Figure 7. Natural variation in C. elegans egg-laying in response to neuromodulatory agents.
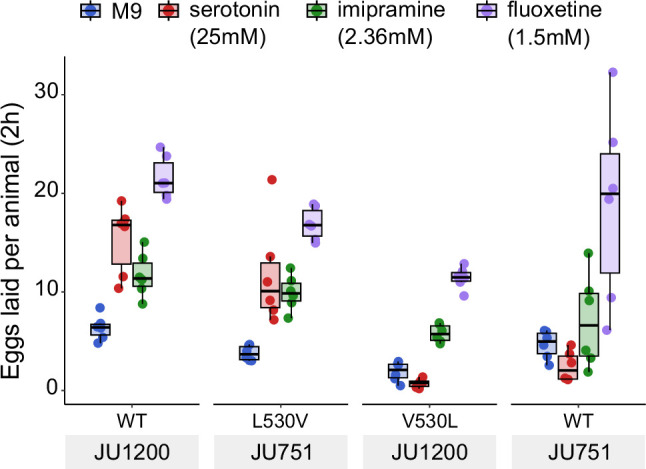
(A) Cartoon of the neural circuit controlling C. elegans egg laying (Collins et al., 2016; Kopchock et al., 2021). The structure of the C. elegans egg-laying circuit is simple, containing two classes of motoneurons, the two serotonergic hermaphrodite-specific motoneurons (HSN) and the six ventral cord motoneurons (VC), which provide synaptic input to the egg-laying muscles (VM). Serotonin from HSN act through vulval muscle receptors to increase muscle excitability, which together with rhythmic signals from motor neurons, causes contractions of VM during egg laying (Collins et al., 2016; Kopchock et al., 2021). Mechanical feedback in response to egg accumulation favours exit from the inactive state (Collins et al., 2016; Medrano and Collins, 2023). Muscles are indicated by rectangles, neurons by circles, and neurosecretory cells (uv1) by triangles. Principal neurotransmitters released by neurons are indicated next to neurons (ACh: Acetylcholine). (B) Natural variation in egg-laying activity in response to exogenous serotonin, fluoxetine, and imipramine. Adult hermaphrodites (mid-L4 +30 hr) were placed in M9 buffer without food (control) or in M9 containing the indicated concentrations of serotonin, fluoxetine, and imipramine. The number of eggs laid were scored after two hours. Assays for each of the three treatments were carried out independently; the 15 strains were scored in parallel in both control and treatment conditions for each of the three assays. Serotonin: for each strain, 11–24 replicates (each containing 3.73±0.36 individuals on average) were scored for serotonin and control (M9 buffer) conditions. Align Rank Transform ANOVA, fixed effect Treatment: F1,390=432.62, p<0.0001, fixed effect Strain: F14,390=42.94, p<0.0001; interaction Treatment x Strain: F14,390=34.40, p<0.0001. Serotonin stimulated egg-laying in Class, II and III A strains but had no effect on egg laying in Class III B and JU2829 (Class III A) (Tukey’s honestly significant difference, ***p<0.0001; ns: not significant). Imipramine: for each strain, 6–18 replicates/wells (each containing 4.62±0.50 individuals on average) were scored for serotonin and control (M9 buffer) conditions. Align Rank Transform ANOVA, fixed effect Treatment: F1,222=562, p<0.0001, fixed effect Strain: F14,222=23.86, p<0.0001; interaction Treatment x Strain: F14,222=8.52, p<0.0001. Imipramine stimulated egg laying in strains from the 4 Class (Tukey’s honestly significant difference, ***p<0.0001; ns: not significant). Fluoxetine: for each strain, 12–24 replicates/wells (each containing 3.31±0.37 individuals on average) were scored for serotonin and control (M9) conditions. Align Rank Transform ANOVA, fixed effect Treatment: F1,378=1005, p<0.0001, fixed effect Strain: F14,378=30.09, p<0.0001; interaction Treatment x Strain: F14,378=16.35, p<0.0001. Imipramine stimulated egg-laying in strains from the 4 Class (Tukey’s honestly significant difference, ***p<0.0001; ns: not significant). For detailed statistical results, see Figure 7—source data 2. (C) Effects of exogenous serotonin (25 mM) on egg laying activity in strains with strongly divergent egg retention due to variation in a single amino acid residue of KCNL-1. Strains JU1200WT (canonical egg retention), JU751KCNL-1 L530V (CRISPR-Cas9-engineered, weak egg retention), JU1200KCNL-1 V530L (CRISPR-Cas9-engineered, strong egg retention) and JU751WT (strong egg retention). Adult hermaphrodites (mid-L4 +30 hr) were placed into M9 buffer without food (control) or M9 with serotonin (25 mM). Serotonin stimulated egg-laying in JU751KCNL-1 L530V and JU1200WT but inhibited egg laying in JU751WT and JU1200KCNL-1 V530L (Kruskal-Wallis Tests were performed separately for each strain to test for the effect of serotonin on the number of eggs laid; *p<0.05). For each strain, six replicates (each containing 5.50±0.92 individuals on average) were scored for serotonin and control conditions. For additional data, see Figure 7—figure supplement 1.

Figure 7—figure supplement 1. Effects of exogenous serotonin, Imipramine and Fluoxetine on egg laying activity in strains with strongly divergent egg retention due to variation in a single amino acid residue of KCNL-1.