
| Not specified | 27 |
| Number of records (N = 453) | |
|---|---|
| India | 429 |
| Pakistan | 24 |
FIGURE F.3.
Percentage of Leucinodes orbonalis infestation on Solanum melongena when pesticides applied.
FIGURE F.4.
Leucinodes orbonalis infestation in India and Pakistan.
In the data set, we have data from India and Pakistan, so we are going to plot the data separately as well. Pakistan is a not suitable area for L. orbonalis and that could be a reason that would explain the low density in this country (Figure F.4).
The review on chemical pest (L. orbonalis) control in Asia on eggplants was completed and visualised in Table F.6.
TABLE F.6.
A–Z of insecticides (active compounds) tested in studies on the control of Leucinodes orbonalis on eggplant (Solanum melongena) in the places of origin.
| Active ingredient | ||||
|---|---|---|---|---|
| Acephate | Carbosulfan | Endosulfan | Malathion | Spinetoram |
| Acetamiprid | Cartap | Endrin | Monocrotophos | Spinosad |
| Avermectin | Chlorantraniliprole | Etofenprox | Neem | Sulphur |
| Azadirachtin | Chlorpyrifos | Fenpropathrin | Novaluron | Thiacloprid |
| Bacillus thuringiensis | Cypermethrin | Fenvalerate | Parathion | Thiamethoxam |
| Beauveria bassiana | Deltamethrin | Fipronil | Parathion‐methyl | Thiometon |
| Bifenthrin | Demeton‐S‐Methyl | Flubendiamide | Permethrin | Triazophos |
| Bt galleriae | Diazinon | Heptachlor | Phosalone | Trichlorfon |
| Bt kustaki | Dichlorvos | Imidacloprid | Phosphamidon | Urea |
| Carbaryl | Dimethoate | Indoxacarb | Profenofos | Zeta‐cypermethrin |
| Carbofuran | Emamectine | Lambda‐Cyhalothrin | Quinalphos | Zineb |
Using mixed models (lme in R with experiment as a random factor), the Panel estimated an overall percentage of infestation of 20.4 ± 3.1% on Solanum melongena when pesticides applied. Data were analysed by country. The Panel estimated an overall percentage of infestation in India of 21.5 ± 3.1% and in Pakistan of 1.9 ± 0.4%. In Pakistan, the infestation is reported in shoot.
Overall percentage of infestation in fruit was 26.0 ± 2.8% and shoot infestation was estimated as 7.3 ± 2.9%. There was one study that did not specified the organ in which they measured the infestation, the estimation of percentage of damage is 25.1 ± 11.2%, it might be in fruit.
In India, the percentage of infestation was 25.9 ± 2.9% in fruit, 7.7 ± 3.2% in shoot and 25.1 ± 11.5% in the not specified category.
F.4. Meta‐analysis of Leucinodes orbonalis infestation reported in Solanum melongena in the literature (without control)
The full texts of the publications on Leucinodes orbonalis, identified during the systematic search, were screened for information on L. orbonalis infestation. This meta‐analysis is based on the results of 24 independent experiments. Publications reported L. orbonalis shoot, fruit and seedlings infestation in Solanum melongena. Data on infestation were extracted and analysed. The datafile consisted of 287 records (observations) from India, Bangladesh, Sri Lanka and Pakistan (Table F.7 and Figures F.5, F.6).
TABLE F.7.
Number of records used to analyse the impact of Leucinodes orbonalis infestation on eggplant (Solanum melongena).
| Country | Number of records (N = 287) |
|---|---|
| India | 206 |
| Bangladesh | 18 |
| Sri Lanka | 5 |
| Pakistan | 4 |
| Number of records (N = 287) | |
|---|---|
| Fruit | 157 |
| Shoot | 84 |
| Seedlings | 39 |
FIGURE F.5.
Infestation of Solanum melongena (without control) and by country.
FIGURE F.6.
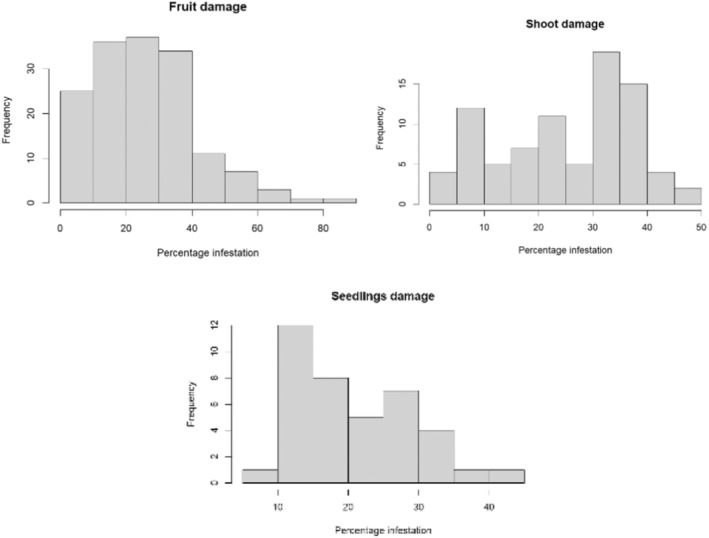
Infestation of Solanum melongena (without control) by plant organ.
Using mixed models (lme in R with experiment as a random factor), the Panel estimated an overall percentage of infestation of 29.7 ± 3.0% on Solanum melongena when no pesticides applied.
The estimated percentage of infestation in Sri Lanka was 47.6 ± 19.2%; in India, it was 31.4 ± 3.1%; in Bangladesh, it was 14.8 ± 0.6%; and in Pakistan, it was 4.7 ± 0.5%. The variable did not show significance in the models. We performed the arcsine Square root transformation and still not significance was shown.
There was a significant effect of the plant organ on the infestation. Higher infestation was estimated in fruit with a 31.0 ± 3.3%, followed by shoot infestation with a percentage of 29.7 ± 3.5%, and seedlings infestation of 20.1 ± 3.8%.
F.5. Analysis of Leucinodes orbonalis infestation reported in Solanum tuberosum in the literature (with control)
Impact on other solanaceous crops of economic importance in the EU (potato, tomato) was not assessed due to the paucity of available data (a single literature reference mentioning impact in tomato, and six references mentioning impact in potato), which indicates that L. orbonalis is not of major concern for potato or tomato cultivation. This is even more evident when comparing cultivation areas of crops in India: 0.53 million hectares for the highly affected eggplant (Indian Council of Agricultural Research, 2016) vs. 0.79 million hectares for tomato (Indian Council of Agricultural Research, undated) and 1.79 million hectares for potato (Scott & Suarez, 2011). Furthermore, neither potato tubers nor tomato fruit are among the commodities on which L. orbonalis was intercepted in the EU (Table C.1).
In potato, L. orbonalis larvae are shoot borers, and infestation rates are highest in the vegetative phase of potato plant growth. The larvae do not naturally feed on the tubers, although they can be reared on cut pieces of tubers under laboratory conditions. Potato is considered a suboptimal host, as L. orbonalis reared on potato tubers had a longer developmental period, a shorter adult life and a higher mortality as compared to specimens reared on eggplant fruit (Maureal et al., 1982). Cultivation conditions (sanitation) of potato in India (from where all reports on impact originate) are unknown, but they are supposedly not as well managed than in the EU.
Reported infestation rates on potato reach up to 40% under natural conditions, but the plants are apparently usually very rarely attacked, and even less so at such severe rates. Nair (1967) reports shoot infestation rates of 10%–40%, with up to four larvae feeding on the same plant. In a study on the efficacy of insecticides, Natikar and Balikai (2021) report a maximum shoot infestation by L. orbonalis of 21.73% for the untreated control group. Natikar and Balikai (2018a) report mean shoot infestation rates of 9.71%–28.86% in the plant's vegetative phase, 5.28%–22.33% in the reproductive phase and 0%–6.72% in the harvesting phase. Natikar and Balikai (2018b) report mean shoot infestation rates of 3.71%–17.12% in the plant's vegetative phase, 2.03%–10.4% in the reproductive phase, and 0%–2.52% in the harvesting phase. Hanapur and Nandihalli (2004) investigated the efficacy of insecticides and botanicals against L. orbonalis feeding on potato and observed a shoot infestation rate in the untreated control group of 40.4%–47.55%. These experimental settings unlikely reflect natural conditions (where natural enemies are present) and therefore likely overstate infestation rates. Furthermore, it was not clear from the two‐page article whether other pests were present in the experiment that may have impacted the yield loss even more than L. orbonalis. Murthy and Nandihalli (2003) report a yield loss of 14.77% of tubers.
As for impact of L. orbonalis on tomato, the only available information is the report by Das & Patnaik (1971), stating that the tomato winter crop of 1964 in Bhubaneswar (Odisha, India) had an infestation rate of 5–10%, with larvae feeding in both fruits and shoots.
The full texts of the publications on Leucinodes orbonalis, identified during the systematic search, were screened for information on levels of infestation when pesticides were used to control L. orbonalis. This analysis is based on the results of two independent experiments. Both publications reported on infestation of L. orbonalis in shoots of Solanum tuberosum (potato) in India. Data on shoot infestation were extracted and analysed. The datafile consisted of 36 records (Table F.8 and Figures F.7, F.8).
TABLE F.8.
Number of records used to analyse the impact of Leucinodes orbonalis infestation on Solanum tuberosum.
FIGURE F.7.
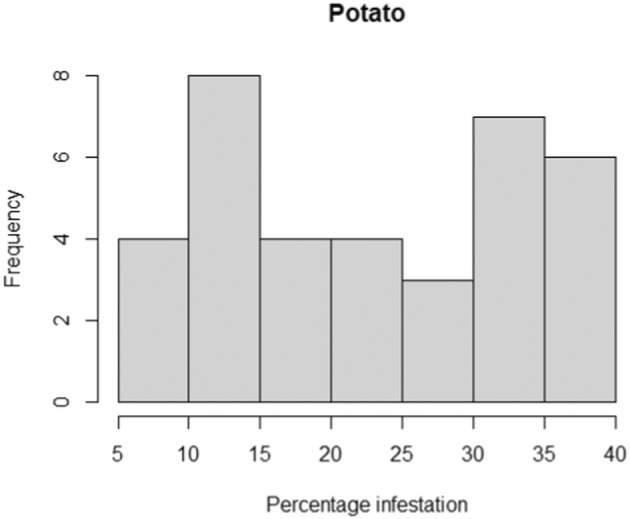
Percentage of Leucinodes orbonalis shoot infestation on Solanum tuberosum in India when pesticides applied.
FIGURE F.8.
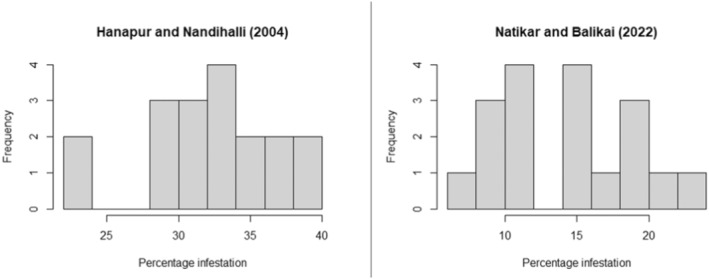
Percentage of Leucinodes orbonalis shoot infestation on Solanum tuberosum in India when pesticides applied.
The review on chemical pest (L. orbonalis) control in Asia on potato was completed and visualised in Table. F.9.
TABLE F.9.
Insecticides (active compounds) tested in studies on the control of Leucinodes orbonalis on potato (Solanum tuberosum) in the places of origin.
| Insecticides | No of records |
|---|---|
| Bifenthrin | 1 |
| Chlorfenapyr | 1 |
| Chlorpyrifos | 2 |
| Cyantraniliprole | 1 |
| Deltamethrin | 1 |
| Diafenthiuron | 1 |
| Emamectine | 1 |
| Endosulfan | 1 |
| Lambda‐Cyhalothrin | 1 |
| Profenofos | 1 |
| Spinosad | 1 |
| Spiromesifen | 1 |
| Thiodicarb | 2 |
| Botanicals | |
| Neem Gold | 1 |
| Neem Seed Kernal Extract (NSKE) | 1 |
| Vitex negundo (Lamiaceae) | 1 |
The reported percentage of shoot infestation in potato ranged from 5% to 40%.
Using a linear regression model (lm in R, with experiment as fixed factor), the Panel estimated an overall average percentage of shoot infestation in potato of 32.7 ± 1.0% in experiment 1 and 14.1 ± 1.0% in experiment 2.
F.6. Meta‐analysis of Leucinodes orbonalis infestation reported in Solanum spp. in the literature (without control)
The full texts of the publications on Leucinodes orbonalis, identified during the systematic search, were screened for information on L. orbonalis infestation. This meta‐analysis is based on the results of five independent experiments, originating from four publications. Publications reported L. orbonalis shoot, fruit and seedlings infestation in Solanum spp. Data on infestation were extracted and analysed. The datafile consisted of 159 records (observations) (Table F.7 and Figure F.9).
FIGURE F.9.
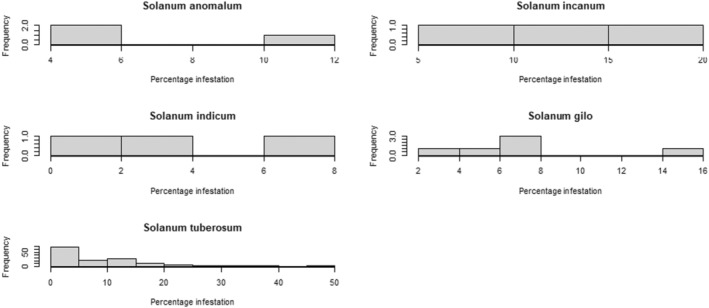
Percentage of Leucinodes orbonalis infestation on Solanum spp. when no control measures were applied.
TABLE F.10. Number of records used to analyse the impact of Leucinodes orbonalis infestation on Solanum spp.
| Host | Country | Number of records (N = 159) |
|---|---|---|
| Solanum tuberosum (potato) | India | 144 |
| Solanum anomalum (sodom apple) | India | 3 |
| Solanum incanum (bitter tomato or Sodom apple) | India | 3 |
| Solanum aethiopicum (bitter tomato) | India | 6 |
| Solanum indicum (African eggplant or Brihati) | India | 3 |
| Experiment | Solanum anomalum | Solanum aethiopicum (as S. gilo) | Solanum incanum | Solanum indicum | Solanum tuberosum |
|---|---|---|---|---|---|
| 1 – Hanapur and Nandihalli (2004) | 0 | 0 | 0 | 0 | 12 |
| 2 – Natikar and Balikai (2018a) | 0 | 0 | 0 | 0 | 120 |
| 3 – Natikar and Balikai (2022) | 0 | 0 | 0 | 0 | 12 |
| 4 – Behera and Gyanendra (2002) | 0 | 3 | 3 | 3 | 0 |
| 5 – Tejavathu et al. (1991) | 3 | 3 | 0 | 0 | 0 |
| Experiment 1–Hanapur and Nandihalli (2004) | Solanum anomalum | Solanum aethiopicum (as S. gilo) | Solanum incanum | Solanum indicum | Solanum tuberosum |
|---|---|---|---|---|---|
| Fruits | 0 | 0 | 0 | 0 | 0 |
| Seedlings | 0 | 0 | 0 | 0 | 0 |
| Shoots | 0 | 0 | 0 | 0 | 12 |
| Experiment 2–Natikar and Balikai (2018b) | Solanum anomalum | Solanum aethiopicum (as S. gilo) | Solanum incanum | Solanum indicum | Solanum tuberosum |
|---|---|---|---|---|---|
| Fruits | 0 | 0 | 0 | 0 | 0 |
| Seedlings | 0 | 0 | 0 | 0 | 0 |
| Shoots | 0 | 0 | 0 | 0 | 120 |
| Experiment 3 – Natikar and Balikai (2022) | Solanum anomalum | Solanum aethiopicum (as S. gilo) | Solanum incanum | Solanum indicum | Solanum tuberosum |
|---|---|---|---|---|---|
| Fruits | 0 | 0 | 0 | 0 | 0 |
| Seedlings | 0 | 0 | 0 | 0 | 0 |
| Shoots | 0 | 0 | 0 | 0 | 12 |
| Experiment 4 – Behera and Gyanendra (2002) | Solanum anomalum | Solanum aethiopicum (as S. gilo) | Solanum incanum | Solanum indicum | Solanum tuberosum |
|---|---|---|---|---|---|
| Fruits | 0 | 1 | 1 | 1 | 0 |
| Seedlings | 0 | 1 | 1 | 1 | 0 |
| Shoots | 0 | 1 | 1 | 1 | 0 |
| Experiment 5 – Tejavathu et al. (1991) | Solanum anomalum | Solanum aethiopicum (as S. gilo) | Solanum incanum | Solanum indicum | Solanum tuberosum |
|---|---|---|---|---|---|
| Fruits | 1 | 1 | 0 | 0 | 0 |
| Seedlings | 1 | 1 | 0 | 0 | 0 |
| Shoots | 1 | 1 | 0 | 0 | 0 |
Using mixed models (lme in R with experiment as a random factor), the Panel estimated an overall percentage of infestation of 13.8 ± 4.9% across all five species, including potato. There was no significant difference in infestation between the five Solanum species analysed (S. anomalum, S. indicum, S. incanum, S. aethiopicum (= S. gilo) and S. tuberosum) or between seedlings, stems, and fruits. There was a big difference among the different experiments, possibly related to growing conditions and management, population level of L. orbonalis in the environment and time in the season that observations were made.
F.7. Pest control in eggplants in Europe
There are a range of pests that effect eggplants in Europe (Table F.11). Aphids and red spider mites (Tetranychus spp.) are common pests of eggplants grown under glass in temperate countries (CABI, datasheet, 2023, other sources).
TABLE F.11.
Invertebrate pests of eggplants with a cosmopolitan or European distribution.
| Pest name | Common name | Class/family | Damage caused | Reference |
|---|---|---|---|---|
| Bemisia tabaci | Tobacco whitefly | Insecta/Aleyrodidae | Infests foliage, sucks sap, virus vector | Major pest, Hill (1987), Pollini (2021) |
| Trialeurodes vaporariorum | Whitefly | Insecta/Aleyrodidae | Infests foliage, sucks sap | CABI (2010) |
| Phthorimaea operculella | Potato tuber moth | Insecta/Gelechiidae | Larvae bore stems and mine leaves | Major pest, Hill (1987) |
| Tuta absoluta | Tomato leaf miner | Insecta/Gelechiidae | Leaves | MAPAMA (2021) |
| Acherontia atropos | African death's‐head hawkmoth | Insecta/Sphingidae | Larvae on the leaves | Pollini (2021) |
| Helicoverpa armigera | Corn earworm | (Insecta/Noctuidae) | Leaves | Pollini (2021) |
| Autographa gamma | Silver Y | (Insecta/Noctuidae) | Leaves | BOJA (2015) |
| Chrysodeixis chalcites | Golden twin‐spot moth | (Insecta/Noctuidae) | Leaves | BOJA (2015) |
| Heliothis peltigera | Bordered straw | (Insecta/Noctuidae) | Leaves | BOJA (2015) |
| Trichoplusia ni | Cabbage looper | (Insecta/Noctuidae) | Leaves | BOJA (2015) |
| Ostrinia nubilalis | European corn borer | Insecta/Crambidae | Branches | Pollini (2021) |
| Meloidogyne spp. | Root knot nematodes | Chromadorea/Meloidogynidae | CABI (2010) | |
| Tetranychus spp. | Spider mites | Arachnida/Tetranychidae | Feeding causes leaf chlorosis | Eggplant is major host (EPPO GD), Pollini (2021) |
| Nezara viridula | Green vegetable bug | Insecta/Pentatomidae | EPPO (2004), CABI (2010) | |
| Manduca quinquemaculata | Tomato hornworm | CABI (2010) | ||
| Aphids (e.g. Rhopalosiphonus latysiphon, Myzus persicaceae, Macrosiphum euphorbiae, Aphis gossypii, Aphis craccivora | Aphids (e.g. Bulb‐and potato aphid, peach‐potato aphid, potato aphid, cotton aphid, cowpea aphid) | Insecta/Aphididae | Infests foliage, sucks sap, virus vectors | EPPO (2004), Pollini (2021), MAPAMA (2021) |
| Leptinotarsa decemlineata | Colorado potato beetle | Insecta/Chrysomelidae | Larvae on the leaves | Pollini (2021) |
| Liriomyza huidobrensis | Pea leaf miner | Insecta/Agromyzidae | Larva on leaves (mining) | Pollini (2021) |
| Thrips (e.g. Frankliniella occidentalis, Thrips tabaci, Thrips angusticeps) | Western flower thrips, potato thrips, cabbage thrips | Insecta/Tripidae | Leaves, fruits | Pollini (2021), Phytoma (2022) |
| Lygus rugulipennis | European tarnished plant bug | Insecta/Myridae | Loss of flowers | Pollini (2021) |
| Erythroneura fasciaticollis | Insecta/Cicadellidae | Young plants, leaves | Pollini (2021) | |
| Empoasca flavescens (= vitis) | A leafhopper | Insecta/Cicadellidae | Sap sucking, causes foliage curling, stunting and leaf yellowing | EPPO (2004), CABI (2010) |
| Deroceras reticulatum | Grey field slug | Gastropoda/Agriolimacidae | Fruits | Pollini (2021) |
F.8. Pest control in solanaceous crops
Examining a range of gross margin budgets for eggplant and related solanaceous or similar crops indicates that from 0.8% to 4.9% of sales value can be expected to be spent managing a range of invertebrate pests (Table F.12). Rational growers would not spend more money on pest control than the minimum amount of damage that would be expected to be lost if pest management was not applied. Hence, the gross margin figures are assumed to represent the minimum losses that may be expected in western and more developed countries. Adding L. orbonalis to the pest fauna would add to the potential losses.
TABLE F.12.
Invertebrate pest management in solanaceous crops (excluding potatoes) as % of sales.
| Crop | Invertebrate pest management as % of sales value | Where | Principle invertebrate pests | Reference |
|---|---|---|---|---|
| Eggplant | 4.9 | Turkey | Aphids, spider mites, thrips, whitefly | Gül et al. (2022) |
| Capsicum | 2.4 | England | Spider mites, thrips, whitefly | Defra, unpublished (b) |
| Tomatoes | 1.8 | England | Lepidoptera, mealybugs, mites | Defra, unpublished (c) |
| Capsicum | 1.4 | Australia (New South Wales) | Aphids, Lepidoptera, thrips | Kelly et al. (2013) |
| Tomatoes | 0.8 | Australia (New South Wales) | Lepidoptera | NSW Government (2009) |
Oerke (2006) estimates annual yield losses in European potatoes due to animal pests (insects, mites, nematodes, slugs, snails, rodents, birds and mammals) to be approximately 7% (Table F.13). The principal insect pests of potatoes in Europe are listed in Table F.14.
TABLE F.13.
Estimates of % loss in yield in potatoes by animal pests around the world and at regional (approximately continental) scale (Oerke, 2006).
| Pest type: | Animal pests 1 | |||
|---|---|---|---|---|
| Losses | Potential losses 2 | Actual losses 3 | ||
| Scale | Worldwide | Regional | Worldwide | Regional 4 |
| Potatoes | 15.3 | 14–20 | 10.9 | 7–13 |
Animal pests are insects, mites, nematodes, slugs & snails, rodents, birds, mammals.
Potential losses are estimates of losses without pest management practices being applied.
Actual losses are yield losses despite pest management practices being applied.
Of all the world regions, losses in western Europe are among the lower losses; hence, actual losses in Europe from animal pests for potatoes is estimated to be 7%.
TABLE F.14.
Example invertebrate pests of potatoes with a cosmopolitan or European distribution.
| Pest name | Common name | Class/family | Damage caused | Reference |
|---|---|---|---|---|
| Myzus persicae | Peach‐potato aphid | Insecta/Aphididae | Infests foliage, sucks sap, virus vector | Major pest, Hill (1987) |
| Leptinotarsa decemlineata | Colorado potato beetle | Insecta/Chrysomelidae | Adults and larvae defoliate | Major pest, Hill (1987) |
| Eupteryx aurata | Potato leafhopper | Insects/Cicadellidae | Infests foliage, sucks sap, virus vector | Minor pest, Hill (1987) |
| Aphis nasturtii | Buckthorn potato aphid | Insecta/Aphididae | Infests foliage, sucks sap, virus vector | Minor pest, Hill (1987) |
| Macrosiphum euphorbiae | Potato aphid | Insecta/Aphididae | Infests foliage, sucks sap, virus vector | Minor pest, Hill (1987) |
| Heliothis spp. | Leafworms | Insecta/Noctuidae | Larvae eat leaves | Minor pest, Hill (1987) |
| Melolontha spp. | Cockchafers | Insecta/Scarabaeidae | Larvae in soil eat roots, holes tubers | Minor pest, Hill (1987) |
| Tipula spp. | Leatherjackets | Insecta/Tipulidae | Larvae is soil eat roots and damage tubers | Minor pest, Hill (1987) |
| Euxoa spp. | Cutworms | Insecta/Noctuidae | Larvae in soil eat roots, holes tubers | Minor pest, Hill (1987) |
| Agrotis spp. | Cutworms | Insecta/Noctuidae | Larvae in soil eat roots, holes tubers | Minor pest, Hill (1987) |
Pesticide use appears to have mixed success on the infestation of L. orbonalis in potato, since a meta‐analysis of data from the literature (Appendix F.5) finds infestation rates ranging from 5% to 40%. These rates are comparable to the results in another meta‐analysis (Appendix F.6) on the infestation of L. orbonalis without control measures in five different Solanum species, including S. tuberosum (Figure F.8). However, potato is rarely affected by L. orbonalis (six papers mentioning impact on potato vs. several hundred papers quantifying impact on eggplant), and pesticide use is therefore likely not very fine‐tuned to address L. orbonalis infestation. Furthermore, many of these infestation rate numbers originate from untreated control groups in pesticide studies, in which the control group likely experiences heavier infestation rates than expected under natural conditions that include natural enemies (parasitoids, predators).
TABLE F.15. Invertebrate pests of tomatoes with a cosmopolitan or European distribution.
| Pest name | Common name | Family | Damage caused | Reference |
|---|---|---|---|---|
| Bemisia tabaci | Tobacco whitefly | Aleyrodidae | Suck sap, infest foliage, transmit virus | Major pest (Hill, 1987) |
| Trialeurodes vaporariorum | Glasshouse whitefly | Aleyrodidae | Suck sap, infest foliage, transmit virus | Major pest (Hill, 1987) |
| Nezara viridula | Green stink bug | Pentatomidae | Suck sap, toxic saliva causes spots on fruit and leaves | Major pest (Hill, 1987) |
| Heliothis spp. | Tomato fruitworms | Noctuidae | Larvae bore into fruits | Major pest (Hill, 1987) |
| Thrips tabaci | Onion thrips | Thripidae | Infest foliage, suck cell contents | Major pest (Hill, 1987) |
| Tetranychus urticae | Red spider mite | Tetranychidae | Scarify (surface damage) | Major pest (Hill, 1987) |
| Empoasca spp. | Leafhoppers | Cicadellidae | Infest foliage, sucks sap | Minor pest (Hill, 1987) |
| Various aphid spp. | Aphids | Aphididae | Infest foliage, sucks sap | Minor pest (Hill, 1987) |
| Frankliniella spp. | Thrips | Thripidae | Infests flowers, foliage | Minor pest (Hill, 1987) |
APPENDIX G. Identification of pest management practices used in solanaceous crops against Leucinodes orbonalis in Asia
G.1.
Risk reduction options were assessed through a literature review. The main results of the literature review are summarised in the following table, and they regard mostly eggplants.
| Chemical | Biological | Biotechnological | Cultural practices | Monitoring |
|---|---|---|---|---|
|
Mass trapping (with sex pheromones) Use of botanical substances such as neem oil derivates, among other plant species extracts Pesticides (active substances): Abamectin (aka avermectin) Acephate Acetamiprid Alpha‐cypermethrin (aka alphamethrin) Azadirachtin Bacillus thuringiensis (subsp. kurstaki, galleriae) Beauveria bassiana Bifenthrin Carbaryl Carbofuran Carbosulfan Cartap Chlorantraniliprole Chlorfenapyr Chlorpyrifos Copper oxychloride Cyantraniliprole Cypermethrin DDT Deltamethrin Demeton‐S‐methyl Diafenthiuron Diazinon Dichlorvos Diflubenzuron Dimethoate Emamectin Endosulfan Endrin Etofenprox Fenpropathrin Fenvalerate Fipronil Flubendiamide Flufenoxuron Formothion Heptachlor Imidacloprid Indoxacarb lambda‐Cyhalothrin Lindane (aka BHC) Lufenuron Malathion |
Parasitoids: Trathala flavoorbitals (Hymenoptera:Ichneumonidae), Trichogramma chilonis and T. pretiosum (Hymenoptera:Trichogrammatidae), Eriborus argenteopilosus (Hymenoptera: Ichneumomidae), Phanerotoma sp. (Hymenoptera: Braconidae), Campyloneura sp. (Diptera:Cecidomyiidae), Pristomerus testaceus (Hymenoptera:Ichneumonidae), Pseudoperichaeta sp. (Diptera:Tachinidae), Priomerus testaceus (Hymenoptera:Ichneumonidae), Chelonus sp. (Hymenoptera: Braconidae), Bracon hebetor and B. brevicornis (Hymenoptera: Braconidae), Antrocephalus mitys (Hymenoptera:Chalcididae), Brachymeria lasus (Hymenoptera:Chalcididae), Spalangia irregularis and S. endius (Hymenoptera:Chalcididae), Apanteles sp. (Hymenoptera: Braconidae), Dermatopelte sp. (Hymenoptera:Eulophidae), Cremastus hapaliae (Hymenoptera: Ichneumomidae), Xanthopimpla punctate (Hymenoptera: Ichneumomidae), Itamoplex sp. (Hymenoptera: Ichneumomidae), Diadegma apostate (Hymenoptera: Ichneumomidae) Predators: Chrysoperla carnea, C. kulingensis (Neuroptera), Orius niger (Hemiptera), Coccinella septempunctata (Coleoptera), Tetraponera sp. (Hymenoptera), Campyloneura sp. (Hemiptera), Cheilomenes sexmaculata (Coleoptera), Brumoides suturalis (Coleoptera) Entomopathogenic nematodes: Heterorhabditis indica, Steinernema carpocapsae, S. bicornutum |
GMO crops to produce resistant varieties: Cry1Ac, Cry1Ab, Cry1Aa3 and Cry2Ab protein in eggplant; Bt eggplant |
Resistant varieties (not genetically modified) Intercropping Proper cutting, removal and destroying of infested plant parts of eggplant (especially up to the first harvest) The harvest of only healthy fruits Timely cleaning of plant residues of host plants at the end of the growing season. Regular pruning Wider spacing between rows (40+ cm) Bagging the fruit during the growing phase to prevent borer attack Vermicompost Low soil moisture |
Sex pheromone traps (gland extracts of the female brinjal fruit and shoot borer; synthetically produced pheromone blend) Traps: Sticky delta trap, sticky wing trap, Uni‐trap, Spodoptera trap, Open‐delta trap, Water trap, Funnel trap, Water trap, Flight‐T trap, Cross winged trap |
|
Methomyl Monocrotophos Novaluron Oxymatrine Parathion Parathion‐methyl Phenthoate Phorate Phosalone Phosphamidon Profenofos Pyriproxyfen Quinalphos Spinetoram Spinosad Spiromesifen Sulphur Teflubenzuron Thiacloprid Thiamethoxam Thiodicarb Thiometon Triazophos Trichlorfon Triflumuron Urea zeta‐Cypermethrin Zineb Beta‐Cyfluthrin |
Entomopathogenic fungus: Trichoderma longibrachiatum, Aspergillus ochraceus, A. niger, Beauveria bassiana, Penicillium expansum, Bipolaris tetramera Entomopathogenic bacteria: Serratia marcescens |
|||
| References | ||||
| Abrol and Singh (2003), Adhi et al. (2022), Adusei and Azupio (2022), Affrin et al. (2017), Ahmad et al. (2018), Ajabe et al. (2019), Akhtar et al., 2019; Ali et al. (2016), Ali and Khan (2022), Ansari et al. (2019), Ashadul et al. (2014), Attygalle et al. (1988), Bade et al. (2017), Bakhat et al. (2018), Baskaran and Kumar (1980), Behera and Gyanendra (2002), Bhandari et al. (2015), Bharati et al. (2015), Biswas et al. (2009), Challa et al. (2021), Chandi and Kaur (2021), Chandi et al. (2022), Chandre Gowda (1980), Chatterjee and Roy (2004), Chi et al. (2021), Chowdhury et al. (2017), Čokl and Millar (2009), Cork et al. (2005), Dash and Singh (1990), David (1966), Dehariya et al. (2017), Deore and Patil (1996), Deotale et al. (2000), Dhamdhere and Sharma (1991), Ebert and Wu (2011), Fasihur et al. (2017), Gahukar and Balpande (1997), Gayathri and Geetha (2020), Ghatak et al. (2009), Gholami and Sadeghi (2016), Ghosal et al. (2013), Ghosh and Senapati (2009), Gowda et al. (1977), Gowrish et al. (2015), Hanapur and Nandihalli (2004), Jayakrishnan and Madhuban (2012), Jaydeep et al. (2022), Joshi and Sharma (1973), Kadam et al. (2006), Kariyanna et al. (2020a), Kariyanna et al., 2020b Kaur (2004), Kaur et al. (2022), Kaur et al. (2014), Kodandaram et al. (2015), Krishnaiah et al. (1981), Krishnamoorthy et al. (2014), Kumar and Shukla (2002), Kumar et al. (2000), Lall and Ahmad (1965), Latif et al. (2010), Latif et al. (2009), Mahata et al. (2014), Mahesh and Men (2007), Mahla et al. (2017), Mandal et al. (2011), Mane and Kulkarni (2010), Manickam et al. (2014), Manoj and Ravi (2019a), Manoj and Ravi (2019b), Marimuthu (2008), Ullah et al. (2022), Mehta et al. (1998), Mishra and Mishra (1995), Mohan and Prasad (1986), Munje et al. (2015), Murali et al. (2017), Murugesan and Murugesh (2009), Muthukumar and Kalyansundaram (2003), Nahar et al. (2020), Naik et al. (2008), Nair and Nair (1977), Narendra et al. (2015), Nathani (1983), Natikar and Balikai (2018a), Natikar and Balikai (2022), Nawale and Sonone (1977), Nusra et al. (2020), Pandey et al. (2021), Pareet and Basavanagoud (2009), Parvatha (2014), Patial et al. (2009), Patnaik et al. (2004), Pawar et al. (1987), Pawar et al. (2016), Pawar et al. (2020), Priya and Ravi (2020), Prodhan et al. (2019), Puranik et al. (2002), Quamruzzaman et al. (2019), Qureshi et al. (1998), Rabindra and Janardan (2012), Radhakrishore et al. (2014), Rahman et al. (2019), Rajavel et al. (1989), Rajendra et al. (2015), Ramawtar et al. (2015), Rani et al. (2017), Rao and Ramana (2017), Khatun et al. (2022), Rohokale et al. (2018), Roy et al. (2016), Roy and Panda (1973), Roy et al. (1973), Sahoo et al. (2013), Samanta et al. (1999), Sandanayake (1992), Sandip et al. (2018), Sandip et al. (2016), Sanjeevi and Muthukrishnan (2017), Sarangdevot et al. (2010), Sardar and Guram (1967), Sarkar et al. (2022), Sarnabati and Ray (2017), Satpathy et al. (2006), Satpute et al. (2002), Singh (2019), Sharma et al. (2004), Sharmin et al. (2017), Shelton (2021), Singh (2018b), Singh et al. (2009), Singh and Kumar (2011), Singh et al. (2015a), Singh et al. (2015b), Singh et al. (2021), Singh and Dwivedi (2020), Singh and Sinha (1964), Sinha et al. (1976), Sinha and Gopal (2002), Sinha and Nath (2011), Sinha and Vishwa (2012), Sivakumar et al. (2003), Sugandha et al. (2017), Tejavathu et al. (1991), Tripathi (2016), Tripathy and Patnaik (2003), Veeravel and Baskaram (1977), Venkatraman and Chander (1967), Vijayasree and Bai (2019), Vinothkumar (2021), Visnupriya and Muthukrishnan (2019), Vlnothkumar (2021), Walunj et al. (1996), Wankhede and Kale (2010a), Wankhede and Kale (2010b), Witzgall et al. (2010), Yadav et al. (2015), Yazdani et al. (1981), Yengkokpam and Mazumder (2021), Yousafi et al. (2015), Yousafi et al. (2016) | Abdullah et al. (2021), Al‐Ani and Mohammed (2020), Chowdhury et al. (2017), Cork et al. (2001), Fathi (2022), Ferreira and Musumeci (2021), Funde et al. (2020), Ghosh et al. (2016), Gulati et al. (2021a), Gulati et al. (2021b) Heinrichs and Muniappan (2016), Hussaini (2014), Hussaini et al. (2002), Jalali (2013), Kaur et al. (2019), Lalitha et al. (2013), Li et al. (2014), Mallik et al. (1989), Manoj and Ravi (2019a), Manoj and Ravi (2019b) Mejía et al. (2021), Mukherjee and Ghosh (2023), Nair (1967), Niranjana et al. (2015), Niranjana and Sridhar (2014), Parvatha (2014), Patial and Mehta (2006), Podder and Ghosh (2019), Poveda (2021), Prasad and Singh (2012), Prasad et al. (2012) Ranjith et al. (2020), Sankaranarayanan and Askary (2017), Singh and Singh (2002), Sivapragasam and Chua (1997), Srinivasan (2008), Talukder et al. (2021), Venkatesan et al. (2009), Verma and Lal (1985), Visalakshy et al. (2009), Wu et al. (2013), Yasodha and Natarajan (2009), Yasodha et al. (2007) | Akshat and Vijay (2007), Alam and Salimullah (2021), Anderson et al. (2019), Baksh and Iqbal (1979), Collonnier et al. (2001), Dattatray et al. (2018), Gowda et al. (1990), Haque and Saha (2020), Hasan et al. (2021), Hautea et al. (2016), Kalia et al. (2013), Kariyanna et al. (2020a), Kariyanna et al. (2020b), Kashyap et al. (2003), Khan and Borikar (2017), Kudlu and Stone (2013), Kumar et al. (1998), Kumar et al. (2011), Lobato‐Gómez et al. (2021), Mally et al. (2015), Narendran et al. (2006), Navasero et al. (2016), Pal et al. (2009), Pray et al. (2011), Prodhan et al. (2019), Prodhan et al. (2019), Rai et al. (2013), Ranjithkumar et al. (2013), Saini and Kaushik (2019), Sharma et al. (2020), Shelton (2010), Shelton et al. (2018), Shelton (2021), Shelton et al. (2019), Shende et al. (2016), Shrivastava et al. (2011), Shukla et al. (2017), Silva Dias and Ryder (2011), Sofiya and Raj (2021), Sultana et al. (2018), Taylo et al. (2016), Thomson (2018), Xiao and Wu (2019) | Ahmad et al. (2018), Akulov et al. (2015), Amin et al. (2019), Baidoo et al. (2018), Chakraborti and Priyadarshini (2021), Chowdhury et al. (2017), Collonnier et al. (2001), Das et al. (2014), Degri (2014), Dehariya et al. (2017), Dhankar (1988), Dhankhar et al. (1979), Ebert (2017), Fathi (2022), Ghorai (2005), Hanur et al. (2011), Kashyap et al. (2003), Kaur et al. (2004), Keatinge et al. (2011), Khorsheduzzaman et al. (2010), Krithika and Ananthanarayana (2017), Kumar et al. (2012), Mainali et al. (2014), Malshe et al. (2016), Muhammad et al. (2021), Nahar (2020), Naik et al. (2017), Najar et al. (2015), Navasero and Calumpang (2013), Nawale et al. (1977), Parvatha (2014), Puthiamadom and Joseph (2022), Raeyat et al. (2021), Raja et al. (1998), Sahu et al. (2017), Sardar and Guram (1967), Satpathy and Mishra (2011), Sharma et al. (2020), Sharmin et al. (2017), Shelton et al. (2019), Shirale et al. (2017), Sidhu and Dhatt (2007), Singh (2018a), Singh (2018b), Sohi (1944), Sorensen et al. (2016), Srinivasan and Huang (2009), Sujayanand et al. (2015), Suresh et al. (2007) | Akulov et al. (2015), Al‐Amin et al. (2022), Cork et al. (2003), Cork et al. (2005), Cork and Hall (1998), Goundalkar and Webster (2007) Gunawardena et al. (1989), Li et al. (2014), Manasa and Gangadhar (2018), Manasa et al. (2016), Manoj et al. (2018), Mazomenos et al. (1994), Mazumder and Khalequzzaman (2010), Nayak (2014), Nayak et al. (2012), Prabakaran et al. (2019), Prasannakumar et al. (2011), Sharmin et al. (2017), Van Vang et al. (2018), Ye et al. (2015) |
APPENDIX H. Area and production of eggplant cultivation in the EU, 2010–2022
H.1.
Appendix H details the area of EU eggplant production (Tables H.1 and H.2).
TABLE H.1.
Eggplant production area (cultivation/harvested/production) (ha).
| EU MS | 2010 | 2011 | 2012 | 2013 | 2014 | 2015 | 2016 | 2017 | 2018 | 2019 | 2020 | 2021 | 2022 | Mean annual area (ha) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| IT | 10,740 | 9420 | 8300 | 8430 | 10,330 | 10,150 | 10,030 | 9450 | 9560 | 9550 | 9510 | 9570 | 9600 | 9588 |
| RO | 5800 | 5390 | 4910 | 4730 | 4890 | 4880 | 4560 | 4420 | 4800 | 4810 | 4410 | 4940 | 4100 | 4818 |
| ES | 3440 | 3670 | 3890 | 3670 | 3410 | 3840 | 3750 | 3580 | 3620 | 3470 | 3700 | 3590 | 3650 | 3637 |
| EL | 2600 | 2500 | 2230 | 2230 | 2300 | 1880 | 1750 | 1700 | 1670 | 1350 | 1500 | 1390 | 1540 | 1895 |
| FR | 620 | 720 | 470 | 700 | 710 | 710 | 730 | 730 | 800 | 710 | 810 | 1420 | 1410 | 811 |
| BG | 0 | 0 | 0 | 300 | 310 | 490 | 310 | 480 | 440 | 390 | 370 | 320 | 420 | 295 |
| NL | 100 | 100 | 110 | 100 | 100 | 110 | 110 | 100 | 110 | 120 | 130 | 120 | 130 | 111 |
| PT | – | 100 | 90 | 90 | 100 | 60 | 100 | 140 | 110 | 80 | 90 | 160 | 90 | 101 |
| HU | 40 | 50 | 70 | 40 | 40 | 90 | 50 | 50 | 50 | 40 | 40 | 50 | 30 | 49 |
| CY | 20 | 20 | 20 | 30 | 40 | 30 | 40 | 30 | 30 | 20 | 30 | 30 | 30 | 28 |
| BE | 20 | 20 | 20 | 20 | 20 | 20 | 20 | 20 | 20 | 20 | 30 | 30 | 30 | 22 |
| PL | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 200 | 0 | 15 |
| SI | 0 | 0 | 0 | 0 | 0 | 20 | 20 | 20 | 20 | 30 | 30 | 20 | 20 | 14 |
| AT | 10 | 10 | 10 | 10 | 10 | 10 | 10 | 10 | 10 | 10 | 10 | 10 | 10 | 10 |
| SK | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 10 | 10 | 0 | 10 | 0 | 0 | 3 |
| CZ | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| DK | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| DE | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| EE | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| IE | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| HR | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| LV | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| LT | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| LU | – | – | – | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| MT | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| FI | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| SE | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Sum | 23,390 | 22,000 | 20,120 | 20,350 | 22,260 | 22,290 | 21,480 | 20,740 | 21,250 | 20,600 | 20,670 | 21,850 | 21,060 | 21,397 |
Source: Eurostat [APRO_CPSH1__custom_6146643] (Code V3410); Countries ranked by annual mean area (ha).
TABLE H.2.
Production of EU eggplant 2010–2022 (tonnes) by EU MS 2010–2022; countries ranked by mean annual production (tonnes)
| 2010 | 2011 | 2012 | 2013 | 2014 | 2015 | 2016 | 2017 | 2018 | 2019 | 2020 | 2021 | 2022 | Mean annual production (tonnes) | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| IT | 302,550 | 243,200 | 217,690 | 225,930 | 308,720 | 300,180 | 317,590 | 286,470 | 298,310 | 300,620 | 304,690 | 306,440 | 307,430 | 286,140 |
| ES | 190,200 | 215,770 | 245,920 | 206,330 | 208,210 | 244,540 | 242,640 | 225,910 | 238,330 | 245,150 | 282,200 | 265,290 | 276,320 | 237,447 |
| RO | 73,620 | 86,390 | 70,310 | 74,810 | 80,870 | 80,670 | 67,680 | 78,460 | 88,150 | 79,660 | 88,420 | 95,860 | 63,250 | 79,088 |
| EL | 70,910 | 78,010 | 73,020 | 72,730 | 67,010 | 69,140 | 70,140 | 63,190 | 70,940 | 54,910 | 60,980 | 59,770 | 58,060 | 66,832 |
| NL | 46,000 | 46,000 | 47,000 | 48,000 | 51,000 | 53,000 | 54,000 | 53,000 | 55,000 | 62,000 | 65,000 | 63,000 | 66,000 | 54,538 |
| FR | 19,810 | 19,260 | 17,790 | 26,420 | 29,780 | 30,550 | 31,770 | 30,080 | 31,990 | 29,570 | 28,830 | 59,170 | 42,480 | 30,577 |
| BG | 8000 | 8800 | 6970 | 6150 | 7650 | 8100 | 8100 | 8100 | 10,120 | 10,580 | 11,400 | 14,160 | 13,820 | 11,086 |
| BE | 0 | 0 | 7800 | 8500 | 9400 | 9930 | 7910 | 11,910 | 11,260 | 11,180 | 9920 | 7800 | 9690 | 8100 |
| PT | – | 4900 | 4900 | 6010 | 6290 | 2970 | 6050 | 8150 | 6850 | 5660 | 6500 | 10,160 | 5390 | 6153 |
| CY | 2510 | 2600 | 2380 | 2440 | 2750 | 2140 | 2520 | 2190 | 1710 | 1230 | 1340 | 1290 | 1300 | 2031 |
| HU | 2150 | 2270 | 1450 | 1270 | 1290 | 2210 | 900 | 830 | 990 | 800 | 830 | 1060 | 640 | 1284 |
| PL | 570 | 410 | 350 | 680 | 890 | 1070 | 980 | 1060 | 1130 | 1170 | 1420 | 1810 | 1770 | 1024 |
| AT | 760 | 750 | 700 | 750 | 790 | 740 | 810 | 790 | 840 | 860 | 850 | 730 | 820 | 784 |
| MT | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 500 | 2400 | 900 | 292 |
| SI | 0 | 0 | 0 | 0 | 0 | 330 | 320 | 330 | 330 | 500 | 550 | 420 | 340 | 390 |
| SK | 0 | 0 | 0 | – | 40 | 90 | 90 | 130 | 210 | 0 | 110 | 0 | 0 | 112 |
| SE | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 20 | 20 | 20 | 20 | |
| Sum | 717,080 | 708,360 | 696,280 | 680,020 | 774,690 | 805,660 | 811,500 | 770,600 | 816,160 | 803,890 | 863,560 | 889,380 | 848,230 | 783,493 |
Note: Data for CZ, DK, DE, EE, IE, HR, LV, LT, LU and FI were zero in all years and therefore omitted. Data for LU were missing in 2011 and 2012.
APPENDIX I. Mathematical description of the pathway model for assessing entry and establishment of new founder populations in the EU territory
I.1.
A pathway model is mathematically the product (multiplication) of a set of random variables (Douma et al., 2016; van der Gaag et al., 2019). Most of the variables are uncertain having a probability distribution, which is either based directly on data or on expert knowledge elicitation. The model also contains some constants.
The model is:
, where the meaning of the symbols is given in the table below.
| Variable | Meaning | Units of measurement | Excel name a | |
|---|---|---|---|---|
|
|
Outcome variable of the pathway model: number of founder populations per year across whole of the EU | Number per year | ab | |
|
|
Import of eggplant + turkey berry from Asian countries in which L. orbonalis is present | t (1000 kg) per year | b | |
|
|
Proportion of the European population living in NUTS regions with an EI equal to or larger than 30 or 15 (depending on the scenario) | – | c | |
|
|
Conversion factor from ton to gram | g/t | ||
|
|
Proportion of turkey berry in the trade in eggplant + turkey berry and other Solanum spp. Collected under the CN code 07093000 | – | a | |
|
|
Weight of a cluster of turkey berries | g | h | |
|
|
Weight of a single eggplant fruit (aubergine) | g | g | |
|
|
Infestation rate: proportion of single fruit (eggplant) or fruit clusters (turkey berry) infested with L. orbonalis | per 10,000 | l | |
|
|
Conversion factor to convert proportion expressed as infested fruit per 10 000 to proportion of infested fruit | |||
|
|
Proportion waste pre‐consumer (commercial waste) | – | n | |
|
|
Proportion waste consumer | – | o | |
|
|
Proportion of adults emerging from commercial waste | r | ||
|
|
Proportion of adults emerging from consumer waste | s | ||
|
|
Proportion of females among adults | – | w | |
|
|
Proportion of females finding mating partner | x | ||
|
|
Likelihood that mated female finds host and founds a persistent population (e.g. survives predation and overcomes an initial Allee effect) | z |
Excel name refers to the name of the variable in the Excel implementation of the pathway model. See column E in tabs 3‐8 of the Excel implementation of the pathway model, which is available in the Supplementary Materials. Each of the tabs 3‐8 represents a scenario (combination of EI 15 or 30, and three compositions of the trade in eggplant + S. torvum).
Each of the variables was random, except (waste‐pre‐consumer) which was set at a value of 0.1 based on empirical data for Elasmopalpus lignosellus in asparagus spears (Gould & Maldonado, 2006; Gould et al., 2006).
The outcome variable represents the number of founder populations per year due to introduction of the organism in the EU with the trade in eggplant and turkeyberry. That is the frequency of founding events.
Calculations with the pathway model are made using Monte Carlo simulation. This is done by randomly drawing the values of the variables on the right hand side from their probability distributions, and calculating for each set of values of the corresponding value of and determining the distribution of across the Monte Carlo replicates.
The inverse of each value of , i.e. represents the expected waiting time until the next founding event.
The pathway model has several intermediate results that are interpretable.
For instance, is the import volume of eggplant and turkey berry into areas suitable for establishment of L. orbonalis in the EU during one year, measured in g/year.
is the total number of pathway units (eggplant + S. torvum fruit clusters) imported into the area suitable for establishment of L. orbonalis in the EU during one year.
is the total number of moths emerging from waste discarded in the suitable areas of the EU during one year.
Additional intermediate results are available. Further information is given in the Excel implementation of the pathway model that is included in the supplementary information of this opinion.
EFSA PLH Panel (EFSA Panel on Plant Health) , Bragard, C. , Baptista, P. , Chatzivassiliou, E. , Di Serio, F. , Gonthier, P. , Jaques Miret, J. A. , Justesen, A. F. , MacLeod, A. , Magnusson, C. S. , Milonas, P. , Navas‐Cortes, J. A. , Parnell, S. , Potting, R. , Reignault, P. L. , Stefani, E. , Thulke, H.‐H. , Civera, A. V. , Yuen, J. , Zappalà, L. … Van der Werf, W. (2024). Pest risk assessment of Leucinodes orbonalis for the European Union. EFSA Journal, 22(3), e8498. 10.2903/j.efsa.2024.8498
Adopted: 30 November 2023
REFERENCES
- Abdullah, N. S. , Doni, F. , Mispan, M. S. , Saiman, M. Z. , Yusuf, Y. M. , Oke, M. A. , & Mohd Suhaimi, N. S. (2021). Harnessing Trichoderma in agriculture for productivity and sustainability. Agronomy, 11. [Google Scholar]
- Abrol, D. P. , & Singh, J. B. (2003). Relative efficacy of some insecticides against Brinjal fruit and shoot borer, Leucinodes orbonalis Guen., and their impact on fruit yield. Journal of Asia‐Pacific Entomology, 6, 83–90. [Google Scholar]
- Adhi, S. , Reddy, T. P. , Reddy, M. J. M. , Afifa, J. , Rajashekhar, M. , Rajashekhar, B. , Ramakrishna, K. , & Bhatt, P. S. (2022). Impact of frontline demonstration on integrated management of Brinjal shoot and fruit borer (Leucinodes orbonalis Guenee) in Nagarkurnool district, Telangana state. International Journal of Bio‐Resource and Stress Management, 13, 292–298. [Google Scholar]
- Adusei, S. , & Azupio, S. (2022). Neem: A novel biocide for Pest and disease control of plants. Journal of Chemistry, 2022, 1–12. [Google Scholar]
- Affrin, K. , Reshma, P. , & Kumar, G. N. (2017). Control and management of L. orbonalis using WSNs. IEEE Technological Innovations in ICT for Agriculture and Rural Development (TIAR), 8–13. [Google Scholar]
- Ahmad, S. I. , Pathania, S. S. , Nehru, R. K. , & Rouf, W. A. (2018). Efficacy of management modules against brinjal shoot and fruit borer Leucinodes orbonalis in Kashmir. Indian Journal of Entomology, 80, 527–530. [Google Scholar]
- Ajabe, S. S. , Jayewar, N. E. , Gaikwad, A. D. , & Ambad, R. B. (2019). Monitoring of brinjal shoot and fruit borer Leucinodes orbonalis by using sex pheromone and the impact of abiotic factors on trap catches. Journal of Entomology and Zoology Studies, 7, 350–352. [Google Scholar]
- Akhtar, N. , Khan, M. A. , Bashir, H. , & Ahmad, A. (2019). Management of eggplant (Solanum melongena L.) shoot and fruit borer (Leucinodes orbonalis Guenee) by using different insecticides. Proceedings of Pakistan Congress of Zoology, 39, 182–183. [Google Scholar]
- Akshat, M. , & Vijay, V. (2007). Successful commercialization of insect‐resistant eggplant by a public‐private partnership: Reaching and benefiting resource‐poor farmers. pp. 1829–1831.
- Akulov, E. N. , Kirichenko, N. I. , & Takhtuev, A. S. (2015). Опасный вредитель пасленовых–огневка Leucinodes orbonalis–обнаружен в импортной продукции в России [Dangerous nightshade pest–Leucinodes orbonalis moth–found in imported products in Russia.]. Защита и карантин растений, 39–41. [Google Scholar]
- Alam, I. , & Salimullah, M. (2021). Genetic engineering of eggplant (Solanum melongena L.): Progress, controversy and potential. Horticulturae, 7. [Google Scholar]
- Alam, S. N. , Rashid, M. A. , Rouf, F. M. A. , Jhala, R. C. , Patel, J. R. , Satpathy, S. , Shivalingaswamy, T. M. , Rai, S. , Wahundeniya, I. , Cork, A. , Ammaranan, C. , & Talekar, N. S. (2003). Development of an integrated pest management strategy for eggplant fruit and shoot borer in South Asia. Shanhua, Taiwan: AVRDC—The world vegetable center. Technical bulletin No. 28. AVRDC publication No. 03‐548. 56 pp.
- Al‐Amin, M. , Hossain, M. , El, A. , Hasan, T. , Alim, F. , & Abdul, M. (2022). Performance of different designed pheromone traps and their placement heights in capturing Leucinodes orbonalis Guenee (Lepidoptera: Pyralidae) Munis. Entomology & Zoology, 17, 589–599. [Google Scholar]
- Al‐Ani LKT and Mohammed AM, 2020. Versatility of Trichoderma in plant disease management. 159–168.
- Ali, F. , Khan, I. , Zakirullah, A. A. , & Anees‐ur‐Rahman, A. (2016). Study on the population trend of brinjal fruit borer Leucinodes orbonalis Guen. (Lepidoptera: Pyralidae) and it's susceptibility to different insecticides ARPN. Journal of Agricultural and Biological Science, 11, 262–266. [Google Scholar]
- Ali, U. , & Khan, M. F. (2022). Population studies of brinjal fruit borer (shoot borer), Leucinodes orbonalis with reference to their chemical management measure. International Journal of Biology and Biotechnology, 19, 45–50. [Google Scholar]
- Amin, S. M. R. , Alam, M. Z. , Rahman, M. M. , Hossain, M. M. , & Mian, I. H. (2019). Evaluation of integrated approaches for the management of brinjal shoot and fruit borer utilizing antixenotic variety of brinjal. Munis Entomology and Zoology, 14, 591–601. [Google Scholar]
- Anderson, J. A. , Ellsworth, P. C. , Faria, J. C. , Head, G. P. , Owen, M. D. K. , Pilcher, C. D. , Shelton, A. M. , & Meissle, M. (2019). Genetically engineered crops: Importance of diversified integrated pest management for agricultural sustainability. Frontiers in Bioengineering and Biotechnology, 7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ansari MS, Basri R and Shekhawat SS, 2019. Insect pests infestation during field and storage of fruits and vegetables. 121–207.
- Ardez, K. P. , Sumalde, A. C. , & Taylo, L. D. (2008). Ovipositional preference, host range and life history of eggplant fruit and shoot borer, Leucinodes orbonalis Guenee (Lepidoptera: Pyralidae). Philippine Entomologist, 22, 173–183. [Google Scholar]
- Ashadul, M. I. , Hussain, M. A. , Shapla, S. A. , Mehraj, H. , & Uddin, A. F. M. J. (2014). Plant extract for the management of brinjal shoot and fruit borer (Leucinodes orbonalis Guenee). American‐Eurasian Journal of Agricultural and Environmental Sciences, 14, 1409–1414. [Google Scholar]
- Attygalle, A. B. , Schwarz, J. , & Gunawardena, N. E. (1988). Sex‐pheromone of brinjal shoot and pod borer Leucinodes orbonalis Guenee (Lepidoptera, Pyralidae, Pyraustinae) Zeitschrift fur Naturforschung C‐a. Journal of Biosciences, 43, 790–792. [Google Scholar]
- Atwal, A. S. (1976). Agricultural pests in India and Southeast Asia (p. 502). Kalyani Publishers. [Google Scholar]
- Bade, B. A. , Kharbade, S. B. , Kadam, M. B. , & Patil, A. S. (2017). Bio‐efficacy of novel pesticides against shoot and fruit borer, Leucinodes orbonalis (Guen.) on brinjal. Trends in Biosciences, 10, 1179–1182. [Google Scholar]
- Baidoo, P. K. , Mochiah, M. B. , Asare, D. , & Sefah, A. A. (2018). The role of soil amendments on population dynamics of insect pests, growth parameters and yield of eggplant, Solanum melongena (L.) Moench. Sustainable Agriculture Research, 7, 7–13. [Google Scholar]
- Baker, R. H. A. (2002). Predicting the limits to the potential distribution of alien crop pests. In Hallman G. J. & Schwalbe C. P. (Eds.), Invasive arthropods in agriculture: Problems and solutions (pp. 207–241). Science Publishers Inc. [Google Scholar]
- Bakhat, H. F. , Bibi, N. , Zia, Z. , Abbas, S. , Hammad, H. M. , Fahad, S. , Ashraf, M. R. , Shah, G. M. , Rabbani, F. , & Saeed, S. (2018). Silicon mitigates biotic stresses in crop plants: A review. Crop Protection, 104, 21–34. [Google Scholar]
- Baksh, S. (1979). Cytogenetic studies on the F1 hybrid Solanum incanum L. x Solanum melongena L. variety ‘Giant of Banaras’. Euphytica, 28, 793–800. [Google Scholar]
- Baksh, S. , & Iqbal, M. (1979). Compatibility relationships in some non‐tuberous species of Solanum . Journal of Horticultural Science, 54, 163–163. [Google Scholar]
- Banerjee, S. N. , & Basu, A. N. (1955). On the control of brinjal stem borer and fruit borer Leucinodes orbonalis Guen. In West Bengal. Science and Culture, 20, 350–351. [Google Scholar]
- Baskaran, P. , & Kumar, A. (1980). Further studies on Dipel‐insecticide combinations against the insect pests of brinjal. Pesticides, 14, 9–11. [Google Scholar]
- Behera, T. K. , & Gyanendra, S. (2002). Studies on resistance to shoot and fruit borer (Leucinodes orbonalis) and interspecific hybridization in eggplant. Indian Journal of Horticulture, 59, 62–66. [Google Scholar]
- Bhandari, P. , Agarwal, M. , Tomar, A. , & Rai, M. K. (2015). Bio‐efficacy and Phytotoxicity studies of Emamectin benzoate 5% Wsg against in Brinjal fruit and shoot borer. Annals of Horticulture, 8, 194–197. [Google Scholar]
- Bharati, M. S. , Shetgar, S. S. , Dongarjal, R. P. , & Kadam, A. R. (2015). Persistence toxicity of insecticides in brinjal. Indian Journal of Entomology, 77, 271–277. [Google Scholar]
- Biswas, K. , Mallikarjunappa, S. , & Bhat, U. G. (2009). Takumi‐an ultimate new generation product for the management of most problematic insect pest, shoot and fruit borer, Leucinodes orbonalis (Guen) of Brinjal. Pestology, 33, 23–25. [Google Scholar]
- BOJA , 2015. Boletin Oficial de la Junta de Andalucia Num. 248 de 24 de diciembre 2015. https://www.juntadeandalucia.es/boja/2015/248/BOJA15‐248‐00105‐21289‐01_00082170.pdf
- Boopal, K. , Arya, V. V. , Reddy, R. , & Hanur, V. S. (2013). Potato a suitable alternative host for brinjal fruit shoot and fruit borer Leucinodes orbonalis Guenee (Lepidoptera: Pyralidae). International Journal of Agricultural Science and Research, 3, 179–184. [Google Scholar]
- Brown, S. D. , Collins, R. A. , Boyer, S. , Lefort, M. C. , Malumbres‐Olarte, J. A. , Vink, C. J. , & Cruickshank, R. H. (2012). Spider: An R package for the analysis of species identity and evolution, with particular reference to DNA barcoding. Molecular Ecology Resources, 12, 562–565. [DOI] [PubMed] [Google Scholar]
- CABI . (2010). Solanum melongena datasheet. https://www.cabi.org/cpc/datasheet/50536
- Caley, P. , Ingram, R. , & De Barro, P. (2015). Entry of exotic insects into Australia: Does border interception count match incursion risk? Biological Invasions, 17, 1087–1094. [Google Scholar]
- CargoHandbook.com Eggplants . 2000. https://cargohandbook.com/Eggplants#:~:text=Fruit%20are%20stored%20at%2010,removal%20to%20typical%20retail%20conditions.
- CBI . (2020). Entering the European market for aubergine. https://www.cbi.eu/market‐information/fresh‐fruit‐vegetables/aubergine/market‐entry
- Chakraborti, S. , & Priyadarshini, S. (2021). Assessing impact of refuge planting and under‐storey repellent crops on incidence and damage of brinjal fruit and shoot borer. Journal of Entomological Research, 45, 714–719. [Google Scholar]
- Challa, N. , Singh, M. , Bharadwaj, R. K. , Sharma, R. , Gaikwad, M. B. , & Thakur, P. (2021). Characterization of eggplant genotypes for different resistance mechanisms against Leucinodes orbonalis . Neotropical Entomology, 50, 643–653. [DOI] [PubMed] [Google Scholar]
- Chandi, A. K. , & Kaur, A. (2021). Evaluation of insecticidal toxicity against Leucinodes orbonalis Guenee populations from Punjab. Agricultural Research Journal, 58, 344–347. [Google Scholar]
- Chandi, A. K. , Kaur, A. , & Chandi, R. S. (2022). Influence of temperature variation on emamectin benzoate toxicity in different Leucinodes orbonalis Guenee populations of Punjab. Indian Journal of Ecology, 49, 208–211. [Google Scholar]
- Chandre Gowda, B. A. (1980). Estimation of loss in brinjal due to shoot and fruit borer, Leucinodes orbonalis Guenee (Lepidoptera: Pyralidae) and evaluation of some new insecticides for its control. Mysore Journal of Agricultural Sciences, 14, 263–263. [Google Scholar]
- Chang, J. C. , Ponnath, D. W. , & Ramasamy, S. (2014). Phylogeographical structure in mitochondrial DNA of eggplant fruit and shoot borer, Leucinodes orbonalis Guenée (Lepidoptera: Crambidae) in south and Southeast Asia. Mitochondrial DNA Part A DNA Mapping, Sequencing, and Analysis, 27, 198–204. [DOI] [PubMed] [Google Scholar]
- Chatterjee, M. L. , & Roy, S. (2004). Bioefficacy of some insecticides against brinjal shoot and fruit borer (Leucinodes orbonalis Guenee) and effect of novaluron on natural enemies of brinjal pests. Pestology, 28, 52–56. [Google Scholar]
- Chi, N. M. , Quang, D. N. , Hien, B. D. , Dzung, P. N. , Nhung, N. P. , Nam, N. V. , Thuy, P. T. T. , Tuong, D. V. , & Dell, B. (2021). Management of Hypsipyla robusta Moore (Pyralidae) damage in Chukrasia tabularis a. Juss (Meliaceae). International Journal of Tropical Insect Science, 41, 2341–2350. [Google Scholar]
- Chowdhury, M. A. H. , Alam, M. Z. , Hossain, M. S. , Suh, S. J. , & Kwon, Y. J. (2017). Management of brinjal shoot and fruit borer Leucinodes orbonalis Guenee using different approaches. Bangladesh Journal of Agricultural Research, 42, 539–548. [Google Scholar]
- Čokl AA and Millar JG, 2009. Manipulation of insect signalling for monitoring and control of pest insects. 279–316.
- Coleman, M. , Kristiansen, P. , Sindel, B. , & Fyfe, C. (2020). Blackberry nightshade (Solanum nigrum): Weed management guide for Australian vegetable production. School of Environmental and Rural Science, University of New England, Armidale. [Google Scholar]
- Collonnier, C. , Fock, I. , Kashyap, V. , Rotino, G. L. , Daunay, M. C. , Lian, Y. , Mariska, I. K. , Rajam, M. V. , Servaes, A. , Ducreux, G. , & Sihachakr, D. (2001). Applications of biotechnology in eggplant. Plant Cell, Tissue and Organ Culture, 65, 91–107. [Google Scholar]
- Cork, A. , Alam, N. , Das, A. , Das, C. S. , Ghosh, G. C. , Farman, D. I. , Hall, D. R. , Maslen, N. R. , Vedham, K. , Phythian, S. J. , Rouf, F. M. A. , & Srinivasan, K. (2001). Female sex pheromone of brinjal fruit and shoot borer, Leucinodes orbonalis blend optimization. Journal of Chemical Ecology, 27, 1867–1877. [DOI] [PubMed] [Google Scholar]
- Cork, A. , Alam, S. N. , Rouf, F. M. A. , & Talekar, N. S. (2003). Female sex pheromone of brinjal fruit and shoot borer, Leucinodes orbonalis (Lepidoptera:Pyralidae): Trap optimization and application in IPM trials. Bulletin of Entomological Research, 93, 107–113. [DOI] [PubMed] [Google Scholar]
- Cork, A. , Alam, S. N. , Rouf, F. M. A. , & Talekar, N. S. (2005). Development of mass trapping technique for control of brinjal shoot and fruit borer, Leucinodes orbonalis (Lepidoptera:Pyralidae). Bulletin of Entomological Research, 95, 589–596. [DOI] [PubMed] [Google Scholar]
- Cork, A. , & Hall, D. R. (1998). Application of pheromones for crop pest management in the Indian sub‐continent. Journal of Asia‐Pacific Entomology, 1, 35–49. [Google Scholar]
- Das, M. S. , & Patnaik, B. H. (1971). A new host of the brinjal shoot and fruit borer Leucinodes orbonalis Guen, and its biology. Journal of the Bombay Natural History Society, 67(3), 601–603. [Google Scholar]
- Das, S. , Patel, J. I. , & Wazire, N. S. (2014). Non‐chemical management of Leucinodes orbonalis Guen. Ecology, Environment and Conservation, 20, 1075–1079. [Google Scholar]
- Dash, A. N. , & Singh, B. R. (1990). Field reaction of brinjal varieties against shoot and fruit borer Leucinodes orbonalis Guen. (Pyraustidae: Lepidoptera). Environment and Ecology, 8, 761–762. [Google Scholar]
- Dattatray, S. , Mangesh, P. , Bharat, C. & Srinivas, P. (2018). Selection and inheritance of Cry1Ac resistance in eggplant fruit and shoot borer (Leucinodes orbonalis). International Journal of Tropical Agriculture, 36, 159–165. [Google Scholar]
- David, A. L. (1966). Trials with insecticides for the control of brinjal leaf‐beetle (Epilacha vigintioctopunctata Fabr.) and fruit‐borer (Leucinodes orbonalis Guen.). Indian Journal of Agricultural Sciences, 36. [Google Scholar]
- Degri, M. M. (2014). The effect of spacing of eggplant (Solanum melongena L.) (Solanaceae) on shoot and fruit borer (Leucinodes orbonalis Guen.) (Lepidoptera:Pyralidae) infestation in the dry savanna zone of Nigeria. Agriculture and Biology Journal of North America, 5, 10–14. [Google Scholar]
- Dehariya, S. K. , Shukla, A. , & Barde, S. K. (2017). Efficacy of botanical pesticides against shoot and fruit borer, Leucinodes orbonalis in brinjal. Biosciences, Biotechnology Research Asia, 14, 721–725. [Google Scholar]
- Deore, S. P. , & Patil, R. S. (1996). Chemical control schedule for brinjal shoot and fruit borer. Journal of Maharashtra Agricultural Universities, 20, 416–419. [Google Scholar]
- Deotale, R. O. , Peshkar, L. N. , Dawane, P. N. , & Gawande, R. B. (2000). Effect of flufenoxuron and teflubenzuron alone and in combination with other insecticides on the management of Leucinodes orbonalis Guen. On brinjal. Journal of Soils and Crops, 10, 256–259. [Google Scholar]
- Dhaliwal, N. K. , & Aggarwal, N. (2021). Development and survival of brinjal shoot and fruit borer Leucinodes orbonalis Guenee (Crambidae: Lepidoptera) at constant and alternating temperatures. International Journal of Tropical Insect Science, 41, 1717–1728. [Google Scholar]
- Dhamdhere, S. V. , & Sharma, N. K. (1991). Chemical control of brinjal shoot and fruit borer Leucinodes orbonalis Guen. Journal of Entomological Research (New Delhi), 15, 109–112. [Google Scholar]
- Dhankar, B. S. (1988). Progress in resistance studies in the eggplant (Solanum melongena L.) against shoot and fruit borer (Leucinodes orbonalis guen.) infestation. Tropical Pest Management, 34, 343–345. [Google Scholar]
- Dhankhar, B. S. , Mehrotra, N. , Chaudhary, B. D. , & Pandita, M. L. (1979). Studies in genetic diversity for shoot and fruit borer (Leucinodes orbonalis Gn.) in normal and ratoon crop of brinjal (Solanum melongena L.). Haryana Journal of Horticultural Sciences, 8, 107–111. [Google Scholar]
- Douma, J. C. , Pautasso, M. , Venette, R. C. , Robinet, C. , Hemerik, L. , Mourits, M. C. M. , Schans, J. , & van der Werf, W. (2016). Pathway models for analysing and managing the introduction of alien plant pests – an overview and categorization. Ecological Modelling, 339, 58–67. 10.1016/j.ecolmodel.2016.08.009 [DOI] [Google Scholar]
- Ebert, A. W. (2017). Vegetable production, diseases, and climate change. World Agricultural Resources and Food Security, 17, 103–124. [Google Scholar]
- Ebert, A. W. , & Wu, T. H. (2011). Standardized protocol for eggplant seed regeneration and seed storage at AVRDC. The World Vegetable Center, 898, 81–87. [Google Scholar]
- EFSA (European Food Safety Authority) . (2014). Guidance on expert knowledge elicitation in food and feed safety risk assessment. EFSA Journal, 12(6), 3734. 10.2903/j.efsa.2014.3734 [DOI] [Google Scholar]
- EFSA (European Food Safety Authority) , & Maiorano, A. (2022). SCAN‐Clim: A tool to support pest climate suitability analysis based on climate classification. EFSA Journal, 20(2), 7104. 10.2903/j.efsa.2022.7104 [DOI] [PMC free article] [PubMed] [Google Scholar]
- EFSA PLH Panel (EFSA Panel on Plant Health) , Jeger, M. , Bragard, C. , Caffier, D. , Candresse, T. , Chatzivassiliou, E. , Dehnen‐Schmutz, K. , Gilioli, G. , Gregoire, J.‐C. , Jaques Miret, J. A. , MacLeod, A. , Navajas Navarro, M. , Niere, B. , Parnell, S. , Potting, R. , Rafoss, T. , Rossi, V. , Van Bruggen, A. , Van Der Werf, W. , … Urek, G. (2016a). Scientific opinion on the risk to plant health of Ditylenchus destructor for the EU territory. EFSA Journal, 14(12), 4602. 10.2903/j.efsa.2016.4602 [DOI] [Google Scholar]
- EFSA PLH Panel (EFSA Panel on Plant Health) , Jeger, M. , Bragard, C. , Chatzivassiliou, E. , Dehnen‐Schmutz, K. , Gilioli, G. , Jaques Miret, J. A. , Mac Leod, A. , Navajas Navarro, M. , Niere, B. , Parnell, S. , Potting, R. , Rafoss, T. , Urek, G. , Van Bruggen, A. , Van der Werf, W. , West, J. , Winter, S. , Maresi, G. , … Rossi, V. (2016b). Scientific opinion on the risk assessment and reduction options for Cryphonectria parasitica in the EU. EFSA Journal, 14(12), 4641. 10.2903/j.efsa.2017.4878 [DOI] [Google Scholar]
- EFSA PLH Panel (EFSA Panel on Plant Health) , Jeger, M. , Bragard, C. , Caffier, D. , Candresse, T. , Chatzivassiliou, E. , Dehnen‐Schmutz, K. , Gilioli, G. , Gregoire, J.‐C. , Jaques Miret, J. A. , MacLeod, A. , Niere, B. , Parnell, S. , Potting, R. , Rafoss, T. , Rossi, V. , Urek, G. , Van Bruggen, A. , Van Der Werf, W. , … Navajas Navarro, M. (2017a). Scientific Opinion on the pest risk assessment of Eotetranychus lewisi for the EU territory. EFSA Journal, 15(10), 4878. 10.2903/j.efsa.2017.4878 [DOI] [Google Scholar]
- EFSA PLH Panel (EFSA Panel on Plant Health) , Jeger, M. , Bragard, C. , Caffier, D. , Candresse, T. , Chatzivassiliou, E. , Dehnen‐Schmutz, K. , Gilioli, G. , Gregoire, J.‐C. , Jaques Miret, J. A. , MacLeod, A. , Navajas Navarro, M. , Niere, B. , Parnell, S. , Potting, R. , Rafoss, T. , Rossi, V. , Van Bruggen, A. , Van Der Werf, W. , … Urek, G. (2017b). Scientific opinion on the pest risk assessment of Radopholus similis for the EU territory. EFSA Journal, 15(8), 4879. 10.2903/j.efsa.2017.4879 [DOI] [Google Scholar]
- EFSA PLH Panel (EFSA Panel on Plant Health) , Jeger, M. , Bragard, C. , Caffier, D. , Candresse, T. , Chatzivassiliou, E. , Dehnen‐Schmutz, K. , Gregoire, J.‐C. , Jaques Miret, J. A. , MacLeod, A. , Navajas‐Navarro, M. , Niere, B. , Parnell, S. , Potting, R. , Rafoss, T. , Rossi, V. , Urek, G. , Van Bruggen, A. , Van Der Werf, W. , … Gilioli, G. (2018). Guidance on quantitative pest risk assessment. EFSA Journal, 16(8), 5350. 10.2903/j.efsa.2018.5350 [DOI] [PMC free article] [PubMed] [Google Scholar]
- EFSA PLH Panel (EFSA Panel on Plant Health) , Bragard, C. , Di Serio, F. , Gonthier, P. , Jaques Miret, J. A. , Justesen, A. F. , Magnusson, C. S. , Milonas, P. , Navas‐Cortes, J. A. , Parnell, S. , Potting, R. , Reignault, P. L. , Thulke, H.‐H. , Van der Werf, W. , Vicent Civera, A. , Yuen, J. , Zappala, L. , Gregoire, J.‐C. , Malumphy, C. , … MacLeod, A. (2021). Scientific opinion on the pest categorisation of Leucinodes orbonalis . EFSA Journal, 19(11), 6890. 10.2903/j.efsa.2021.6890 [DOI] [PMC free article] [PubMed] [Google Scholar]
- EFSA PLH Panel (EFSA Panel on Plant Health) , Bragard, C. , Baptista, P. , Chatzivassiliou, E. , Di Serio, F. , Gonthier, P. , Jaques Miret, J. A. , Fejer Justesen, A. , MacLeod, A. , Magnusson, C. S. , Navas‐Cortes, J. A. , Parnell, S. , Potting, R. , Reignault, P. L. , Stefani, E. , Thulke, H. H. , Vicent Civera, A. , Van der Werf, W. , Yuen, J. , … Milonas, P. (2023a). Assessment of the probability of introduction of Thaumatotibia leucotreta into the European Union with import of cut roses. EFSA Journal, 21(10), 8107. 10.2903/j.efsa.2023.8107 [DOI] [PMC free article] [PubMed] [Google Scholar]
- EFSA PLH Panel (EFSA Panel on Plant Health) , Bragard, C. , Baptista, P. , Chatzivassiliou, E. , Di Serio, F. , Gonthier, P. , Jaques Miret, J. A. , Justesen, A. F. , MacLeod, A. , Magnusson, C. S. , Milonas, P. , Navas‐Cortes, J. A. , Parnell, S. , Potting, R. , Reignault, P. L. , Stefani, E. , Thulke, H.‐H. , Vicent Civera, A. , Yuen, J. , … Van der Werf, W. (2023b). Scientific opinion on the pest risk assessment of Elasmopalpus lignosellus for the European Union. EFSA Journal, 21(5), 8004. 10.2903/j.efsa.2023.8004 [DOI] [Google Scholar]
- EFSA Scientific Committee , Benford, D. , Halldorsson, T. , Jeger, M. J. , Knutsen, H. K. , More, S. , Naegeli, H. , Noteborn, H. , Ockleford, C. , Ricci, A. , Rychen, G. , Schlatter, J. R. , Silano, V. , Solecki, R. , Turck, D. , Younes, M. , Craig, P. , Hart, A. , Von Goetz, N. , … Hardy, A. (2018). Guidance on uncertainty analysis in scientific assessments. EFSA Journal, 16(1), 5123. 10.2903/j.efsa.2018.5123 [DOI] [PMC free article] [PubMed] [Google Scholar]
- EPPO . (2004). Good plant protection practice: Solanaceous crops under protected cultivation. EPPO Bulletin, 34, 65–77. [Google Scholar]
- FAO (Food and Agriculture Organization of the United Nations) . (2018). ISPM (International standards for Phytosanitary measures) 5. Glossary of phytosanitary terms. https://www.ippc.int/static/media/files/publication/en/2018/06/ISPM_05_2018_En_Glossary_2018‐05‐20_PostCPM13_R9GJ0UK.pdf [Google Scholar]
- Fasihur, R. , Asim, G. , Munir, A. , Tariq, M. , & Muhammad, A. (2017). Efficacy of new chemistry insecticides against brinjal shoot and fruit borer (Leucinodes orbonalis Guenee). Pakistan Entomologist, 39, 33–36. [Google Scholar]
- Fathi, S. A. A. (2022). The role of intercrops of eggplant and cowpea on the control of Leucinodes orbonalis Guenee (Lepidoptera: Crambidae). BioControl, 67, 307–317. [Google Scholar]
- Ferreira, F. V. , & Musumeci, M. A. (2021). Trichoderma as biological control agent: Scope and prospects to improve efficacy. World Journal of Microbiology and Biotechnology, 37. [DOI] [PubMed] [Google Scholar]
- Fick, S. E. , & Hijmans, R. J. (2017). WorldClim 2: New 1‐km spatial resolution climate surfaces for global land areas. International Journal of Climatology, 37(12), 4302–4315. [Google Scholar]
- FPJ . (2011). re:Fresh Directory, 2011. Fresh Produce Journal. [Google Scholar]
- Funde T , Bhalkare, S. K. , Undirwade, D. B. , & Satpute, N. S. (2020). Performance of Trichogramma chilonis Ishii on different hosts. Journal of Biological Control, 34, 208–214. [Google Scholar]
- Gahukar, R. T. , & Balpande, P. B. (1997). Field evaluation of a new neem‐based formulation against major insect pests of brinjal. Pestology, 11, 14–18. [Google Scholar]
- Gayathri, S. , & Geetha, B. (2020). Evaluation of botanicals and biopesticides against major insect pests of brinjal. Indian Journal of Entomology, 82, 864–870. [Google Scholar]
- GBIF.org . (2023). GBIF occurrence download Leucinodes laisalis, Europe. 10.15468/dl.hg3263 [DOI]
- Ghatak, S. S. , Mondal, S. , & Vishwakarma, R. (2009). Bioefficacy of botanicals and biopesticides against brinjal shoot and fruit borer, Leucinodes orbonalis Guen. (Pyraustidae: Lepidoptera). Indian Journal of Entomology, 71, 284–287. [Google Scholar]
- Gholami, Z. , & Sadeghi, A. (2016). Management strategies for western flower thrips in vegetable greenhouses in Iran: A review. Plant Protection Science, 52, 87–98. [Google Scholar]
- Ghorai, A. K. (2005). Avoidance of brinjal fruit borer (Leucinodes orbonalis Guenn.) through eco‐friendly physical barriers. Journal of Entomological Research (New Delhi), 29, 127–129. [Google Scholar]
- Ghosal, A. , Chatterjee, M. L. , & Bhattacharya, A. (2013). Evaluation of some biorational pesticides for the management of shoot and fruit borer of brinjal and okra. Pesticide Research Journal, 25, 146–151. [Google Scholar]
- Ghosh, S. K. , & Senapati, S. K. (2009). Seasonal fluctuation in the population of Leucinodes orbonalis Guen. In the sub‐Himalayan region of West Bengal, India and its control on eggplant (Solanum melongena L.). Precision Agriculture, 10, 443–449. [Google Scholar]
- Ghosh, S. K. , & Pal, S. (2016). Entomopathogenic potential of Trichoderma longibrachiatum and its comparative evaluation with malathion against the insect pest Leucinodes orbonalis . Environmental Monitoring and Assessment, 188(1). [DOI] [PubMed] [Google Scholar]
- Gilligan, T. M. , & Passoa, S. C. (2014). Leucinodes orbonalis. In LepIn–ercept–An identification resource for intercepted Lepidoptera larvae. https://idtools.org/id/lepintercept/orbonalis.html [Google Scholar]
- Gould, J. R. , & Maldonado, M. H. (2006). Copitarsia decolora (Lepidoptera: Noctuidae) larvae escaping from discarded asparagus: Data in support of a pathway risk analysis. Journal of Economic Entomology, 99, 1605–1609. 10.1093/jee/99.5.1605 [DOI] [PubMed] [Google Scholar]
- Gould, J. R. , Caton, B. P. , & Venette, R. C. (2006). A pathway assessment of the risk of establishment in the contiguous United States by Copitarsia decolora (Guenne) on Asparagus from Peru. USDA APHIS. Available online: https://www.yumpu.com/en/document/read/27534062/copitarsia‐asparagus‐peru‐original
- Goundalkar, M. J. , & Webster, F. X. (2007). Recent advances in the chemistry of insect pheromones. 285–349.
- Gowda, J. V. N. , Sulladmath, U. V. , Rao, K. P. G. , Narayana Gonda, J. V. , Gopalkrishna Rao, K. P. , & Sulladmath, U. V. (1977). Phenthoate–a new effective phosphatic insecticide against cabbage diamond moth, brinjal borer and onion thrips. University of Agricultural Sciences Bangalore Current Research, 6, 42–43. [Google Scholar]
- Gowda, P. H. R. , Shivashankar, K. T. , & Joshi, S. (1990). Interspecific hybridization between Solanum melongena and Solanum macrocarpon: Study of the F1 hybrid plants. Euphytica, 48, 59–61. [Google Scholar]
- Gowrish, K. R. , Ramesha, B. , & Ushakumari, R. (2015). Biorational management of major pests of brinjal. Indian Journal of Entomology, 77, 51–55. [Google Scholar]
- Grimm, V. , & Wissel, C. (1997). Babel, or the ecological stability discussions: An inventory and analysis of terminology and a guide for avoiding confusion. Oecologia, 109, 323–334. [DOI] [PubMed] [Google Scholar]
- Gül, M. , Değirmenci, N. , Şirikçi, B. S. , & Kadakoğlu, B. (2022). Cost and profitability analysis of greenhouse eggplant production: A case study of Antalya Province, Turkey. Custos e Agronegocio Online, 18, 440–457. [Google Scholar]
- Gulati, P. , Chauhan, S. , Mubayi, A. , Singh, T. , & Rana, P. (2021a). Dynamical analysis, optimum control and pattern formation in the biological pest (EFSB) control model (p. 147). [Google Scholar]
- Gulati, P. , Chauhan, S. , Mubayi, A. , Singh, T. , & Rana, P. (2021b). Dynamical analysis, optimum control and pattern formation in the biological pest (EFSB) control model chaos. Solitons & Fractals, 147, 110920. [Google Scholar]
- Gunawardena, N. E. , Attygalle, A. B. , & Bandara Herath, H. M. W. K. (1989). The sex pheromone of the brinjal pest, Leucinodes orbonalis Guenee (Lepidoptera): Problems and perspectives. Journal of the National Science Council of Sri Lanka, 17, 161–171. [Google Scholar]
- Hanapur, R. H. , & Nandihalli, B. S. (2004). Bio‐efficacy of certain insecticides and botanicals against the potato shoot borer, Leucinodes orbonalis Guenee. Karnataka Journal of Agricultural Sciences, 17, 602–603. [Google Scholar]
- Hanur, V. S. , Reddy, P. V. R. , Boopal, K. , Srividya, K. N. , Poovarasan, S. , Bhalewar, S. , & Saraswathi, M. S. (2011). Solanum macrocarpon, a wild brinjal, is not a source of resistance against brinjal shoot and fruit borer, Leucinodes orbonalis Guenee. Current Science, 101, 1427–1429. [Google Scholar]
- Haque, M. S. , & Saha, N. R. (2020). Biosafety measures, socio‐economic impacts and challenges of Bt‐brinjal cultivation in Bangladesh. Frontiers in Bioengineering and Biotechnology, 8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hasan, W. U. , Kumar, A. , & Akhtar, N. , (2021). Use of Bt‐gene in the Solanum melongena L. and its biological and social consequences. 363–374.
- Hautea, D. M. , Taylo, L. D. , Masanga, A. P. L. , Sison, M. L. J. , Narciso, J. O. , Quilloy, R. B. , Hautea, R. A. , Shotkoski, F. A. , & Shelton, A. M. (2016). Field performance of Bt eggplants (Solanum melongena L.) in The Philippines: Cry1Ac expression and control of the eggplant fruit and shoot borer (Leucinodes orbonalis Guenee). PLoS One, 11, e0157498. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hayden, J. E. , Lee, S. , Passoa, S. C. , Young, J. , Landry, J.‐F. , Nazari, V. , Mally, R. , Somma, L. A. , & Ahlmark, K. M. (2013). Digital identification of Microlepidoptera on Solanaceae. Fort Collins, CO. https://idtools.org/id/leps/micro/ [Google Scholar]
- Hebert, P. D. N. , Cywinska, A. , Ball, S. L. , & de Waard, J. R. (2003). Biological identifications through DNA barcodes. Proceedings of the Royal Society B, 270, 313–321. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heinrichs, E. A. , & Muniappan, R. (2016). IPM for food and environmental security in the tropics. 1–31.
- Herms, D. A. , & McCullough, D. G. (2014). Emerald ash borer invasion of North America: History, biology, ecology, impact and management. Annual Review of Entomology, 59, 13–30. [DOI] [PubMed] [Google Scholar]
- Hill, D. S. (1987). Agricultural insect pests of temperate regions and their control (p. 659). Cambridge University Press. [Google Scholar]
- Huertas‐Dionisio, M. (2000). Estados inmaturos de Lepidoptera (XII). Tres especies de origen tropical de la subfamilia Pyraustinae Meyrick, 1890 (Lepidoptera: Pyraloidea, Crambidae). SHILAP Revista de lepidopterología, 28, 321–334. [Google Scholar]
- Hussaini, S. S. (2014). Potential of Entomopathogenic nematodes in integrated Pest management. 193–223.
- Hussaini, S. S. , Singh, S. P. , & Nagesh, M. (2002). In vitro and field evaluation of some indigenous isolates of Steinernema and Heterorhabditis indica against shoot and fruit borer, Leucinodes orbonalis . Indian Journal of Nematology, 32, 63–65. [Google Scholar]
- Hutson, J. C. (1931). Report of Insect Pests in Ceylon during 1930.
- Ilmu Majlis, 2018. Knowledge convention. Harvesting and post handling. https://www.majlisilmu.gov.bn/Shared%20Documents/Pameran/2018/WEEK%208%20Post%20Harvest%20Handling.pdf [Google Scholar]
- INaturalist . (2023). Brown Tipped Pearl (Leucinodes laisalis). https://www.inaturalist.org/observations?taxon_id=782633
- Indian Council of Agricultural Research . (2016). Biology of brinjal. https://biosafety.icar.gov.in/biology‐of‐brinjal/
- Indian Council of Agricultural Research . (undated). Package of practices for tomato cultivation. https://kvk.icar.gov.in/API/Content/PPupload/K0650_61.pdf
- Isahaque, N. M. M. , & Chadhuri, R. P. (1983). A new alternate host plant of brinjal shoot and fruit borer Leucinodes orbonalis Guen. In Assam. Journal of Research, Assam Agricultural University, 4, 83–85. [Google Scholar]
- Islam, M. A. , Bin Mohi Uddin, M. M. , Rasul, M. G. , Haque Swapon, M. A. , Ahmed, M. , & Hasan, M. (2022). In vitro screening and field performance of EMS‐treated eggplants for the selection of shoot and fruit borer‐resistant plants. Agronomy, 12(8), 1832. 10.3390/agronomy12081832 [DOI] [Google Scholar]
- Islam, T. , Roknuzzaman, A. H. M. , Hassan, K. , & Ullah, M. S. (2020). Temperature impacts on the eggplant shoot and fruit borer Leucinodes orbonalis: A life table approach. International Journal of Tropical Insect Science, 40, 351–360. [Google Scholar]
- Jalali, S. K. (2013). Natural occurrence, host range and distribution of trichogrammatid egg parasitoids. In Sithanantham S., Ballal C. R., Jalali S. K., & Bakthavatsalam N. (Eds.), Biological control of insect pests using egg parasitoids (pp. 67–76). Springer. 10.1007/978-81-322-1181-5_4 [DOI] [Google Scholar]
- Jayakrishnan, S. , & Madhuban, G. (2012). Evaluation of synthetic and natural insecticides for the management of insect pest control of eggplant (Solanum melongena L.) and pesticide residue dissipation pattern. American. Journal of Plant Sciences, 3(2), 17547. https://www.scirp.org/journal/PaperInformation.aspx?PaperID=17547 [Google Scholar]
- Jaydeep, H. , Totan, A. , & Sujan, M. (2022). Bioefficacy, phytotoxicity and insecticide residue dynamics of chlorantraniliprole in brinjal (Solanum melongena) under field condition. Indian Journal of Agricultural Sciences, 92, 1219–1224. [Google Scholar]
- Jethva, D. M. , & Vyas, H. J. (2009). Studies on life fecundity tables of Leucinodes orbonalis Guenee on tomato. Indian Journal of Entomology, 71, 143–147. [Google Scholar]
- Joshi, M. L. , & Sharma, J. C. (1973). Insecticides for the control of brinjal aphid Aphis gossypii, jassid Amrasca devastans and fruit borer Leucinodes orbonalis in Rajasthan India. Indian Journal of Agricultural Sciences, 43, 436–438. [Google Scholar]
- Kadam, J. R. , Bhosale, U. D. , & Chavan, A. P. (2006). Influence of insecticidal treatment sequences on population of Leucinodes orbonalis Gn and its predators. Journal of Maharashtra Agricultural Universities, 31, 379–382. [Google Scholar]
- Kalia, V. K. , Tanu, S. , & Gujar, G. T. (2013). Susceptibility of brinjal shoot and fruit borer, Leucinodes orbonalis (Guenee) to bacillus thuringiensis and its cry toxins. Biopesticides International, 9, 88–92. [Google Scholar]
- Kariyanna, B. , Prabhuraj, A. , Asokan, R. , Babu, P. , Jalali, S. K. , Venkatesa, T. , Gracy, R. G. , & Mohan, M. (2020a). Identification of suitable reference genes for normalization of RT‐qPCR data in eggplant fruit and shoot borer (Leucinodes orbonalis Guenée). Biologia, 75, 289–297. [Google Scholar]
- Kariyanna, B. , Prabhuraj, A. , Mohan, M. , Bheemanna, M. , Basavaraj, K. , Pampanna, Y. , & Diwan, J. R. (2020b). Insecticide usage pattern and evolution of resistance in eggplant shoot and fruit borer, Leucinodes orbonalis Guenée (Lepidoptera: Crambidae) in India. Plant Archives, 20(Suppl. 2), 1255–1261. [Google Scholar]
- Karthika, P. , Vadivalagan, C. , Thirumurugan, D. , & Murugan, K. (2019). Intra‐species variation and geographic differentiation among the populations of the quarantine agricultural pest Leucinoides orbonalis (Lepidoptera: Crambidae) in the global assemblage – A prospective of DNA barcoding. Mitochondrial DNA Part A DNA Mapping, Sequencing, and Analysis, 30(4), 682–693. 10.1080/24701394.2019.1622691 [DOI] [PubMed] [Google Scholar]
- Kashyap, V. , Vinod Kumar, S. , Collonnier, C. , Fusari, F. , Haicour, R. , Rotino, G. L. , Sihachakr, D. , & Rajam, M. V. (2003). Biotechnology of eggplant. Scientia Horticulturae, 97, 1–25. [Google Scholar]
- Kassi, A. K. , Javed, H. , & Mukhtar, T. (2019). Screening of different Aubergine cultivars against infestation of Brinjal fruit and shoot borer (Leucinodes orbonalis Guenee) Pakistan. Journal of Zoology, 51, 603–609. [Google Scholar]
- Kaur, A. , Saini, M. K. , Sharma, S. , & Kooner, R. (2022). Novel molecules targeted control of Brinjal shoot and fruit borer, Leucinodes orbonalis in subtropical conditions of Punjab (India). Pesticide Research Journal, 34, 41–45. [Google Scholar]
- Kaur, J. , Kang, B. K. , & Singh, B. (2014). Base line data for insecticide resistance monitoring in brinjal shoot and fruit borer, Leucinodes orbonalis Guenee. Bioscan, 9, 1395–1398. [Google Scholar]
- Kaur, D. N. , & Aggarwal, N. (2019). Biointensive management of brinjal shoot and fruit borer Leucinodes orbonalis Guenee Indian. Journal of Entomology, 81(4), 760–764. [Google Scholar]
- Kaur, S. (2004). Testing of IPM modules for the management of brinjal pests. Indian Journal of Entomology, 66, 270. [Google Scholar]
- Kaur, S. , Bal, S. S. , Singh, G. , Sidhu, A. S. , & Dhillon, T. S. (2004). Management of brinjal shoot and fruit borer, Leucinodes orbonalis Guenee through net house cultivation. In: VII international symposium on protected cultivation in mild winter climates: Production, Pest management and global competition 659 (pp. 345–350).
- Kavitha, V. S. , Revathi, N. , & Kingsley, S. (2008). Biology of brinjal pest, Leucinodes orbonalis Guen. Of Erode region in Tamil Nadu. Journal of Entomological Research, 32(3), 255–257. [Google Scholar]
- Keatinge JDH, Easdown WJ, Hughes JDA and Tenkouano A, 2011. Small and medium‐scale public‐private partnerships in horticulture for development: An example fro– AVRDC–the world vegetable center in XXVIII international horticultural congress on science and horticulture for people (IHC2010): International symposium on 921. pp. 25–33.
- Kelly, G. , Napier, T. , & Watts, S. (2013). Gross margin budget Capsicum. NSW Dept of Primary Industries. [Google Scholar]
- Kenis, M. , Rabitsch, W. , Auger‐Rozenberg, M. A. , & Roques, A. (2007). How can alien species inventories and interception data help us prevent insect invasions? Bulletin of Entomological Research, 97, 489–502. [DOI] [PubMed] [Google Scholar]
- Khan, F. S. , & Borikar, P. S. (2017). Performance of Bt and non Bt brinjal hybrids against brinjal shoot and fruit borer, Leucinodes orbonalis Guenee. Journal of Entomological Research, 41, 283–286. [Google Scholar]
- Khatun, R. , Alam, K. , Rana, S. , Mashud, A. A. , Masud, A. A. , Ahmed, S. , Islam, R. , & Jamal, A. H. M. (2022). In vitro efficiency of crude extract of Ricinus communis, Abroma augusta, and Bombax ceiba seed on brinjal shoot and fruit borer, Leucinodes orbonalis Guenee. African Journal of Agricultural Research, 18, 73–79. [Google Scholar]
- Khorsheduzzaman, A. K. M. , Alam, M. Z. , Rahman, M. M. , Mian, M. A. K. , & Mian, M. I. H. (2010). Biochemical basis of resistance in eggplant (Solanum melongena L.) to Leucinodes orbonalis Guenee and their correlation with shoot and fruit infestation Bangladesh. Journal of Agricultural Research, 35, 149–155. [Google Scholar]
- Knapp, S. (2023). Solanum virginianum. Solanaceae Source–A global taxonomic resource for the nightshade family. https://solanaceaesource.myspecies.info/content/solanum‐virginianum
- Kodandaram, M. H. , Rai, A. B. , Sireesha, K. , & Halder, J. (2015). Efficacy of cyantraniliprole a new anthranilic diamide insecticide against Leucinodes orbonalis (Lepidoptera: Crambidae) of brinjal. Journal of Environmental Biology, 36, 1415–1420. [Google Scholar]
- Kottek, M. , Grieser, J. , Beck, C. , Rudolf, B. , & Rubel, F. (2006). World map of the Köppen‐Geiger climate classification updated. Meteorologische Zeitschrift, 15, 259–263. 10.1127/0941-2948/2006/0130 [DOI] [Google Scholar]
- Koundinya, A. V. V. , Pandit, M. K. , & Saha, A. (2019). Association of fruit and shoot borer infestation in eggplant with morphological, yield, quality and external weather variables. Agricultural Research, 8(3), 309–319. 10.1007/s40003-018-0380-0 [DOI] [Google Scholar]
- Krishnaiah, K. , Mohan, N. J. , & Prasad, V. G. (1981). Efficacy of Bacillus thuringiensis for the control of lepidopterous pests of vegetable crops. Entomon, 6, 87–93. [Google Scholar]
- Krishnamoorthy, S. V. , Banuchitra, R. , Devi, S. J. , & Karuppuchamy, P. (2014). Management of brinjal shoot and fruit borer Leucinodes orbonalis Guenee with novel Insecticides. Journal of Insect Science (Ludhiana), 27, 17–22. [Google Scholar]
- Krithika, B. N. , & Ananthanarayana, S. R. (2017). Role of abiotic factors and farming methods in the management of the pest Leucinodes orbonalis (Guenee) on Solanum melongena l. International Journal of Current Research, 9(12), 62253–62255. [Google Scholar]
- Kriticos, D. J. , Maywald, G. F. , Yonow, T. , Zurcher, E. J. , Herrmann, N. I. , & Sutherst, R. W. (2015). CLIMEX Version 4: Exploring the effects of climate on plants, animals and diseases (p. 184). CSIRO. [Google Scholar]
- Kriticos, D. J. , Webber, B. L. , Leriche, A. , Ota, N. , Macadam, I. , Bathols, J. , & Scott, J. K. (2012). CliMond: Global high resolution historical and future scenario climate surfaces for bioclimatic modelling. Methods in Ecology and Evolution, 3, 53–64. 10.1111/j.2041-210X.2011.00134.x [DOI] [Google Scholar]
- Kudlu, C. , & Stone, G. D. (2013). The trials of genetically modified food: Bt eggplant and ayurvedic medicine in India. Food, Culture and Society, 16, 21–42. [Google Scholar]
- Kumar, A. , & Shukla, A. (2002). Varietal preference of fruit and shoot borer, Leucinodes orbonalis Guen. On brinjal. Insect Environment, 8(1), 44. [Google Scholar]
- Kumar, B. , Anil Rao, B. , & Narasimha Sriramulu, M. (2000). Studies on the efficacy of certain insecticides and their mixtures against shoot and fruit borer, Leucinodes orbonalis Guen. On brinjal. Indian Journal of Plant Protection, 28, 25–28. [Google Scholar]
- Kumar, C. A. , Rao, G. R. , Chalam, M. S. V. , & Rao, P. S. (2012). Influence of Nutrients on Incidence of Pests of Solanum Melongena Linn. Annals of Plant Protection Sciences, 20(2), 302–305. [Google Scholar]
- Kumar, N. K. K. , & Sadashiva, A. T. (1991). Solanum macrocarpon: A wild species of brinjal resistant to brinjal shoot and fruit borer, Leucinodes orbonalis (Guen.). Insect. Environment, 2(2), 41–42. [Google Scholar]
- Kumar, P. A. , Mandaokar, A. , Sreenivasu, K. , Chakrabarti, S. K. , Bisaria, S. , Sharma, S. R. , Kaur, S. , & Sharma, R. P. (1998). Insect‐resistant transgenic brinjal plants. Molecular Breeding, 4, 33–37. [Google Scholar]
- Kumar, S. , Misra, A. , Verma, A. K. , Roy, R. , Tripathi, A. , Ansari, K. M. , Das, M. , & Dwivedi, P. D. (2011). Bt brinjal in India: A long way to go. GM Crops, 2, 92–98. [DOI] [PubMed] [Google Scholar]
- Kunjwal N and Srivastava RM, 2018. Insect pests of vegetables. pp. 163–221. In: Omkar (ed.), Pests and their management, Springer Singapore. 10.1007/978-981-10-8687-8_7 [DOI] [Google Scholar]
- Lal, O. P. (1975). Zur Überwinterung von Leucinodes orbonalis Guen. (Lep., Pyralidae), eines Schädlings der Aubergine, Solanum melongena L., im Kulu‐Tal, West‐Himalaya, Indien. Anzeiger für Schädlingskunde, Pflanzenschutz, Umweltschutz, 48, 181–182. [Google Scholar]
- Lalitha, Y. , Venkatesan, T. , & Jalali, S. K. (2013). Safety testing of pesticides for integration with trichogrammatids. In Sithanantham S., Ballal C., Jalali S., & Bakthavatsalam N. (Eds.), Biological control of insect pests using egg parasitoids (pp. 127–173). Springer. 10.1007/978-81-322-1181-5_7 [DOI] [Google Scholar]
- Lall, B. S. , & Ahmad, S. Q. (1965). The biology and control of brinjal (eggplant) fruit and shoot borer, Leucinodes orbonalis . Journal of Economic Entomology, 58(3), 448–451. 10.1093/jee/58.3.448 [DOI] [Google Scholar]
- Latif, M. A. , Rahman, M. M. , & Alam, M. Z. (2010). Efficacy of nine insecticides against shoot and fruit borer, Leucinodes orbonalis Guenee (Lepidoptera: Pyralidae) in eggplant. Journal of Pest Science, 83, 391–397. [Google Scholar]
- Latif, M. A. , Rahman, M. M. , Alam, M. Z. , Hossain, M. M. , & Solaiman, A. R. M. (2009). Threshold based spraying of flubendiamide for the management of brinjal shoot and fruit borer, Leucinodes orbonalis (Lep.: Pyralidae). Journal of Entomological Society of Iran, 29, 1–10. [Google Scholar]
- Lepiforum . (2023). Leucinodes laisalis (Walker, 1859). https://lepiforum.org/wiki/page/Leucinodes_laisalis
- Leung, B. , Drake, J. M. , & Lodge, D. M. (2004). Predicting invasions: Propagule pressure and the gravity of Allee effects. Ecology, 85, 1651–1660. [Google Scholar]
- Li, Y. , Yu, Y. , Chen, J. , Gao, Y. , Lu, F. , Tang, Y. , Huang, M. , Feng, B. , Fang, C. , & Li, H. (2014). A study on the developmental periods of eggplant fruit and shoot borers in Shanghai and analysis of its pathogenic factors. [in Chinese] China Plant Protection, 34(8), 51–54. [Google Scholar]
- Lobato‐Gómez, M. , Hewitt, S. , Capell, T. , Christou, P. , Dhingra, A. , & Girón‐Calva, P. S. (2021). Transgenic and genome‐edited fruits: Background, constraints, benefits, and commercial opportunities. Horticulture Research, 8, 166. 10.1038/s41438-021-00601-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mahata, S. , Das, B. C. , Patra, S. , Biswas, A. K. , Chatterjee, M. L. , & Samanta, A. (2014). New Diamide insecticides against fruit and shoot borer (Leucinodes orbonalis Guen.) in Brinjal. Pesticide Research Journal, 26, 197–201. [Google Scholar]
- Mahesh, P. , & Men, U. B. (2007). Bio‐efficacy of bacillus thuringiensis Berliner against Leucinodes orbonalis Guen. Indian Journal of Entomology, 69, 234–237. [Google Scholar]
- Mahla, M. K. , Kumar, A. , Lekha, A. V. , & Singh, V. (2017). Bio‐efficacy of carbosulfan 25 EC against fruit and shoot borer of brinjal and their natural enemies. Journal of Entomology and Zoology Studies, 5, 260–264. [Google Scholar]
- Mainali, R. P. , Thapa, R. B. , Tiwari, S. , Pokhrel, P. , & Ansari, A. R. (2014). Knowledge and practices on eggplant fruit and shoot borer, Leucinodes orbonalis Guenee management in Dhading and bara districts of Nepal Albanian. Journal of Agricultural Sciences, 13, 6–13. [Google Scholar]
- Mallik, S. N. , Kumar, M. , Sinha, A. N. , & Karn, B. P. (1989). Trathala flavoorbitalis Cameron (Ichneumonidae)–parasite of Leucinodes orbonalis Guen from Bihar. Current Science, 58, 1098–1099. [Google Scholar]
- Mally, R. , Hayden, J. E. , Neinhuis, C. , Jordal, B. H. , & Nuss, M. (2019). The phylogenetic systematics of Spilomelinae and Pyraustinae (Lepidoptera: Pyraloidea: Crambidae) inferred from DNA and morphology. Arthropod Systematics & Phylogeny, 77, 141–204. 10.26049/ASP77-1-2019-07 [DOI] [Google Scholar]
- Mally, R. , Korycinska, A. , Agassiz, D. J. L. , Hall, J. , Hodgetts, J. , & Nuss, M. (2015). Discovery of an unknown diversity of Leucinodes species damaging Solanaceae fruits in sub‐Saharan Africa and moving in trade (Insecta, Lepidoptera, Pyraloidea). ZooKeys, 472, 117–162. 10.3897/zookeys.472.8781 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Malshe, K. V. , Palshetkar, M. G. , Desai, B. G. , & Mane, S. B. (2016). Performance of different varieties of brinjal (Solanum melongena L.) under north Konkan conditions of Maharashtra, India. Plant Archives, 16, 568–571. [Google Scholar]
- Manasa, K. , & Gangadhar, N. (2018). Study on field longevity of pheromone lures against brinjal shoot and fruit borer Leucinodes orbonalis (Guenee) in relation with weather parameters. Trends in Biosciences, 11, 1954–1956. [Google Scholar]
- Manasa, K. , Gangadhar, N. , Kotikal, Y. K. , Haadimani, H. P. , Nadaf, A. M. , & Goud, C. R. (2016). Evaluation of different sex pheromone trap designs in catching male moths of brinjal shoot and fruit borer, Leucinodes orbonalis Guene (Lepidoptera:Crambidae). Journal of Entomological Research, 40, 35–37. [Google Scholar]
- Mandal, S. K. , Karam, N. , & Das, A. (2011). Bio‐efficacy of Rynaxypyr 20% SC (Chlorantraniliprole), a new molecule, against eggplant shoot and fruit borer. ISHS Acta Horticulturae 917: XXVIII international horticultural congress on science and horticulture for people (IHC2010): International symposium on plant protection, 917, 63–68. [Google Scholar]
- Mane, P. D. , & Kulkarni, S. N. (2010). Bio‐efficacy of neem products against Leucinodes orbonalis G. On brinjal. Green Farming, 1(2), 220. [Google Scholar]
- Manickam, P. , Kathirvelu, B. , Sundaram, J. , & Munusamy, A. (2014). Bio‐efficacy of crude leaf extracts of Acalypha fruticosa Forssk. Against some agriculturally important insect pests. Asian Pacific Journal of Tropical Disease, 4, S890–S894. [Google Scholar]
- Mannan, M. A. , Islam, K. S. , Jahan, M. , & Tarannum, N. (2015). Some biological parameters of brinjal shoot and fruit borer, Leucinodes orbonalis Guenee (Lepidoptera: Pyralidae) on potato in laboratory condition. Bangladesh Journal of Agricultural Research, 40(3), 381–390. [Google Scholar]
- Manoj, P. D. , & Ravi, G. (2019a). Studies on host sex pheromone interaction in parasitization behaviour of Trichogramma chilonis Ishii (Trichogrammatidae: Hymenoptera) in brinjal ecosystem. International Journal of Entomology Research, 4, 51–52. [Google Scholar]
- Manoj, P. D. , & Ravi, G. (2019b). Studies on influence of coriander Coriandrum sativum L. on moth recruitment pattern of trap design and validation of pheromone trap designs for the management of brinjal shoot and fruit borer, Leucinodes orbonalis Guenee (Crambidae: Lepidoptera). Journal of Entomological Research, 43, 131–133. [Google Scholar]
- Manoj, P. D. , Ravi, G. , & Sithanantham, S. (2018). Mating status of brinjal shoot and fruit borer Leucinodes orbonalis Guenee (Crambidae:Lepidoptera) male adults caught in sex pheromone traps. Entomon, 43, 291–292. [Google Scholar]
- MAPAMA . (2021). Ministerio de Agricultura, Pesca y Alimentacion. Available online: https://www.mapa.gob.es/es/
- Marimuthu, S. (2008). Effectiveness of insecticides in the control of brinjal shoot and fruit borer Leucinodes orbonalis (Pyralidae: Lepidoptera). Journal of Ecobiology, 23, 33–36. [Google Scholar]
- Mathur, A. , & Jain, N. (2006). A study on the biology of shoot and fruit borer Leucinodes orbonalis Guene (Lepidoptera: Pyralidae). Journal of Experimental Zoology India, 9, 225–228. [Google Scholar]
- Maureal, A. M. , Cariño, M. F. A. , Noriel, L. M. , & Esguerra, N. M. (1982). Life history and behavior of eggplant fruit borer, Leucinodes orbonalis Guenee. Annals of Tropical Research, 4, 178–188. [Google Scholar]
- Mazomenos, B. E. , Raptopoulos, D. , Lefkidou, I. , & Mazomenos‐Pantazi, A. (1994). Female sex pheromone components of jasmine moth Palpita unionalis (Lepidoptera: Pyralidae). Journal of Chemical Ecology, 20, 745–751. [DOI] [PubMed] [Google Scholar]
- Mazumder, F. , & Khalequzzaman, M. (2010). Eggplant shoot and fruit borer Leucinodes orbonalis Guenee male moth catch in sex pheromone trap with special reference of lure elevation and IPM. Journal of Bio‐Science, 18, 9–15. [Google Scholar]
- Mehta, P. K. , Vaidya, D. N. , & Kashyap, N. P. (1998). Bioefficacy of some insecticides against brinjal fruit and shoot borer, Leucinodes orbonalis (Guen.). Journal of Insect Science (Ludhiana), 11, 80–81. [Google Scholar]
- Mejía, C. , Ardila, H. D. , Espinel, C. , Brandão, P. F. B. , & Villamizar, L. (2021). Use of Trichoderma koningiopsis chitinase to enhance the insecticidal activity of Beauveria bassiana against Diatraea saccharalis . Journal of Basic Microbiology, 61, 814–824. [DOI] [PubMed] [Google Scholar]
- Mishra, N. C. , & Mishra, S. N. (1995). Insecticides for management of brinjal fruit and shoot borer Leucinodes orbonalis Guen. South Indian Horticulture, 43, 171–173. [Google Scholar]
- Mohan, N. J. , & Prasad, V. G. (1986). Role of synthetic pyrethroids in the control of brinjal pests. Indian Journal of Entomology, 46, 179–182. [Google Scholar]
- Muhammad, W. , Javed, H. , Ahmad, M. , & Mukhtar, T. (2021). Optimizing transplanting dates for the Management of Brinjal Shoot and Fruit Borer and better crop yield under field conditions Pakistan. Journal of Zoology, 53, 967–973. [Google Scholar]
- Muirhead, J. R. , Leung, B. , van Overdijk, C. , Kelly, D. W. , Nandakumar, K. , Marchant, K. R. , & MacIsaac, H. J. (2006). Modelling local and long‐distance dispersal of invasive emerald ash borer Agrilus planipennis (Coleoptera) in North America. Diversity and Distributions, 12, 71–79. [Google Scholar]
- Mukherjee, A. , & Ghosh, S. K. (2023). An eco‐friendly approach of biocontrol of aphid (Aphis gossypii Glover) by Trichoderma harzianum . Environmental Monitoring and Assessment, 195. [DOI] [PubMed] [Google Scholar]
- Munje, S. S. , Salunke, P. B. , & Botre, B. S. (2015). Toxicity of newer insecticides against Leucinodes orbonalis (Guen.). Asian Journal of Bio Science, 10, 106–109. [Google Scholar]
- Muñoz Sabater, J. , Dutra, E. , Agustí‐Panaeda, A. , Albergel, C. , Arduini, G. , Balsamo, G. , Boussetta, S. , Choulga, M. , Harrigan, S. , Hersbach, H. , Martens, B. , Miralles, D. G. , Piles, M. , Rodríguez‐Fernández, N. J. , Zsoter, E. , Buontempo, C. , & Thépaut, J. N. (2021). ERA5‐land: A state‐of‐the‐art global reanalysis dataset for land applications. Earth System Data Science, 13, 4349–4383. [Google Scholar]
- Murali, S. , Jalali, S. K. , Shylesha, A. N. , Swamy, T. M. S. , Baskar, R. , & Mruthunjayaswamy, P. (2017). Molecular characterization and their phylogenetic relationship based on mitochondrial cytochrome oxidase I of Brinjal shoot and fruit borer, Leucinodes orbonalis (Guenée) (Lepidoptera: Pyralidae). International Journal of Current Microbiology and Applied Sciences, 6, 2527–2539. 10.20546/ijcmas.2017.607.298 [DOI] [Google Scholar]
- Murthy, P. N. , & Nandihalli, B. S. (2003). Crop loss estimation caused by Leucinodes orbonalis Guenee in potato. Pest Management in Horticultural Ecosystems, 9, 59–62. https://www.scinapse.io/papers/2615855790 [Google Scholar]
- Murugesan, N. , & Murugesh, T. (2009). Bioefficacy of some plant products against brinjal fruit borer, Leucinodes orbonalis Guenee (Lepidoptera: Pyrallidae). Journal of Biopesticides, 2, 60–63. [Google Scholar]
- Muthukumar, M. , & Kalyansundaram, M. (2003). Studies on the efficacy of certain insecticides against Brinjal shoot and fruit borer, Leucinodes orbonalis Guenee. Pestology, 27, 13–16. [Google Scholar]
- Nahar, N. (2020). Co‐designing integrated pest and disease management strategies in eggplant production in Bangladesh. PhD Thesis (p. 164). Wageningen University. [Google Scholar]
- Nahar, N. , Uddin, M. M. , de Jong, P. , Struik, P. C. , & Stomph, T. J. (2020). Technical efficacy and practicability of mass trapping for insect control in Bangladesh. Agronomy for Sustainable Development, 40, 19. [Google Scholar]
- Naik, K. V. , Jalgaonkar, V. N. , Raut, P. P. , Desai, V. S. , & Rajemahadik, V. A. (2017). Studies on some fruit characters of brinjal plants in relation to varying degree of infestation of shoot and fruit borer. Trends in Biosciences, 10, 9392–9400. [Google Scholar]
- Naik, V. C. B. , Rao, P. A. , Krishnayya, P. V. , & Rao, V. S. (2008). Seasonal incidence and management of Leucinodes orbonalis Guence on brinjal. Annals of Plant Protection Sciences, 16, 329–332. [Google Scholar]
- Nair, G. M. , & Nair, M. R. G. K. (1977). Use of some insecticides and their mixtures for control of brinjal pests. Agricultural Research Journal of Kerala, 14, 101–104. [Google Scholar]
- Nair, K. R. (1967). Leucinodes orbonalis Guen. (Lepidoptera:Pyralidae) as a serious pest of potato plants in Mysore state. Indian Journal of Entomology, 29, 96–97. [Google Scholar]
- Najar, I. A. , Khan, A. B. , & Hai, A. (2015). Effect of macrophyte vermicompost on growth and productivity of brinjal (Solanum melongena) under field conditions. international journal of recycling of organic waste in Agriculture, 4, 73–83. [Google Scholar]
- Narendra, S. , Satyakumari, S. , Bhadauria, N. S. , & Bhuva, H. M. (2015). Integrated pest management of shoot and fruit borer (Leucinodes orbonalis G.) of brinjal under semi‐arid condition. Journal of Experimental Zoology, India, 18, 387–392. [Google Scholar]
- Narendran, M. , Lalwani, S. , Girhepuje, P. , Shirale, D. , Mustafa, G. , Gandhi, R. , Parimi, S. , Char, B. R. , & Zehr, U. B. (2006). Production of marker‐free insect resistant transgenic brinjal (Solanum melongena) carrying CRY2AB gene. International Society for Horticultural Science, 752, 535–538. [Google Scholar]
- Nathani, R. K. (1983). Studies on varietal susceptibility and chemical control of brinjal shoot and fruit borer (Leucinodes orbonalis Guenee). Thesis Abstracts, 9, 51–51. [Google Scholar]
- Natikar, P. K. , & Balikai, R. A. (2018a). Survey for insect pests of potato and their natural enemies in Dharwad, Belagavi and Hassan districts of Karnataka during kharif season. Journal of Farm Sciences, 31, 440–446. http://14.139.155.167/test5/index.php/kjas/article/view/9008/9240 [Google Scholar]
- Natikar, P. K. , & Balikai, R. A. (2018b). Status of insect pests of potato and their natural enemies in Karnataka during rabi season. Journal of Experimental Zoology India, 21(2), 1163–1172. [Google Scholar]
- Natikar, P. K. , & Balikai, R. A. (2021). Bio‐efficacy of insecticides against major insect pests of potato during kharif season in India. Potato Research, 65(2022), 379–393. 10.1007/s11540-021-09531-y [DOI] [Google Scholar]
- Natikar, P. K. , & Balikai, R. A. (2022). Bio‐efficacy of insecticides against major insect pests of potato during kharif season in India. Potato Research, 65, 1–15. [Google Scholar]
- Natikar, P. K. , Balikai, R. A. , & Kambrekar, D. N. (2022). Genetic diversity of potato shoot borer Leucinodes orbonalis from major potato‐growing districts of Karnataka, India. Annals of Entomology, 40(1), 50–53. [Google Scholar]
- National Horticulture Board . (2023). Brinjal Post Harvest Technology. https://nhb.gov.in/pdf/vegetable/brinjal/bri008.pdf
- Navasero, M. V. , & Calilung, V. J. (1990). Biology of the eggplant fruit and shoot borer, Leucinodes orbonalis Guenee (Lepidoptera: Pyralidae) in the Philippines. The Philippine Entomologist, 8(2), 749–760. [Google Scholar]
- Navasero, M. V. , & Calumpang, S. M. F. (2013). Influence of intercropping vegetable crops on the population of some arthropods associated with eggplant (Solanum melongena L.) and damage potential of eggplant fruit and shoot borer (Leucinodes orbonalis Guenee) in the presence of radish (Raphanus sativus L.) Philippine agricultural. Scientist, 96, 247–255. [Google Scholar]
- Navasero, M. V. , Candano, R. N. , Hautea, D. M. , Hautea, R. A. , Shotkoski, F. A. , & Shelton, A. M. (2016). Assessing potential impact of Bt eggplants on non‐target arthropods in The Philippines. PLoS One, 11, e0165190. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nawale, R. N. , & Sonone, H. N. (1977). Studies on Resistance in some of the Brinjal Solanum‐Melongena Varieties to the Shoot and Fruit Borer Leucinodes orbonalis . Journal of Maharashtra Agricultural Universities, 2(2), 184–184. [Google Scholar]
- Nayak, U. S. (2014). Influence of Environmental Factors on Population Dynamics and Infestation Pattern of Leucinodes orbonalis in Winter Brinjal of Odisha Vegetos. International Journal of Plant Research, 27, 244–250. [Google Scholar]
- Nayak, U. S. , Baral, K. , Rath, L. K. , & Mondal, P. (2012). Seasonal incidence of Leucinodes orbonalis Guenee in brinjal and the effect of abiotic factors on its population dynamics. Journal of Plant Protection and Environment, 9, 24–27. [Google Scholar]
- Nee, M. (2023). Solanum myriacanthum. Solanaceae Source–A global taxonomic resource for the nightshade family. https://solanaceaesource.myspecies.info/content/solanum‐myriacanthum
- Nee, M. , Vorontsova, M. S. , & Knapp, S. (2023). Solanum viarum. Solanaceae Source–A global taxonomic resource for the nightshade family. https://solanaceaesource.myspecies.info/content/solanum‐viarum
- Naimi, B. , & Araújo, M. B. (2016). SDM: a reproducible and extensible R platform for species distribution modelling. Ecography, 39(4), 368–375. [Google Scholar]
- Niranjana, R. F. , Devi, M. , Shanika, W. , & Sridhar, R. P. (2015). Potential use of egg parasitoids, Trichogramma pretiosum Riley and Trichogramma chilonis Ishii against Brinjal shoot and fruit borer, Leucinodes orbonalis Guenee. Tropical Agricultural Research, 27, 88–94. [Google Scholar]
- Niranjana, V. , & Sridhar, R. P. (2014). Parasitism efficiency of Trichogramma pretiosum on the eggs of brinjal shoot and fruit borer, Leucinodes orbonalis Guenee. Journal of Biological Control, 28, 144–146. [Google Scholar]
- NSW Government . (2009). Tomatoes (fresh) drip irrigation gross margin budget. https://archive.dpi.nsw.gov.au/__data/assets/pdf_file/0003/126732/tomato‐fresh.pdf
- Nusra, M. S. F. , Paranagama, P. A. , Amarasinghe, L. D. , & Udukala, D. N. (2020). Pheromone baited biopesticide for control of Leucinodes orbonalis Guenee in brinjal plant. Frontiers in Bioscience (Elite Edition), 12, 35–47. [DOI] [PubMed] [Google Scholar]
- Nuss, M. , Landry, B. , Mally, R. , Vegliante, F. , Tränkner, A. , Bauer, F. , Hayden, J. , Segerer, A. , Schouten, R. , Li, H. , Trofimova, T. , Solis, M. A. , De Prins, J. , & Speidel, W. (2003. –2023). Global Information System on Pyraloidea. www.pyraloidea.org
- Oerke, E. C. (2006). Crop losses to pests. Journal of Agricultural Science, 144, 31–43. [Google Scholar]
- Padwal, K. G. , Chakravarthy, S. , & Srivastava, C. P. (2022). Genetic variability and population structure of Leucinodes orbonalis (Guenée), a severe insect pest of brinjal in India. Journal of Environmental Biology, 43, 59–65. 10.22438/jeb/43/1/MRN-1868 [DOI] [Google Scholar]
- Pal, J. K. , Singh, M. , Rai, M. , Satpathy, S. , Singh, D. V. , & Kumar, S. (2009). Development and bioassay of Cry1Ac‐transgenic eggplant (Solanum melongena L.) resistant to shoot and fruit borer. Journal of Horticultural Science & Biotechnology, 84, 434–438. [Google Scholar]
- Palraju, M. , Paulchamy, R. , & Sundaram, J. (2018). Population genetic structure and molecular diversity of Leucinodes orbonalis based on mitochondrial COI gene sequences. Mitochondrial DNA Part A DNA Mapping, Sequencing, and Analysis, 29(8), 1231–1239. 10.1080/24701394.2018.1436169 [DOI] [PubMed] [Google Scholar]
- Pandey, S. , Sundria, M. M. , & Ameta, H. K. (2021). Bio‐efficacy of some insecticides against fruit and shoot borer, Leucinodes orbonalis Guenee of brinjal and their effect on natural enemies. Journal of Entomological Research, 45, 95–97. [Google Scholar]
- Paradis, E. , Claude, J. , & Strimmer, K. (2004). APE: Analyses of phylogenetics and evolution in R language. Bioinformatics, 20, 289–290. 10.1093/bioinformatics/btg412 [DOI] [PubMed] [Google Scholar]
- Pareet, J. D. , & Basavanagoud, K. (2009). Evaluation of bio‐pesticides against brinjal shoot and fruit borer and sucking pests. Annals of Plant Protection Sciences, 17, 463–464. [Google Scholar]
- Parvatha RP, 2014. Biointensive integrated pest management in horticultural ecosystems. 1–277.
- Patel, G. A. , & Basu, A. C. (1948). Bionomics of Leucinodes orbanalis Guen. (Lepidoptera) and Epilachna spp. (Coleoptera) the important pest of the brinjal, Solanum melongena L. in Bengal. Proceedings of the Zoological Society of Bengal, 1(1), 117–129. [Google Scholar]
- Patial, A. , & Mehta, P. K. (2006). Record of parasitoid of Leucinodes orbonalis Guenee under mid‐hills of Himachal Pradesh insect. Environment, 12, 138–139. [Google Scholar]
- Patial, A. , Mehta, P. K. , & Sharma, P. C. (2009). Field efficacy of some insecticides and biopesticides against Leucinodes orbonalis Guenee on brinjal. Environment and Ecology, 27, 320–325. [Google Scholar]
- Patnaik, H. P. , Mohapatra, L. N. , & Maity, B. K. (2004). Effectiveness of thiamethoxam 25 WG against the insect pests of brinjal under field conditions. Journal of Plant Protection and Environment, 1, 39–46. [Google Scholar]
- Pawar, D. B. , Kala, P. N. , Ajri, D. S. , & Lawander, K. E. (1987). Chemical control of jassids, aphids and fruit borer of brinjal with synthetic pyrethroids. Journal of Maharashtra Agricultural University, 12, 211–213. [Google Scholar]
- Pawar, S. A. , Datkhile, R. V. , Bhalekar, M. N. , & Bachkar, C. B. (2016). Evaluation of insecticides for management of Leucinodes orbonalis in Brinjal. Annals of Plant Protection Sciences, 24(2), 425–427. [Google Scholar]
- Pawar, S. A. , Patil, C. S. , & Bhalekar, M. N. (2020). Evaluation of ready mix insecticide against sucking pests, shoot and fruit borer in Brinjal. Annals of Plant Protection Sciences, 28, 213–215. [Google Scholar]
- Phytoma (2022). https://www.phytoma.com/sanidad‐vegetal/avisos‐de‐plagas/pimiento‐plagas‐y‐enfermedades‐julio‐2022
- Podder, D. , & Ghosh, S. K. (2019). A new application of Trichoderma asperellum as an anopheline larvicide for eco ‐friendly management in medical science. Scientific Reports, 9, 1108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pollini, A. (2021). La difesa delle piante da orto: Melanzana. Avversità, sintomatologia, provvedimenti. Edagricole.
- Poveda, J. (2021). Trichoderma as biocontrol agent against pests: New uses for a mycoparasite. Biological Control, 15, 104634. [Google Scholar]
- Prabakaran, M. , Sanjayan, K. P. , Amsa, T. , Sithanantham, S. , & Nalini, M. (2019). Enhancing the efficacy of pheromone trap against brinjal shoot and fruit borer. Indian Journal of Entomology, 81, 140–143. [Google Scholar]
- Prasad, C. S. , Abid, H. M. , Pal, R. , & Prasad, M. (2012). Virulence of nematode Heterorhabditis indica (Meerut strain). Against Lepidopteran and Coleopteran Pests Vegetos, 25, 343–351.9. [Google Scholar]
- Prasad, H. , Singh, H. M. , & Singh, A. K. (2005). Effective range of sex pheromone of Leucinodes orbonalis Guen. Journal of Applied Zoological Researches, 16(1), 81–82. [Google Scholar]
- Prasad, R. , & Singh, J. (2012). Efficacy and economics of insecticides against shoot and fruit borer of brinjal. Journal of Eco‐Friendly Agriculture, 7, 60–63. [Google Scholar]
- Prasannakumar, N. R. , Chakravarthy, A. K. , Naveen, A. H. , & Narasimhamurthy, T. N. (2011). Influence of weather parameters on pheromone traps catches of selected lepidopterous insects pests on vegetable crops. Current Biotica, 4, 442–452. [Google Scholar]
- Pray, C. E. , Nagarajan, L. , Huang, J. , Hu, R. , & Ramaswami, B. (2011). Chapter 4 the impact of Bt cotton and the potential impact of biotechnology on other crops in China and India. Frontiers of Economics and Globalization, 10, 83–114. [Google Scholar]
- Priya, R. V. , & Ravi, G. (2020). Validated performance of organic Indian traditional knowledge (ITK) concoctions for the management of brinjal shoot and fruit borer Leucinodes orbonalis Guenee. Insect Environment, 22, 15–25. [Google Scholar]
- Prodhan, M. Z. H. , Hasan, M. T. , Chowdhury, M. M. I. , Alam, M. S. , Rahman, M. L. , Azad, A. K. , Hossain, M. J. , Naranjo, S. E. , & Shelton, A. M. (2019). Bt eggplant (Solanum melongena L.) in Bangladesh: Fruit production and control of eggplant fruit and shoot borer (Leucinodes orbonalis Guenee), effects on non‐target arthropods and economic returns. PLoS One, 13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pugalendhi, L. , Veeraragavathatham, D. , Natarjan, S. , & Praneetha, S. (2010). Utilizing wild relative (Solanum viarum) as resistant source to shoot and fruit borer in brinjal (Solanum melongena Linn.). Electronic journal of. Plant Breeding, 1(4), 643–648. [Google Scholar]
- Puranik, T. R. , Hadapad, A. B. , Salunkhe, G. N. , & Pokharakar, D. S. (2002). Management of shoot and fruit borer, Leucinodes orbonalis Guenee through Bacillus thuringiensis formulations on brinjal. Journal of Entomological Research, 26, 229–232. [Google Scholar]
- Puth, L. M. , & Post, D. M. (2005). Studying invasion: have we missed the boat? Ecology Letters, 8, 715–721. [Google Scholar]
- Puthiamadom, N. , & Joseph, J. (2022). Variability in Solanum species: Morphological characterization. Electronic Journal of Plant Breeding, 13, 137–145. [Google Scholar]
- Quamruzzaman, A. , Islam, F. , Uddin, M. N. , & Chowdhury, M. A. Z. (2019). Performance of green type eggplant in relation of insect and diseases. American Journal of Plant Sciences, 10(1), 139–146. [Google Scholar]
- Qureshi, Q. G. , Mathur, N. M. , & Srivastava, R. C. (1998). Management of brinjal shoot and fruit borer, Leucinodes orbonalis Guenee, using Bacillus thuringiensis var. kurstaki. Advances in IPM for horticultural crops. Proceedings of the First National Symposium on Pest Management in Horticultural Crops: environmental implications and thrusts, Bangalore, India, 15–17 October 1997. pp. 81–83.
- Rabindra, P. , & Janardan, S. (2012). Efficacy and economics of insecticides against shoot and fruit borer of brinjal. Journal of Eco‐friendly Agriculture, 7(1), 60–63. [Google Scholar]
- Radhakrishore, R. K. , Singh, T. K. , & Shah, M. A. S. (2014). Efficacy of certain insecticides against brinjal shoot and fruit borer, Leucinodes orbonalis Guen in Manipur. Journal of Experimental Zoology, India, 17, 551–554. [Google Scholar]
- Raeyat, Z. , Razmjou, J. , Naseri, B. , Ebadollahi, A. , & Krutmuang, P. (2021). Evaluation of the susceptibility of some eggplant cultivars to green peach aphid, Myzus persicae (Sulzer) (Hemiptera: Aphididae). Agriculture (Switzerland), 11, 1–10. [Google Scholar]
- Rahman, M. W. , Das, G. , & Uddin, M. M. (2019). Field efficacy of some new insecticides against brinjal shoot and fruit borer, Leucinodes orbonalis (Guen.) (Lepidoptera: Pyralidae) and their toxic effects on natural enemies. Journal of Bangladesh Agricultural University, 17, 319–324. [Google Scholar]
- Rai, N. P. , Rai, G. K. , Kumar, S. , Kumari, N. , & Singh, M. (2013). Shoot and fruit borer resistant transgenic eggplant (Solanum melongena L.) expressing cry1Aa3 gene: Development and bioassay. Crop Protection, 53, 37–45. [Google Scholar]
- Raja, J. , Rajendran, B. , & Pappiah, C. M. (1998). Management of egg plant shoot and fruit borer Leucinodes orbonalis Guen. Advances in IPM for horticultural crops. Proceedings of the First National Symposium on Pest Management in Horticultural Crops: environmental implications and thrusts, Bangalore, India, 15–17 October 1997, pp. 84–86.
- Rajavel, D. S. , Gopalan, M. , & Balasubramanian, G. (1989). Evaluation of synthetic pyrethroids in the control of brinjal shoot and fruit borer, Leucinodes orbonalis Guen. Madras Agricultural Journal, 76, 114–116. [Google Scholar]
- Rajendra, S. , Anuj, B. , & Singh, D. V. (2015). Efficacy of different doses in combination of chlorantraniliprole and lambda cyhalothrin against pests of brinjal. Annals of Plant Protection Sciences, 23(1), 59–61. [Google Scholar]
- Ramawtar, Y. , Hemant, L. , Sanjay, K. , & Sanp, R. K. (2015). Efficacy of certain botanical insecticides against shoot and fruit borer, Leucinodes orbonalis (Guenee) on brinjal (Solanum melongena L.). The Bioscan, 10(3), 987–990. [Google Scholar]
- Rambaut, A. (2006. –2018). FigTree – Tree Figure Drawing Tool, Version 1.4.4. https://github.com/rambaut/figtree/releases
- Rani, A. T. , Chakravarthy, A. K. , Vasudev, K. , & Prabhakara, M. S. (2017). Sex pheromone trapping plan for the management of brinjal shoot and fruit borer Leucinodes orbonalis Guenee. Indian Journal of Entomology, 79, 440–448. [Google Scholar]
- Ranjith, M. , Nelson, S. J. , Sithanantham, S. , & Hegde, D. R. (2020). Parasitisation of brinjal shoot and fruit borer, Leucinodes orbonalis Guenee by Trathala flavoorbitalis Cameron. Pest Management in Horticultural Ecosystems, 26, 104–108. [Google Scholar]
- Ranjithkumar, L. , Patil, B. V. , Ghante, V. N. , Bheemanna, M. , & Arunkumar, H. (2013). Baseline sensitivity of brinjal shoot and fruit borer, Leucinodes orbonalis (Guenee) in South India to Cry1Ac insecticidal protein of bacillus thuringiensis. Current Science, 105, 366–371. [Google Scholar]
- Rao, V. G. , & Ramana, P. V. (2017). Sex pheromone based integrated pest management strategy for the control of shoot and fruit borer in Brinjal under agency tracts of Rampachodavaram, east Godavari District of AP. Environment and Ecology, 35(4C), 3403–3406. [Google Scholar]
- Re:Fresh Directory . (2018). Published by FPJ, Peterborough (p. 372). [Google Scholar]
- Rider, D. A. (2015). Plant Host Records, Plataspidae. https://www.ndsu.edu/faculty/rider/Pentatomoidea/Hosts/plant_Plataspididae.htm
- Robinet, C. , Roques, A. , Pan, H. , Fang, G. , Ye, J. , Zhang, Y. , & Sun, J. (2009). Role of human‐mediated dispersal in the spread of the pinewood nematode in China. PLoS One, 4, e4646. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Robinson, G. S. , Ackery, P. R. , Kitching, I. , Beccaloni, G. W. , & Hernández, L. M. (2023). HOSTS (from HOSTS–a database of the World's lepidopteran hostplants) [Data Set Resource]. Natural History Museum. https://data.nhm.ac.uk/dataset/hosts/resource/877f387a‐36a3‐486c‐a0c1‐b8d5fb69f85a [Google Scholar]
- Rohokale, Y. A. , Sonkamble, M. M. , Bokan, S. C. , & Bhede, B. V. (2018). Efficacy of newer insecticide combinations against brinjal shoot and fruit borer. International Journal of Entomology Research, 3, 36–39. [Google Scholar]
- Rossi, E. , De Santis, M. , Stancanelli, G. , & Maiorano, A. (2023). EFSA climate suitability analysis of Leucinodes orbonalis . Zenodo. 10.5281/zenodo.8388938 [DOI] [Google Scholar]
- Roy, G. , Gazmer, R. , Sarkar, S. , Laskar, N. , Das, G. , & Samanta, A. (2016). Comparative bioefficacy of different insecticides against fruit and shoot borer (Leucinodes orbonalis Guenee) of brinjal and their effect on natural enemies. International Journal of Green Pharmacy, 10, S257–S260. [Google Scholar]
- Roy, N. C. , & Panda, B. N. (1973). A note on control of BrinjaI fruit and shoot borer (Leucinodes orbonalis) with insecticide application. Science and Culture, 39. [Google Scholar]
- Roy, P. , Shome, M. G. , & Dasgupta, A. (1973). Studies on the control of brinjal shoot and fruit borer, Leucinodes orbonalis Guen. (Pyralidae, Lepidoptera). Science and Culture, 39, 270–272. [Google Scholar]
- Rubel, F. , Brugger, K. , Haslinger, K. , & Auer, I. (2017). The climate of the European Alps: Shift of very high resolution Köppen‐Geiger climate zones 1800–2100. Meteorologische Zeitschrift, 26, 115–125. [Google Scholar]
- Sagarbarria, M. G. S. , Marasigan, J. C. M. , Taylo, L. D. , & Hautea, D. M. (2018). Identity and genetic structure of eggplant fruit and shoot borer, Leucinodes orbonalis Guenée (Lepidoptera:Crambidae) populations in The Philippines inferred from morphological traits and COI sequence data. Journal of Asia‐Pacific Entomology, 21, 1009–1019. 10.1016/j.aspen.2018.07.012 [DOI] [Google Scholar]
- Sahoo, S. K. , Mandal, K. , Kaur, R. , Battu, R. S. , & Singh, B. (2013). Persistence of thiacloprid residues on brinjal (Solanum melongena L.). Environmental Monitoring and Assessment, 185, 7935–7943. [DOI] [PubMed] [Google Scholar]
- Sahu, R. , Kumar, A. , Khan, H. H. , & Sahu, P. S. (2017). Seasonal incidence of brinjal shoot and fruit borer, Leucinodes orbonalis Guen. Under field conditions. Journal of Experimental Zoology India, 20, 709–710. [Google Scholar]
- Saini, D. K. , & Kaushik, P. (2019). Visiting eggplant from a biotechnological perspective: A review. Scientia Horticulturae, 253, 327–340. [Google Scholar]
- Samanta, A. , Roy, P. , Das, A. K. , Majumdar, D. , & Somchoudhury, A. K. (1999). Bio‐efficacy of a new formulation of quinalphos against insect pests of brinjal. Journal of Interacademicia, 3, 49–52. [Google Scholar]
- Sandanayake, W. R. M. (1992). Instar determination and larval distribution in brinjal shoot and fruit borer, Leucinoides orbonalis (Guen).
- Sandip, P. , Sarkar, S. , Alam, S. K. F. , Das, B. C. , Dhar, P. P. , & Samanta, A. (2018). Field efficacy of flubendiamide 480SC against brinjal shoot and fruit borer (Leucinodes orbonalis). Indian Journal of Entomology, 80, 432–434. [Google Scholar]
- Sandip, P. , Thakur, N. S. A. , & Firake, D. M. (2016). Evaluation of bio‐pesticides and insecticides against brinjal shoot and fruit borer (Leucinodes orbonalis guenee) in Meghalaya of north‐eastern India. International Journal of Bio‐Resource and Stress Management, 7, 1032–1036. [Google Scholar]
- Sanjeevi, K. A. , & Muthukrishnan, N. (2017). In‐vivo and field evaluation of spinetoram 12 SC against Lampides boeticus on pigeon pea. Legume Research, 40, 1126–1132. [Google Scholar]
- Sankaranarayanan C and Askary TH, 2017. Status of entomopathogenic nematodes in integrated pest management strategies in India. 362–382.
- Sarangdevot, S. S. , Kumar, S. , Naruka, P. S. , & Panchauri, C. P. (2010). Studies of bio‐efficacy of some insecticides against shoot and fruit borer Leucinodes orbonalis of brinjal Solanum melongena Linn. Pestology, 34, 40–44. [Google Scholar]
- Sardar, S. , & Guram, M. S. (1967). Trials for the control of brinjal fruit and shoot borer. Plant Protection Bulletin, 18, 13–17. [Google Scholar]
- Sarkar, S. , Pal, S. , Sahoo, S. , Laskar, N. , & Ghosh, J. (2022). Field efficacy study of different biorationals and insecticides against brinjal shoot and fruit borer (Leucinodes orbonalis Guenee) under terai region of West Bengal. Uttar Pradesh Journal of Zoology, 43, 57–66. [Google Scholar]
- Särkinen, T. , Poczai, P. , Barboza, G. E. , van der Werden, G. M. , Baden, M. , & Knapp, S. (2023). Solanum nigrum. Solanaceae Source–A global taxonomic resource for the nightshade family. https://solanaceaesource.myspecies.info/content/solanum‐nigrum [Google Scholar]
- Sarnabati, L. , & Ray, D. C. (2017). Efficacy of newer insecticides against brinjal shoot and fruit borer Leucinodes orbonalis Guen. In Manipur. Indian Journal of Entomology, 79, 55–58. [Google Scholar]
- Satpathy, S. , & Mishra, D. S. (2011). Use of intercrops and antifeedants for management of eggplant shoot and fruit borer Leucinodes orbonalis (Lepidoptera: Pyralidae). International Journal of Tropical Insect Science, 31, 52–58. [Google Scholar]
- Satpathy, S. , Tripathi, U. K. , Srivastava, D. , & Rai, S. (2006). Field evaluations of chemical insecticides, a growth regulator and a bio‐insecticide against eggplant shoot and fruit borer on eggplant. Tests of Agrochemicals and Cultivars, 5–6. [Google Scholar]
- Satpute BB, Mote UN and Shivu B, 2002. Efficacy of spark, Bacillus thuringiensis and Azadirachtin in Different Sequences against Pest Complex of Brinjal. 30–32.
- Saxena, P. N. (1965). The life history and biology of Leucinodes orbonalis Quen, (Lepidoptera: Pysaustidae) [sic]. Journal of the Zoological Society of India, 17, 64–70. [Google Scholar]
- Scott, G. J. , & Suarez, V. (2011). Growth rates for potato in India and their implications for industry. Potato Journal, 38(2), 100–112. [Google Scholar]
- Sharma, I. D. , Nargeta, D. S. , & Nath, A. (2004). Management of brinjal shoot and fruit borer (Leucinodes orbonalis Guen.) and persistence of beta‐cyfluthrin in brinjal fruits. Pesticide Research Journal, 16, 29–32. [Google Scholar]
- Sharma, S. , Sharma, A. , & Katoch, V. (2020). Biotechnological interventions in eggplant (Solanum melongena L.). Journal of Horticultural Science and Biotechnology, 95, 273–285. [Google Scholar]
- Sharmin, A. , Alam, M. Z. , Rahman, M. M. , & Akanda, A. M. (2017). Evaluation of some management options against brinjal (Solanum melongena L.) shoot and fruit borer (Leucinodes orbonalis Guenee). The Agriculturists, 15, 49–57. [Google Scholar]
- Shashank, P. R. , Ohja, R. , Venkatesan, T. , Jalali, S. K. , & Bhanu, K. R. M. (2015). Molecular characterization of brinjal shoot and fruit borer, Leucinodes orbonalis (Guenée) (Lepidoptera: Crambidae) based on mitochondrial marker cytochrome oxidase I and their phylogenetic relationship. Indian Journal of Experimental Biology, 52, 51–55. [PubMed] [Google Scholar]
- Shelton, A. M. (2010). The long road to commercialization of Bt brinjal (eggplant) in India. Crop Protection, 29, 412–414. [Google Scholar]
- Shelton, A. M. (2021). Bt eggplant: A personal account of using biotechnology to improve the lives of resource‐poor farmers. American Entomologist, 67, 52–59. [Google Scholar]
- Shelton, A. M. , Hossain, M. J. , Paranjape, V. , Azad, A. K. , Rahman, M. L. , Khan, A. S. M. M. R. , Prodhan, M. Z. H. , Rashid, M. A. , Majumder, R. , Hossain, M. A. , Hussain, S. S. , Huesing, J. E. , & McCandless, L. (2018). Bt eggplant project in Bangladesh: History, present status, and future direction. Frontiers in Bioengineering and Biotechnology, 6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shelton, A. M. , Hossain, M. J. , Paranjape, V. , Azad, A. K. , Rahman, M. L. , Khan, A. S. M. M. R. , Prodhan, M. Z. H. , Rashid, M. A. , Majumder, R. , Hossain, M. A. , Hussain, S. S. , Huesing, J. E. , & McCandless, L. (2019). Bt Brinjal in Bangladesh: The first genetically engineered food crop in a developing country cold Spring Harbor. Perspectives in Biology, 11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shende, V. D. , Chattopadhyay, A. , Yadav, Y. , Seth, T. , & Mukherjee, S. (2016). Breeding round fruited brinjal for export from eastern India. Agricultural Research, 5(3), 219–229. [Google Scholar]
- Shinde, K. G. (2007). Gene action of fruit yield and its components in brinjal (Solanum melongena L.). Advances in Plant Sciences, 20, 63. [Google Scholar]
- Shirale, D. , Patil, M. , & Parimi, S. (2017). Insecticide resistance in field populations of Leucinodes orbonalis (Lepidoptera: Crambidae) in India. The Canadian Entomologist, 149, 399–407. 10.4039/tce.2017.3 [DOI] [Google Scholar]
- Shrestha, B. , & Lee, Y. (2020). Cellular and molecular mechanisms of DEET toxicity and disease‐carrying insect vectors: A review. Genes and Genomics, 42, 1131–1144. [DOI] [PubMed] [Google Scholar]
- Shrivastava, D. , Dalal, M. , Nain, V. , Sharma, P. C. , & Kumar, P. A. (2011). Targeted integration of Bacillus Thuringiensis δ‐endotoxin cry1Fa1 in Brinjal (Solanum melongena L.). Current Trends in Biotechnology and Pharmacy, 5, 1149–1156. [Google Scholar]
- Shukla, R. P. (1986). Spatial pattern of egg plant shoot and fruit borer, Leucinodes orbonalis Guen (Lepidoptera, Pyraustidae) larvae. Insect Science and its Application, 7(5), 611–615. [Google Scholar]
- Shukla, S. , Singh, D. , Bairwa, B. , & Jat, B. L. (2017). Study on biophysical and biochemical basis of shoot and fruit borer tolerance in brinjal. International Journal of Plant Protection, 10, 206–228. [Google Scholar]
- Sidhu, A. S. & Dhatt, A. S . (2017). Current status of brinjal research in India. pp. 243–247.
- Silva Dias, J. , & Ryder, E. J. (2011). World vegetable industry: Production, breeding, Trends. Horticultural Reviews, 38, 299–356. [Google Scholar]
- Simberloff, D. (2009). The role of propagule pressure in biological invasions. Annual Review of Ecology, Evolution, and Systematics, 40, 81–102. [Google Scholar]
- Singh, B. K. , Pandey, R. , Singh, A. K. , & Dwivedi, S. V. (2021). Efficacy of certain insecticides against brinjal shoot and fruit borer Leucinodes orbonalis Guenee. Indian Journal of Entomology, 83, 464–467. [Google Scholar]
- Singh, D. K. , Pal, S. , Singh, D. N. , & Pandey, V. (2014). Non‐chemical practices against important pests of Brinjal, Solanum Melongena L. Annals of Agricultural Biological Research, 19, 97–101. [Google Scholar]
- Singh, D. K. , Singh, R. , Datta, S. D. , & Singh, S. K. (2009). Seasonal incidence and insecticidal management of shoot and fruit borer (Leucinodes orbonalis Guenee) in brinjal. Annals of Horticulture, 2, 187–190. [Google Scholar]
- Singh, H. S. , Krishna Kumar, N. K. , & Krishnakumari, B. (2007). Effect of pheromone lure‐distance and direction in trapping brinjal fruit and shoot borer (Leucinodes orbonalis Guen.) male moths. Journal of Horticultural Science, 2, 67–70. [Google Scholar]
- Singh, J. (2018a). Role of earthworm in sustainable agriculture. Sustainable Food Systems from Agriculture to Industry, Academic Press, 83–122. 10.1016/B978-0-12-811935-8.00003-2. [DOI] [Google Scholar]
- Singh, M. P. (2018b). Field efficacy of insecticides against brinjal shoot and fruit borer in rainfed farming situations of Uttarakhand hills. Journal of Hill Agriculture, 9, 104–108. [Google Scholar]
- Singh, M. P. , & Sinha, M. M. (1964). Control schedule for shoot and fruit borer of brinjal (Leucinodes orbonalis Guen.). Proceedings of the Bihar Academy of Agricultural Sciences, 12–13, 26–30. [Google Scholar]
- Singh, R. , Bhatnagar, A. , & Singh, D. V. (2015). Efficacy of different doses in combination of chlorantraniliprole and lambda cyhalothrin against pests of brinjal. Annals of Plant Protection Sciences, 23, 59–61. [Google Scholar]
- Singh, R. K. R. , Singh, T. K. , & Shah, M. A. S. (2015). Field efficacy of insecticides against brinjal shoot and fruit borer. Indian Journal of Entomology, 77, 313–316. [Google Scholar]
- Singh, S. (2019). Efficacy and economics of pest management modules against Brinjal shoot and fruit borer (Leucinodes orbonalis). Annals of Plant Protection Sciences, 27, 320–323. [Google Scholar]
- Singh, S. P. N. , & Kumar, N. (2011). Insecticidal evaluation against Leucinodes orbonalis Guen. On Brinjal. Pesticide Research Journal, 23, 227–229. [Google Scholar]
- Singh, S. S. , & Dwivedi, S. K. (2020). Evaluation of different newer chemical pesticides through on farm testing (OFTs) against major pests and diseases of vegetable crops. Progressive Horticulture, 52, 193–198. [Google Scholar]
- Singh, Y. P. , & Singh, P. P. (2002). Natural parasites and extent of parasitism to shoot and fruit borer (Leucinodes orbonalis) of brinjal (Solanum melongena) at medium high‐altitude hills of Meghalaya. Indian Journal of Entomology, 64, 222–226. [Google Scholar]
- Singla, P. , Bhullar, M. B. , & Kaur, P. (2018). Biological studies on brinjal shoot and fruit borer, Leucinodes orbonalis Guenee. Journal of Entomology and Zoology Studies, 6(1), 161–165. [Google Scholar]
- Sinha, K. K. , Chakraborty, D. P. , Dasgupta, K. P. , & Dhua, S. P. (1976). Studies on the NPK and insecticide interaction in respect of pest infestation and yield in brinjal fertilizer technology. India, 13, 15–18. [Google Scholar]
- Sinha, S. , & Gopal, M. (2002). Evaluating the safety of β‐cyfluthrin insecticide for usage in eggplant (Solanum melongena L.) crop. Bulletin of Environmental Contamination and Toxicology, 68, 400–405. [DOI] [PubMed] [Google Scholar]
- Sinha, S. R. , & Nath, V. (2011). Management of insect pests through insecticides and mixture in brinjal. Annals of Plant Protection Sciences, 19, 318–320. [Google Scholar]
- Sinha, S. R. , & Vishwa, N. (2012). Evaluation of new insecticides against insect pests of brinjal. Annals of Plant Protection Sciences, 20, 287–289. [Google Scholar]
- Sivakumar, R. , Nachiappan, R. M. , & Selvanarayanan, V. (2003). Field efficacy of profenofos against shoot and fruit borer, Leucinodes orbonalis (Guenee) of egg plant. Pestology, 27, 23–25. [Google Scholar]
- Sivapragasam, A. , & Chua, T. H. (1997). Natural enemies for the cabbage webworm, Hellula undalis (Fabr.) (Lepidoptera: Pyralidae) in Malaysia. Researches on Population Ecology, 39, 3–10. [Google Scholar]
- Sofiya, M. , & Raj, R. N. (2021). G × E interaction, biochemical characterization and SSR marker analysis of eggplant (Solanum melongena) for shoot and fruit borer resistance. Research on Crops, 22, 608–615. [Google Scholar]
- Sohi, G. S. (1944). Insect pests of vegetables in the Punjab and their control. Punjab Fruit Journal, 8, 87–94. [Google Scholar]
- Solis, M. A. (2006). Key to selected Pyraloidea (Lepidoptera) larvae intercepted at U. S. ports of entry: Revision of Pyraloidea in Keys to some frequently intercepted lepidopterous larvae by Weiman 1986. USDA systematic entomology The Laboratory, 1. https://digitalcommons.unl.edu/systentomologyusda/1 [Google Scholar]
- Sorensen, K. A. , Mohankumar, S. , & Thangaraj, S. R. (2016). Physical, mechanical and cultural control of vegetable insects. In Muniappan R. & Heinrichs E. Z. (Eds.), Integrated pest management of tropical vegetable crops (pp. 131, 131–148, 148). Springer. [Google Scholar]
- Srinivasan, R. (2008). Integrated Pest management for eggplant fruit and shoot borer (Leucinodes orbonalis) in south and southeast Asia: Past, present and future. Journal of Biopesticides, 1, 105–112. 10.57182/jbiopestic.1.2.105-112 [DOI] [Google Scholar]
- Srinivasan, R. , & Huang, C. C. (2009). The effect of simulated borer infested shoot pruning on yield parameters of eggplant. Journal of Asia‐Pacific Entomology, 12, 41–43. [Google Scholar]
- Srinivasappa, K. N. , Gowda, P. H. R. , Thimmaiah, S. K. , & Mahadevu, P. (1998). Studies on growth, yield and shoot and fruit borer (Leucinodes orbonalis Guenee.) resistance in M3 progenies of Solanum macrocarpon L. Journal of Nuclear Agriculture and Biology, 27, 65–68. [Google Scholar]
- Sugandha, S. , & Sharma, P. C. (2017). Relative toxicity of novel insecticides and biopesticides against Leucinodes orbonalis Guenee (Lepidoptera: Pyralidae). Journal of Insect Science (Ludhiana), 30(2), 176–180. [Google Scholar]
- Sujayanand, G. K. , Sharma, R. K. , Shankarganesh, K. , Supradip, S. , & Tomar, R. S. (2015). Crop diversification for sustainable insect pest management in eggplant (Solanales: Solanaceae). Florida Entomologist, 98, 305–314. [Google Scholar]
- Sultana, H. , Mannan, M. A. , Kamal, M. M. , Quddus, K. G. , & Das, S. (2018). Appraisal of resistant genotypes against brinjal shoot and fruit borer (BSFB), Leucinodes orbonalis, Guenee. Journal of the Bangladesh Agricultural University, 16, 227–231. [Google Scholar]
- Suresh, K. , Rani, B. U. , Rajendran, R. , & Baskaran, M. R. K. (2007). Management of brinjal shoot and fruit borer through nutritional manipulation. Journal of Entomological Research (New Delhi), 31, 191–196. [Google Scholar]
- Talukder, P. , Dutta, D. , Ghosh, E. , Bose, I. , & Bhattacharjee, S. (2021). Role of plant secondary metabolites in combating pest induced stress in brinjal (Solanum melongena L.). Journal of Environmental Engineering and Landscape Management, 29, 449–453. [Google Scholar]
- Taylo, L. D. , Sison, M. L. J. , & Hautea, D. M. (2016). Use of artificial infestation for field bioefficacy assessment of Bt eggplant against the eggplant fruit and shoot borer, Leucinodes orbonalis Guenee (Lepidoptera: Crambidae) Philippine. Agricultural Scientist, 99, 119–126. [Google Scholar]
- Tejavathu, H. S. , Kalda, T. S. , & Gupta, S. S. (1991). Note on relative resistance to shoot and fruit borer in eggplant. Journal of Horticulture, 48, 356–359. [Google Scholar]
- Thomson, J. A. (2018). The pros and cons of GM crops. Functional Plant Biology, 45, 297–304. [DOI] [PubMed] [Google Scholar]
- Tribikram, S. , & Patnaik, H. P. (2010). Patterns of larval incidence of shoot and fruit borer, Leucinodes orbonalis Guenee in eggplant ecosystem of coastal Orissa. Current Biotica, 4(2), 249–255. [Google Scholar]
- Tripathi, M. K. (2016). Use of chemical and biopesticides against Leucinodes orbonalis and yield in Brinjal. Annals of Plant Protection Sciences, 24, 429–430. [Google Scholar]
- Tripathi, S. R. , & Singh, A. K. (1991). Some observations on population dynamics of brinjal borer, Leucinodes orbonalis (Guen.) (Lepidoptera: Pyralidae). Annals of Entomology (Dehra Dun), 9(1), 15–24. [Google Scholar]
- Tripathy, P. , & Patnaik, H. P. (2003). On farm evaluation of insecticidal schedules against the shoot and fruit borer (Leucinodes orbonalis Guen) infestation in brinjal. Orissa Journal of Horticulture, 31, 107–109. [Google Scholar]
- Udayagiri, S. , & Mohan, N. J. (1985). New records of insect pests associated with the medicinal plant Solanum viarum . Indian Drugs, 23, 1–4. [Google Scholar]
- Ullah, M. , Ullah, F. , Khan, M. A. , Ahmad, S. , Jamil, M. , Sardar, S. , Tariq, K. , & Ahmed, N. (2022). Efficacy of various natural plant extracts and the synthetic insecticide cypermethrin 25EC against Leucinodes orbonalis and their impact on natural enemies in brinjal crop. International Journal of Tropical Insect Science, 42, 173–182. [Google Scholar]
- Valdés, B. (2012). Solanaceae. In: Euro+Med Plantbase – the information resource for Euro‐mediterranean plant diversity. https://ww2.bgbm.org/EuroPlusMed/PTaxonDetail.asp?NameCache=Solanaceae&PTRefFk=7100000
- Van Vang, L. , Yan, Q. , Nghia, N. T. N. , Khanh, C. N. Q. , & Ando, T. (2018). Unsaturated Cuticular hydrocarbon components of the sex pheromone of eggplant fruit borer, Leucinodes orbonalis Guenée (Lepidoptera:Crambidae). Journal of Chemical Ecology, 44, 631–636. [DOI] [PubMed] [Google Scholar]
- van der Gaag, D. J. , Holt, J. Leach, A. W. , & Loomas, A. J. M. (2019). Model of the probability of pest transfer to a site suitable for establishment following their arrival on imported fruit, cut‐flower or vegetable produce. Crop Protection, 117, 135–146. 10.1016/j.cropro.2018.11.016 [DOI] [Google Scholar]
- Veeravel, R. , & Baskaram, P. (1977). Performance of certain insecticides in the control of insect pests of brinjal (Solanum melongena L.). Pesticides, 11, 31–32. [Google Scholar]
- Venette, R. C. (2017). Climate analyses to assess risks from invasive forest insects: Simple matching to advanced models. Current Forestry Reports, 3, 255–268. https://www.fs.usda.gov/nrs/pubs/jrnl/2017/nrs_2017_venette_001.pdf [Google Scholar]
- Venkatesan, T. , Jalali, S. K. , & Srinivasamurthy, K. (2009). Competitive interactions between Goniozus nephantidis and Bracon brevicornis, parasitoids of the coconut pest Opisina arenosella . International Journal of Pest Management, 55, 257–263. [Google Scholar]
- Venkatraman, T. V. , & Chander, R. (1967). Experiments on the possible use of Bacillus thuringiensis Berliner in the control of crop pests–II. Susceptibility of some lepidopterous pests to B. thuringiensis Proceedings of the Indian Academy of Sciences–Section B, 66, 231–236. [Google Scholar]
- Verma, T. S. , & Lal, O. P. (1985). A new record of Itamoplex sp. (Hymenoptera: Inchneumonidae) parasitizing eggplant shoot and fruit borer in Kulu Valley, Himachal Pradesh. Bulletin of Entomology, 26, 219–222. [Google Scholar]
- Vijayasree, V. , & Bai, H. (2019). Efficacy and biosafety of new generation insecticides for the management of Leucinodes orbonalis Guenee (Lepidoptera: Pyralidae) in brinjal and Earias vitella Fabricius (Lepidoptera: Noctuidae) in okra. Entomon, 44, 203–212. [Google Scholar]
- Vinod, S. , Ramesh, L. , & Anil, C. (2001). Screening of brinjal (Solanum spp.) germplasm against shoot and fruit borer, Leucinodes orbonalis Guen. Insect Environment, 7(3), 126–127. [Google Scholar]
- Vinothkumar, B. (2021). Field evaluation of Carbosulfan 25EC against Brinjal shoot and fruit borer Leucinodes orbonalis Guenee. Indian Journal of Entomology, 83(2), 207–210 [Google Scholar]
- Visalakshy, P. N. , Ganga, K. A. , & Hussaini, S. S. (2009). Field efficacy of the entomopathogenic nematode Steinernema carpocapsae (Weiser, 1955) against brinjal shoot and fruit borer, Leucinodes orbonalis Guenee. Nematologia Mediterranea, 37, 133–137. [Google Scholar]
- Visnupriya, M. , & Muthukrishnan, N. (2019). In vivo and field evaluation of newer green insecticide spinetoram 12 SC against shoot and fruit borer, Leucinodes orbonalis Guenee on brinjal. Crop Research (Hisar), 54(1‐2), 46–52. [Google Scholar]
- Vorontsova, M. S. (2023). Solanum aethiopicum. Solanaceae Source–A global taxonomic resource for the nightshade family. https://solanaceaesource.myspecies.info/content/solanum‐aethiopicum
- Vorontsova, M. S. , & Knapp, S. (2023a). Solanum torvum. Solanaceae Source–A global taxonomic resource for the nightshade family. https://solanaceaesource.myspecies.info/content/solanum‐torvum
- Vorontsova, M. S. , & Knapp, S. (2023b). Solanum anomalum. Solanaceae Source–A global taxonomic resource for the nightshade family. https://solanaceaesource.myspecies.info/content/solanum‐anomalum
- Walunj, A. R. , Mote, U. N. , Desai, A. C. , & Parikh, K. M. (1996). Efficacy of ZA −199, a neem‐based insecticide against brinjal shoot and fruit borer. Pestology, 20, 7–9. [Google Scholar]
- Wankhede, S. M. , & Kale, V. D. (2010a). Effect of insecticides on brinjal shoot and fruit borer, Leucinodes orbonalis Guenee. Annals of Plant Protection Sciences, 18, 336–339. [Google Scholar]
- Wankhede, S. M. , & Kale, V. D. (2010b). Performance of some insecticides against Leucinodes orbonalis G. International Journal of Plant Protection, 3, 257–259. [Google Scholar]
- Witzgall, P. , Kirsch, P. , & Cork, A. (2010). Sex pheromones and their impact on Pest management. Journal of Chemical Ecology, 36, 80–100. [DOI] [PubMed] [Google Scholar]
- Wu, H. , Ju, Z. , Li, Y. , Liu, C. , Huang, J. , Tang, Y. , Huang, M. , Yu, Y. , Yang, G. , & Li, H. (2013). New occurrence and green control techniques of eggplant fruit and shoot borers in Shanghai. China Plant Protection, 33(12), 30–32. [Google Scholar]
- Xiao, Y. , & Wu, K. (2019). Recent progress on the interaction between insects and Bacillus thuringiensis crops (p. 374). Philosophical Transactions of the Royal Society B, Biological Sciences. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yadav, R. , Lyall, H. , Kumar, S. , & Sanp, R. K. (2015). Efficacy of certain botanical insecticides against shoot and fruit borer, Leucinodes orbonalis (Guenee) on brinjal (Solanum melongena L.) the. Bioscan, 10, 987–990. [Google Scholar]
- Yasodha, P. , & Natarajan, N. (2009). Record of parasitoids and pathogens on Leucinodes orbonalis . Madras Agricultural Journal, 96, 194–199. [Google Scholar]
- Yasodha, P. , Natarajan, N. , & Udayasuriyan, V. (2007). Record of entomopathogens on Leucinodes orbonalis Guenee (Pyraustidae: Lepidoptera). Indian Journal of Entomology, 69, 373–380. [Google Scholar]
- Yazdani, S. S. , Mehto, D. N. , Singh, R. , & Kumar, A. (1981). Control of brinjal shoot and fruit borer Leucinodes orbonalis with granules alone and in combination with spray formulations. Indian Journal of Entomology, 43, 297–301. [Google Scholar]
- Ye, B. , Wang, F. , Deng, J. , & WangYulei, Y. (2015). Identification and field application of sex pheromone componentsfrom eggplant fruit and shoot borers and factors affecting the trapping efficiencyof sex attractants. Journal of Environmental Entomology. [Google Scholar]
- Yengkokpam, P. , & Mazumder, P. B. (2021). Antioxidant enzymatic activities and profiling of gene expression associated with organophosphate stress tolerance in Solanum melongena L. cv. Longai. 3. Biotech, 11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yousafi, Q. , Afzal, M. , & Aslam, M. (2015). Management of brinjal shoot and fruit borer, Leucinodes orbonalis Guenee, with selected insecticides Pakistan. Journal of Zoology, 47. [Google Scholar]
- Yousafi, Q. , Afzal, M. , Aslam, M. , & Abid, A. D. (2016a). Effectiveness and benefit cost ratio of selected insecticides at different application on intervals for brinjal shoot and fruit borer, Leucinodes orbonalis (G.) management on brinjal, Solanum melongena (L.) at Sahiwal, Pakistan. Phytoparasitica, 44, 423–427. [Google Scholar]
- Yousafi, Q. , Afzal, M. , Aslam, M. , & Abid, A. D. (2016b). Monitoring of brinjal shoot and fruit borer (Leucinodes orbonalis Guenee) adult populations in brinjal (Solanum melongena L.) using light traps. Pakistan Journal of Zoology, 48, 333–343. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supplementary material to the 'Pest risk assessment of Leucinodes orbonalis for the European Union' ‐ Entry model