

Abstract
Hepatocellular carcinoma (HCC) is one of the most common malignant tumors worldwide. Macrophage-mediated innate immune responses play a crucial role in tumor development. This study revealed the mechanism of SHP-1 in regulating HCC progression. SHP-1 inhibits tumour development in vivo. Increasing SHP-1 expression in macrophages promotes the expression of p-SHP-1, SHP2, and p-SHP-2. In macrophages GM-CSF recruits SHP-2 to the GM-CSF receptor GM-CSFR induces p-SHP-2 dephosphorylation. GM-CSF recruits p-SHP-2 for dephosphorylation by up-regulating HoxA10HOXA10 activates the transcription of TGFβ2 by interacting with tandem cis-elements in the promoter thereby regulating the proliferation and migration of liver cancer cells. GM-CSF inhibits SHP-1 regulation of p-SHP-1, SHP2, and p-SHP-2 in macrophages. Detailed studies have shown that SHP-1 regulates SHP2 expression, and SHP-1 and SHP2 are involved in macrophage M2 polarisation. SHP-1 inhibits HOXA10 and TGFβ2 which in turn regulates the expression of the migration-associated proteins, MMP2/9, and the migration of hepatocellular carcinoma cells. Overexpression of SHP-1 inhibits macrophage M2 polarisation via the p-STAT3/6 signalling pathway Classical markers arginase-1, CD206, CD163 and regulate the expression of M2 polarisation cytokines IL-4 and IL-10. In addition, hypoxia-induced ROS inhibited SHP-1 regulation by suppressing the expression of p-SHP-1. The combined effect of GM-CSF and ROS significantly increased p-HOXA10/TGFβ2 and macrophage M2 polarisation, and the regulatory effect of ROS was significantly suppressed by GM-CSF knockdown. These findings suggest that increasing the expression of tyrosine phosphatase SHP-1 can inhibit hepatocellular carcinoma progression by modulating the SHP2/GM-CSF pathway in TAM and thus inhibit the progression of hepatocellular carcinoma.
Keywords: SHP-1, SHP2, GM-CSF, HOXA10, Hepatocellular carcinoma
Subject terms: Cancer microenvironment, Tumour-suppressor proteins, Cell migration
Introduction
Hepatocellular carcinoma (HCC) is a highly malignant tumor and ranks second in terms of global mortality rates. Despite significant advances in HCC treatment over the past few decades, its effectiveness remains limited. Furthermore, the prognosis for HCC patients is poor due to recurrence and metastasis. Therefore, the search for new treatment strategies and molecular targets is of great importance in improving the survival rate of HCC patients1,2.
Recent studies have shown that immunotherapy has significant potential in the field of tumor treatment3,4. Macrophages, as important immune cells, play a crucial role in tumor occurrence and development, particularly their innate immune response in tumor suppression5. Tumor-associated macrophages (TAMs) are one of the major immune cell components in liver cancer tissues, and they function in angiogenesis, promoting tumor cell proliferation and migration, and suppressing anti-tumor immunity6. To reflect the Th1/Th2 immune response, the classical concept suggests that macrophages have two distinct polarization states: M1 (induced by lipopolysaccharide and IFN-γ) and M2 (induced by IL-4 or IL-13). Studies have shown that TAMs primarily exhibit an M2-like phenotype, characterized by immune suppression and favoring tumor progression. Therefore, targeting M2-like TAMs and depleting them within TAMs, or reversing M2-like TAMs into an M1-like phenotype, is a potential immunotherapeutic strategy against tumors7,8.
Src homology 2 domain-containing protein tyrosine phosphatase 1 (SHP-1) is a widely expressed inhibitory protein tyrosine phosphatase (PTP)9. It has been implicated in inhibiting tumor development in various cancer types, such as lymphoma, lung cancer, prostate cancer, and breast cancer10–12. Evidence suggests that SHP-1 may contribute to the inhibition of aberrant innate immune cell activity. However, whether SHP-1 can inhibit the M2 polarization of TAMs remains largely unknown. The specific regulatory mechanisms of SHP-1 in liver cancer are still largely unknown. Therefore, this study aims to investigate the role and mechanism of SHP-1 in the progression of liver cancer, in order to provide new clues for a better understanding of liver cancer development and lay a foundation for the development of liver cancer treatment strategies based on SHP-1 regulatory pathways (Supplementary Information).
Methods
Animal modeling and grouping
Male Balb/c nude mice aged 6–8 weeks were purchased from Henan Scitek Bio-technology Co., Ltd. The mice were kept in a specific pathogen-free (SPF) animal facility with ad libitum access to water and food, under a 12-h light/12-h dark cycle. All animals received humane care. The study followed the guidelines for the ethical and humane use of laboratory animals, and all animal procedures were approved by the Medical Ethics Committee, Bethune International Peace Hospital (2021-KY-79). All methods reported in this manuscript are in accordance with ARRIVE guidelines.
Sixteen mice were randomly divided into two groups: THP-1-NC mimic and THP-1-SHP-1 mimic groups, with 8 mice in each group. The model group was induced to develop liver cancer by intraperitoneal injection of DEN (Sigma, N0258, i.p. 25 mg/kg)13. The THP-1-SHP-1 mimic group was injected with THP-1 cells infected with SHP-1 mimic. The cell suspension (5 × 106) was injected into the subcutaneous area of the upper right abdomen14. The mice were euthanized 21 days later (mice were deeply anesthetized and euthanized by cervical dislocation). Tumor tissues from the mice were collected, stained with hematoxylin and eosin (HE), embedded in paraffin, sectioned, and stained with suoxin red for 4 min. After rinsing, the slides were stained, mounted with resin, and observed and photographed under a microscope.
Cell culture
Transfection: SMMC-7721, a human liver cancer cell line, was purchased from Thermo Fisher Scientific Inc. (Shanghai, China, 61870036), THP-1, a human monocytic leukemia cell line, was purchased from Wuhan Punoise Life Technologies Co., Ltd. (Wuhan, China, CL-0233), RAW264.7, a mouse monocyte macrophage leukemia cell line, was purchased from Wuhan Punoise Life Technologies Co., Ltd. (Wuhan, China,CL-0190). SHP-1 mimic and NC mimic were purchased from Genecopoeia (Guangzhou, China). Mimics were diluted with serum-free dilution solution and mixed thoroughly with EntransterTM-R4000. After incubating at room temperature for 15 min, the transfection complex was dropped onto THP-1 cells and incubated for 24 h.
Macrophage were seeded at a density of 1 × 106 cells/mL in RPMI 1640 complete medium treated with PMA (100 ng/mL) for 6 h, followed by the addition of IL-13 (20 ng/mL) and IL-4 (20 ng/mL) for 18 h to generate M2 polarized macrophages.
The co-culture system of macrophages and HCC cells: PMA-treated THP-1 macrophages were seeded into the upper insert of a 6-well Transwell device and co-cultured with SMMC-7721 cells in the lower chamber of a 6-well plate. The two cell types did not directly contact each other. After co-culturing for 16 h, the cells were co-cultured with GM-CSF or without GM-CSF (25 ng/mL R&D Systems) and under normoxic (20% O2) or hypoxic (1% O2) conditions for 24 h.
Western blot
RIPA lysis buffer (Thermo Fisher Scientific) was used to lyse and extract proteins from THP-1 and SMMC7721 cells. The protein concentration was determined using a BCA protein assay kit (KeyGen Biotech). Then, 20 μg of protein was separated by 10% SDS-PAGE gel electrophoresis and transferred to PVDF membranes. The membranes were incubated with 5% skim milk for 2 h, followed by overnight incubation at 4 °C with primary antibodies. After incubation with secondary IgG (1:5000, Abcam) at room temperature for 2 h, the protein bands were quantified using ImageJ software. The primary antibodies used were SHP-1 (1:1000, Abcam), p-SHP-1 (1:1000, Thermo Fisher Scientific Inc.), SHP-2 (1:1000, Abcam), p-SHP2 (1:1000, Abcam), HOXA10 (1:1000, Abcam), TGFβ (1:1000, Abcam), MMP2 (1:1000, Abcam), MMP-9 (1:200, Abcam), p-STAT3 (1:1000, Abcam), p-STAT6 (1:1000, Abcam), IL-10 (1:1000, Abcam), IL-4 (1:2000, Abcam), and GAPDH (1:1000, Abcam).
Cell scratching and immunofluorescence
Single-cell suspensions were dropped onto culture plates containing slides, and after 24 h of incubation at 37 °C in a 5% CO2 incubator, the slides were taken out. The cells were fixed with 4% paraformaldehyde for 30 min, blocked with 5% goat serum at room temperature for 30 min, incubated with p-SHP-1 antibody (Abcam) overnight at 4 °C, and then incubated with fluorescent goat anti-rabbit IgG secondary antibody (diluted 1:500 in PBS) (Abcam, ab150081) in a dark environment at room temperature for 1 h. Images were captured using a fluorescence microscope.
Cell scratching
HCC cells from the co-culture system were collected and suspended in DMEM. After seeding the cells in a 6-well culture plate, a scratch was made using a 200 μL plastic pipette tip. The width of the scratch was measured and recorded by microscopy after 0 h, and the cells were incubated in the incubator for 48 h before measuring the scratch width again.
Transwell assay
SMMC-7721 cells were resuspended in serum-free medium and seeded in the upper chamber of a Transwell device. The lower chamber was pre-filled with 10% FBS medium. After incubation at room temperature for 24 h, the cells were fixed with 4% paraformaldehyde for 10 min, stained with 0.1% crystal violet for 20 min, and counted under a microscope.
Cell cloning formation
The co-cultured cells were digested with trypsin-collagenase and then diluted to a density of 1 × 104/mL. The cells were seeded in a 6-well plate at a density of 400 cells per well and cultured for 2 weeks. The cells were fixed with 4% paraformaldehyde for 30 min, stained with 0.1% crystal violet for 15 min, and photographed.
CCK8 assay
Cell suspensions were prepared using culture medium containing 5% fetal bovine serum. Then, 100 μL of cell suspension was seeded in a 96-well plate, and 10 μL of CCK-8 solution was added to each well. After incubation at 37 °C and 5% CO2 for 4 h, the absorbance at 450 nm was measured.
Statistical analysis
The data are presented as the mean ± standard deviation (SD) of at least three independent experiments. Multiple group comparisons were performed using one-way analysis of variance (ANOVA), and two-group comparisons were analyzed using t-tests. Statistical analysis was conducted using GraphPad Prism software. *P < 0.05 was considered statistically significant.
Results
Upregulation of TAM SHP-1 inhibits the progression of liver cancer
To study the effects of SHP-1 on tumor development in mice, we injected NC-macrophage and SHP-1 mimic-macrophage into liver cancer mice for animal modeling. The results of HE staining showed that the tumor area in SHP-1 mimic mice was significantly smaller than in NC mice (Fig. 1).
Figure 1.
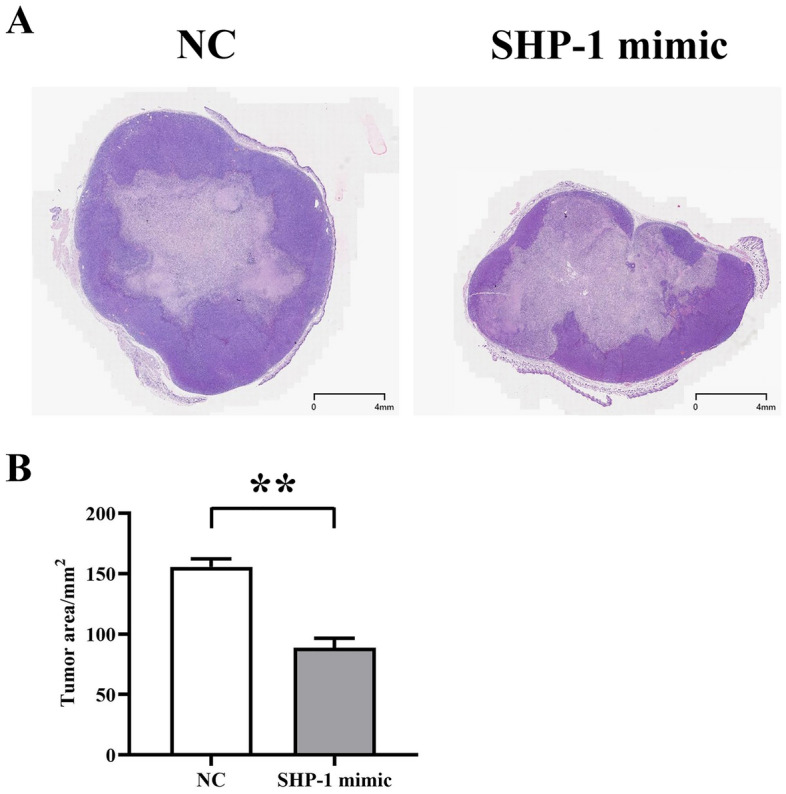
Representative images showing the inhibition of liver cancer progression in mice by overexpression of SHP-1. (A) Comparison of tumor volume between the two groups of mice. (B) Comparison of tumor area between the two groups of mice.
SHP-1 overexpression promotes SHP2 protein expression in macrophage, and GM-CSF and hypoxia inhibit the expression of p-SHP-1 and p-SHP2
To investigate the effects of GM-CSF and hypoxia on the protein expression of p-SHP-1, SHP2, p-SHP2, we analyzed the expression of SHP-1, p-SHP-1, SHP2, p-SHP2 after GM-CSF and hypoxia interference. The results showed that compared to the NC group, the overexpression of SHP-1 in the SHP mimic group promoted the expression of p-SHP-1, SHP2, and p-SHP2. GM-CSF inhibited the protein expression of p-SHP-1, SHP2, and p-SHP2 in macrophage. When comparing the NC group with the GM-CSF + NC group and the SHP-1 mimic group with the GM-CSF + SHP-1 mimic group, GM-CSF was found to promote the protein expression of p-SHP-1, SHP2, and p-SHP2 in macrophage. Comparing the NC group with the GM-CSF + SHP-1 mimic group, GM-CSF was found to inhibit the regulation of p-SHP-1, SHP2, and p-SHP2 by SHP-1 in macrophage. Hypoxia drives an increase in reactive oxygen species (ROS) in tumor cells15, so we added a low oxygen culture experiment to investigate the effects of oxidative stress and GM-CSF on protein expression in macrophage. The combined effect of GM-CSF and oxidative stress significantly enhanced the inhibitory effect of GM-CSF (Fig. 2A). Immunofluorescence staining of p-SHP-1 yielded consistent results (Fig. 2B).
Figure 2.

Regulation of protein expression in macrophage by overexpression of SHP-1 and the influence of GM-CSF and hypoxia. (A) Western blot results showing the expression of SHP-1, p-SHP-1, SHP2, and p-SHP-2 proteins in macrophage under the influence of SHP-1, GM-CSF, and hypoxia. (B) Immunofluorescence staining showing the effect of SHP-1, GM-CSF, and hypoxia on the expression of p-SHP-1 protein in macrophage.
Effects of SHP-1 overexpression, GM-CSF, and hypoxia on p-HOXA10/TGFβ2 protein expression in macrophage and migration of SMMC7721 cells
To further study the mechanism of SHP-1 inhibition of liver cancer, we analyzed the expression of p-HOXA10/TGFβ2 in macrophage and the migration ability of SMMC7721 cells under the interference of GM-CSF and hypoxia. The results showed that compared to the NC group, SHP-1 inhibited the protein expression of p-HOXA10/TGFβ2 in macrophage. GM-CSF promoted the protein expression of p-HOXA10/TGFβ2 and M2 polarization-related proteins in macrophage and inhibited the inhibitory effect of SHP-1. Under the combined action of GM-CSF and hypoxia, the protein expression of p-HOXA10/TGFβ2 in macrophage was significantly increased, and the inhibitory effect of SHP-1 was insignificant (P > 0.05) (Fig. 3A). SHP-1 inhibited the protein expression of MMP-2 and MMP-9 in liver cancer cells. SHP-1 can inhibit the expression of MMP-2 and MMP-9 protein in hepatocellular carcinoma cells related to migration.. Under the combined action of GM-CSF and oxidative stress, the protein expression of MMP-2 and MMP-9 in liver cancer cells was significantly increased, and the inhibitory effect of SHP-1 was insignificant (Fig. 3B). Cell scratching and Transwell assay results showed that SHP-1 inhibited the migration ability of liver cancer cells, GM-CSF promoted the migration of liver cancer cells and suppressed the inhibitory ability of SHP-1, and the combined action of GM-CSF and oxidative stress promoted the migration of liver cancer cells (Fig. 3C,D).
Figure 3.

Effects of SHP-1 overexpression, GM-CSF, and hypoxia on the expression of p-HOXA10/TGFβ2 proteins in macrophage and migration of SMMC7721 cells. (A) Influence of SHP-1, GM-CSF, and hypoxia on the expression of p-HOXA10/TGFβ2 proteins in macrophage. (B) Influence of SHP-1, GM-CSF, and hypoxia on the expression of MMP-2 and MMP-9 proteins in SMMC7721 cells. (C) Evaluation of the migration ability of SMMC7721 cells by cell scratch assay under the influence of SHP-1, GM-CSF, and hypoxia. (D) Evaluation of the migration ability of SMMC7721 cells by Transwell assay under the influence of SHP-1, GM-CSF, and hypoxia.
Effects of SHP-1 overexpression, GM-CSF, and hypoxia on the expression of M2 polarization-related proteins in macrophage and the proliferation of liver cancer cells
To further study the mechanism of SHP-1 inhibition of liver cancer, we analyzed the expression of M2 polarization-related proteins in macrophage and the proliferation of SMMC7721 cells under the interference of GM-CSF, GM-CSF knockdown and hypoxia. The results showed that compared to the NC group, SHP-1 inhibited the protein expression of p-STAT3, p-STAT6, and M2 polarization-related proteins IL-10, IL-4, Arginase-1, CD206 and CD163 in macrophage. GM-CSF promoted the protein expression of p-STAT3, p-STAT6, IL-10, IL-4, Arginase-1, CD206 and CD163 in macrophage and suppressed the inhibitory effect of SHP-1. Under the combined action of GM-CSF and hypoxia, the protein expression of p-STAT3, p-STAT6, IL-10, IL-4, Arginase-1, CD206 and CD163 in macrophage was significantly increased, and the inhibitory effect of SHP-1 was insignificant (P > 0.05). Protein expression of p-STAT3, p-STAT6 and M2 polarization-associated proteins IL-10, IL-4, Arginase-I, CD206 and CD163 was significantly suppressed in macrophages under the combined effect of GM-CSF knockdown and hypoxia (Fig. 4A).
Figure 4.

Effects of SHP-1 overexpression, GM-CSF, GM-CSF knockdown, and hypoxia on the expression of M2 polarization-related proteins in macrophage and proliferation of liver cancer cells. (A) Influence of SHP-1 overexpression, GM-CSF, GM-CSF knockdown, and hypoxia on the expression of p-STAT3, p-STAT6, IL-10, IL-4, and Arginase-I proteins involved in M2 polarization in macrophage. (B) Evaluation of the proliferation ability of SMMC7721 cells by clonogenic assay under the influence of SHP-1, GM-CSF, GM-CSF knockdown, and hypoxia. (C) Evaluation of the proliferation ability of SMMC7721 cells by CCK-8 assay under the influence of SHP-1, GM-CSF, GM-CSF knockdown, and hypoxia.
Cell cloning and CCK8 assay results showed that SHP-1 inhibited the proliferation of liver cancer cells, GM-CSF promoted the proliferation of liver cancer cells and suppressed the inhibitory effect of SHP-1, and the combined action of GM-CSF and oxidative stress promoted the proliferation of liver cancer cells. GM-CSF knockdown and oxidative stress in conjunction with hypoxia inhibits the proliferation of hepatocellular carcinoma cells (Fig. 4B,C).
Discussion
Liver cancer, as a highly malignant tumor, faces many challenges in its treatment process. Therefore, it is of great significance to search for new treatment strategies and molecular targets. In recent years, immunotherapy has been recognized as having tremendous potential in cancer treatment. This study aimed to explore the role of tyrosine phosphatase SHP-1 in the progression of liver cancer and lay the foundation for the development of liver cancer treatment strategies based on the SHP-1 regulatory pathway. Here, we demonstrated that hypoxia and GM-CSF synergistically promoted M2 polarization and migration and proliferation of liver cancer through the inhibition of SHP-1 and SHP-2 phosphorylation and the increase of p-HOXA10 expression (Fig. 5).
Figure 5.

Research on the mechanism of tyrosine phosphatase SHP-1 inhibiting liver cancer progression through the regulation of SHP2/GM-CSF pathway in TAMs.
Protein tyrosine phosphatases (PTPs) serve as regulators of cellular functions including differentiation, metabolism, migration, and survival. PTPs counteract tyrosine kinases by removing phosphate molecules from molecular signaling residues, thereby inhibiting signal transduction. Two PTPs, SHP-1 and SHP-2 (protein tyrosine phosphatase 1 and 2 with SH2 domains, respectively), as well as another inhibitory phosphatase, SH2-containing inositol phosphatase (SHIP), are crucial for cellular functions. Recent studies have shown the dysregulation of SHP-1, SHP-2, and SHIP is associated with uncontrolled cell growth and metabolism16. In most systems, SHP-1 has been demonstrated as a negative regulator and shown to directly bind to the regulated receptor itself or associated co-receptors, whereas SHP-2 is considered to positively promote signal transduction17. However, recent studies have revealed complex associations between SHP-1, SHP-2, and tumor occurrence and development. For example, the research results of Beldi-Ferchiou A showed that the gene expression of SHP-1 and SHP-2 effectively inhibited the activation of the JAK/STAT pathway and induced apoptosis in tumor cells18. Xiao-hua Tao’s study demonstrated that SHP-1 and SHP-2 play promoting roles in the development of human papillomavirus-induced condyloma acuminatum and cervical cancer19. SHP-1 down-regulated CSF1R expression, which induces macrophage M2 polarization, and inhibited STAT3/NFκB signaling, leading to suppression of macrophage survival and disturbance of macrophage polarization20.
It has been shown that tumor-associated macrophages (TAMs), especially M2 subtype macrophages, promote cancer cell metastasis by inducing EMT21. M2 polarization of TAMs is a multifactorial, multistep, and complex pathological process. IL-10 is a major cell factor involved in the negative feedback control of inflammation. Yannick Degboé’s research has shown that IL-10 signals through the STAT3 pathway to promote macrophage phenotypic transformation22. The anti-inflammatory cytokine IL-10 activates innate immune signaling pathways to promote M2 polarization of macrophages23. CD206 and Arginase-I are markers of macrophage polarization24,25. Previous studies have shown that STAT can inhibit nuclear factor kappaB (NF-κB) expression through the CD206/Arginase-I/IL-4/IL-10 signaling pathway26–29. Importantly, the activation of the NFκB-dependent inflammatory signaling pathway drives SHP2 gene expression30.
Our experimental results showed that increasing the expression of SHP-1 can inhibit tumor development in vivo, further confirming the importance of SHP-1 in the suppression of liver cancer, which is consistent with other research results31. Furthermore, overexpression of SHP-1 promoted the expression of SHP2/GM-CSF pathway-regulated proteins, including p-SHP-1, SHP2, and p-SHP2. Both GM-CSF and hypoxia regulated the expression of these proteins, with GM-CSF inhibiting the regulatory role of SHP-1 in macrophage. To investigate the effects of GM-CSF and hypoxia-induced oxidative stress on SHP-1, we conducted further studies. In this study, we found that SHP-1 lost its inhibitory effect on the expression of proteins p-STAT3, p-STAT6, IL-10, IL-4, and Arginase-I associated with M2 polarization in macrophages under the combined effect of GM-CSF and hypoxic environment. In addition, GM-CSF also promoted the expression of p-HOXA10 and TGFβ2 in liver cancer cells and inhibited the inhibitory effect of SHP-1. These results further support the important role of SHP-1 in regulating the progression of liver cancer.
Although we focused on p-HOXA10 and TGF-β2, we also recognize the possible effects of other M2-type polarization-related cytokines or proteins on SHP-1. Our study did not exclude the presence of these factors, but rather focused on analyzing the effects of GM-CSF and the hypoxic environment on the SHP-1 regulatory network. Future studies could further explore the role of other cytokines or proteins in this process to fully understand the complex mechanisms of hepatocellular carcinoma development and immune regulation.
In summary, this study revealed the mechanism by which SHP-1 inhibits the progression of liver cancer by regulating the SHP2/GM-CSF pathway in TAMs. Overexpression of SHP-1 can inhibit the proliferation and migration ability of liver cancer cells and suppress the expression of M2 polarization-related proteins. However, under the combined action of GM-CSF and hypoxia, the inhibitory effect of SHP-1 becomes limited and contributes to the proliferation and migration of liver cancer cells. These research findings provide new clues for the development of liver cancer treatment strategies based on the SHP-1 regulatory pathway and offer new avenues for improving the effectiveness of liver cancer treatment. However, further research is needed to validate these findings and explore deeper molecular mechanisms.
Supplementary Information
Author contributions
QW participated in planning, design the study, methodology, data analysis and writing—original draft, SL performed the and data analysis, writing—review and editing, GH participated in planning, design the study and investigation, participated in planning and design the study. All authors read and approved the manuscript.
Funding
Open access funding provided by Hebei Provincial Health and Hygiene Commission (20220245).
Data availability
This published article contains all of the data that were examined throughout this investigation.
Competing interests
The authors declare no competing interests.
Footnotes
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
The online version contains supplementary material available at 10.1038/s41598-024-59725-w.
References
- 1.Fu XT, Song K, Zhou J, Shi YH, Liu WR, Shi GM, Gao Q, Wang XY, Ding ZB, Fan J. Tumor-associated macrophages modulate resistance to oxaliplatin via inducing autophagy in hepatocellular carcinoma. Cancer Cell Int. 2019;19:71. doi: 10.1186/s12935-019-0771-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Yao RR, Li JH, Zhang R, Chen RX, Wang YH. M2-polarized tumor-associated macrophages facilitated migration and epithelial–mesenchymal transition of HCC cells via the TLR4/STAT3 signaling pathway. World J. Surg. Oncol. 2018;16(1):9. doi: 10.1186/s12957-018-1312-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Cao J, Yan Q. Cancer epigenetics, tumor immunity, and immunotherapy. Trends Cancer. 2020;6(7):580–592. doi: 10.1016/j.trecan.2020.02.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Du H, Xu T, Cui M. cGAS-STING signaling in cancer immunity and immunotherapy. Biomed. Pharmacother. 2021;133:110972. doi: 10.1016/j.biopha.2020.110972. [DOI] [PubMed] [Google Scholar]
- 5.Sun Y, Hu H, Liu Z, Xu J, Gao Y, Zhan X, Zhou S, Zhong W, Wu D, Wang P, et al. Macrophage STING signaling promotes NK cell to suppress colorectal cancer liver metastasis via 4–1BBL/4-1BB co-stimulation. J. Immunother. Cancer. 2023 doi: 10.1136/jitc-2022-006481. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Chen D, Zhang X, Li Z, Zhu B. Metabolic regulatory crosstalk between tumor microenvironment and tumor-associated macrophages. Theranostics. 2021;11(3):1016–1030. doi: 10.7150/thno.51777. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Pathria P, Louis TL, Varner JA. Targeting tumor-associated macrophages in cancer. Trends Immunol. 2019;40(4):310–327. doi: 10.1016/j.it.2019.02.003. [DOI] [PubMed] [Google Scholar]
- 8.Chen D, Xie J, Fiskesund R, Dong W, Liang X, Lv J, Jin X, Liu J, Mo S, Zhang T, et al. Chloroquine modulates antitumor immune response by resetting tumor-associated macrophages toward M1 phenotype. Nat. Commun. 2018;9(1):873. doi: 10.1038/s41467-018-03225-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Watson HA, Wehenkel S, Matthews J, Ager A. SHP-1: The next checkpoint target for cancer immunotherapy? Biochem. Soc. Trans. 2016;44(2):356–362. doi: 10.1042/BST20150251. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Chi D, Wang D, Zhang M, Ma H, Chen F, Sun Y. CLEC12B suppresses lung cancer progression by inducing SHP-1 expression and inactivating the PI3K/AKT signaling pathway. Exp. Cell Res. 2021;409(2):112914. doi: 10.1016/j.yexcr.2021.112914. [DOI] [PubMed] [Google Scholar]
- 11.Ren X, Peng M, Xing P, Wei Y, Galbo PM, Jr, Corrigan D, Wang H, Su Y, Dong X, Sun Q, et al. Blockade of the immunosuppressive KIR2DL5/PVR pathway elicits potent human NK cell-mediated antitumor immunity. J. Clin. Investig. 2022;132(22):163620. doi: 10.1172/JCI163620. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Dempke WCM, Uciechowski P, Fenchel K, Chevassut T. Targeting SHP-1, 2 and SHIP pathways: A novel strategy for cancer treatment? Oncology. 2018;95(5):257–269. doi: 10.1159/000490106. [DOI] [PubMed] [Google Scholar]
- 13.Filliol A, Saito Y, Nair A, Dapito DH, Yu LX, Ravichandra A, Bhattacharjee S, Affo S, Fujiwara N, Su H, et al. Opposing roles of hepatic stellate cell subpopulations in hepatocarcinogenesis. Nature. 2022;610(7931):356–365. doi: 10.1038/s41586-022-05289-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Yu J, Green MD, Li S, Sun Y, Journey SN, Choi JE, Rizvi SM, Qin A, Waninger JJ, Lang X, et al. Liver metastasis restrains immunotherapy efficacy via macrophage-mediated T cell elimination. Nat. Med. 2021;27(1):152–164. doi: 10.1038/s41591-020-1131-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Perillo B, Di Donato M, Pezone A, Di Zazzo E, Giovannelli P, Galasso G, Castoria G, Migliaccio A. ROS in cancer therapy: The bright side of the moon. Exp. Mol. Med. 2020;52(2):192–203. doi: 10.1038/s12276-020-0384-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Hao F, Wang C, Sholy C, Cao M, Kang X. Strategy for leukemia treatment targeting SHP-1,2 and SHIP. Front. Cell Dev. Biol. 2021;9:730400. doi: 10.3389/fcell.2021.730400. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Lorenz U. SHP-1 and SHP-2 in T cells: Two phosphatases functioning at many levels. Immunol. Rev. 2009;228(1):342–359. doi: 10.1111/j.1600-065X.2008.00760.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Beldi-Ferchiou A, Skouri N, Ben Ali C, Safra I, Abdelkefi A, Ladeb S, Mrad K, Ben Othman T, Ben Ahmed M. Abnormal repression of SHP-1, SHP-2 and SOCS-1 transcription sustains the activation of the JAK/STAT3 pathway and the progression of the disease in multiple myeloma. PLoS One. 2017;12(4):e0174835. doi: 10.1371/journal.pone.0174835. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Takagane K, Umakoshi M, Itoh G, Kuriyama S, Goto A, Tanaka M. SKAP2 suppresses inflammation-mediated tumorigenesis by regulating SHP-1 and SHP-2. Oncogene. 2022;41(8):1087–1099. doi: 10.1038/s41388-021-02153-1. [DOI] [PubMed] [Google Scholar]
- 20.Hong SY, Lu YT, Chen SY, Hsu CF, Lu YC, Wang CY, Huang KL. Targeting pathogenic macrophages by the application of SHP-1 agonists reduces inflammation and alleviates pulmonary fibrosis. Cell Death Dis. 2023;14(6):352. doi: 10.1038/s41419-023-05876-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Wei C, Yang C, Wang S, Shi D, Zhang C, Lin X, Liu Q, Dou R, Xiong B. Crosstalk between cancer cells and tumor associated macrophages is required for mesenchymal circulating tumor cell-mediated colorectal cancer metastasis. Mol. Cancer. 2019;18(1):64. doi: 10.1186/s12943-019-0976-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Liu Q, Yang C, Wang S, Shi D, Wei C, Song J, Lin X, Dou R, Bai J, Xiang Z, et al. Wnt5a-induced M2 polarization of tumor-associated macrophages via IL-10 promotes colorectal cancer progression. Cell Commun. Signaling CCS. 2020;18(1):51. doi: 10.1186/s12964-020-00557-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Chistyakov DV, Gavrish GE, Goriainov SV, Chistyakov VV, Astakhova AA, Azbukina NV, Sergeeva MG. Oxylipin profiles as functional characteristics of acute inflammatory responses in astrocytes pre-treated with IL-4, IL-10, or LPS. Int. J. Mol. Sci. 2020;21(5):1780. doi: 10.3390/ijms21051780. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Bardi GT, Smith MA, Hood JL. Melanoma exosomes promote mixed M1 and M2 macrophage polarization. Cytokine. 2018;105:63–72. doi: 10.1016/j.cyto.2018.02.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Wang J, Xu L, Xiang Z, Ren Y, Zheng X, Zhao Q, Zhou Q, Zhou Y, Xu L, Wang Y. Microcystin-LR ameliorates pulmonary fibrosis via modulating CD206(+) M2-like macrophage polarization. Cell Death Dis. 2020;11(2):136. doi: 10.1038/s41419-020-2329-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Amini-Farsani Z, Yadollahi-Farsani M, Arab S, Forouzanfar F, Yadollahi M, Asgharzade S. Prediction and analysis of microRNAs involved in COVID-19 inflammatory processes associated with the NF-kB and JAK/STAT signaling pathways. Int. Immunopharmacol. 2021;100:108071. doi: 10.1016/j.intimp.2021.108071. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Liu L, Guo H, Song A, Huang J, Zhang Y, Jin S, Li S, Zhang L, Yang C, Yang P. Progranulin inhibits LPS-induced macrophage M1 polarization via NF-кB and MAPK pathways. BMC Immunol. 2020;21(1):32. doi: 10.1186/s12865-020-00355-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Jaruga B, Hong F, Sun R, Radaeva S, Gao B. Crucial role of IL-4/STAT6 in T cell-mediated hepatitis: Up-regulating eotaxins and IL-5 and recruiting leukocytes. J. Immunol. 2003;171(6):3233–3244. doi: 10.4049/jimmunol.171.6.3233. [DOI] [PubMed] [Google Scholar]
- 29.Yuan ZH, Feng L, Jiang WD, Wu P, Liu Y, Jiang J, Kuang SY, Tang L, Zhou XQ. Dietary choline-enhanced skin immune response of juvenile grass carp might be related to the STAT3 and NF-kB signaling pathway (Ctenopharyngodon idella) Front. Nutr. 2021;8:652767. doi: 10.3389/fnut.2021.652767. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Dorenkamp M, Nasiry M, Semo D, Koch S, Löffler I, Wolf G, Reinecke H, Godfrey R. Pharmacological targeting of the RAGE-NFκB signalling axis impedes monocyte activation under diabetic conditions through the repression of shp-2 tyrosine phosphatase function. Cells. 2023;12(3):513. doi: 10.3390/cells12030513. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Wen LZ, Ding K, Wang ZR, Ding CH, Lei SJ, Liu JP, Yin C, Hu PF, Ding J, Chen WS, et al. SHP-1 acts as a tumor suppressor in hepatocarcinogenesis and HCC progression. Cancer Res. 2018;78(16):4680–4691. doi: 10.1158/0008-5472.CAN-17-3896. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
This published article contains all of the data that were examined throughout this investigation.