
Fig. 1 |. Mechanisms of transcription activation in cis by long non-coding RNAs.
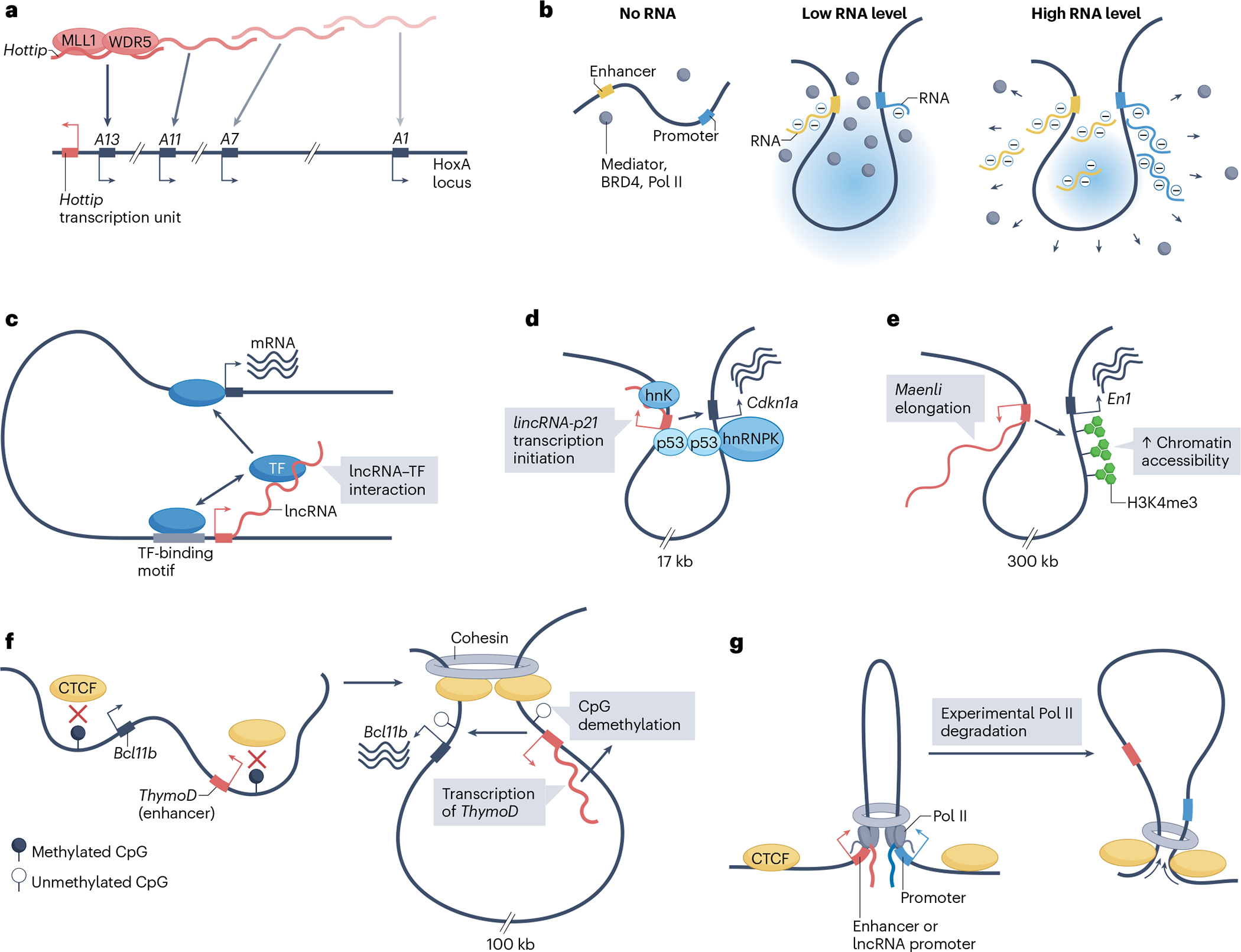
a, The long non-coding RNA (lncRNA) Hottip is expressed from the HoxA locus and serves as a scaffold for the local recruitment of the histone methyltransferase complex comprising MLL1 (also known as KMT2A) and WDR5 to HoxA gene (A1–A13) sites of transcription. Consistent with local activity, RNAi-mediated depletion of Hottip preferentially affects Hottip proximal, compared with distal, HoxA genes (fading colour gradient). b, Local RNA abundance provides feedback on transcription initiation. Left: the Mediator complex, the histone-acetylation reader bromodomain-containing protein 4 (BRD4) and RNA polymerase II (Pol II) are present in low abundance at transcriptionally inactive promoter and enhancer elements. Middle: upon transcription initiation, nascent RNAs produced from promoter and enhancer regions nucleate the formation of a condensate, which increases the local concentration of transcription regulators, thereby causing a burst in transcription. Right: as transcription proceeds, the increase in local RNA abundance beyond a certain threshold generates electrostatic repulsive forces that disperse the transcriptional condensates, thereby ending the transcription burst. c, Many transcription factors (TF) have RNA-binding domains, which potentially interact with nascent transcripts, including of lncRNAs. These lncRNA–TF interactions could contribute to the targeting or the strength of association of the TFs to their genomic target sites by taking advantage of their pre-existing 3D proximity. d, lincRNA-p21 and its cis-activated target, the neighbouring gene Cdkn1a (encoding p21), are in 3D proximity, and are co-regulated by the TF p53. Transcription initiation of lincRNA-p21 is sufficient to enhance the expression of Cdkn1a by creating a scaffold for the recruitment of the Cdkn1a transcriptional co-activator heterogeneous nuclear ribonucleoprotein K (hnRNPK). e, Transcription elongation of the lncRNA Maenli increases local histone H3 trimethylated at lysine 4 (H3K4me3), which is a mark of transcriptionally active chromatin, and promotes the expression of the neighbouring gene En1. f, Transcription of a lncRNA in the Bcl11b locus named thymocyte differentiation factor (ThymoD) promotes the demethylation of CTCF-binding sites and, therefore, CTCF recruitment and chromatin reorganization. This process brings the ThymoD-associated enhancer region in proximity with their target, the promoter of Bcl11b, resulting in transcription activation. g, Pol II recruitment to enhancers and promoters blocks chromatin-loop extrusion and stabilizes the loops at a configuration that brings active enhancers in proximity of active promoters. Experimental degradation of Pol II leads to the formation of larger loops, extrusion of which is now limited by CTCF.