

Abstract
Zinc is a transition metal that displays wide physiological implications ranging from participation in hundreds of enzymes and proteins to normal growth and development. In the reproductive tract of both sexes, zinc maintains a functional role in spermatogenesis, ovulation, fertilization, normal pregnancy, fetal development, and parturition. In this work, we review evidence to date regarding the importance of zinc in oocyte maturation and development, with emphasis on the role of key zinc-binding proteins, as well as examine the effects of zinc and reactive oxygen species (ROS) on oocyte quality and female fertility. We summarize our current knowledge about the participation of zinc in the developing oocyte bound to zinc finger proteins as well as loosely bound zinc ion in the intracellular and extracellular environments. These include aspects related to (1) the impact of zinc deficiency and overwhelming production of ROS under inflammatory conditions on the offset of the physiological antioxidant machinery disturbing biomolecules, proteins, and cellular processes, and their role in contributing to further oxidative stress; (2) the role of ROS in modulating damage to proteins containing zinc, such as zinc finger proteins and nitric oxide synthases (NOS), and expelling the zinc resulting in loss of protein function; and (3) clarify the different role of oxidative stress and zinc deficiency in the pathophysiology of infertility diseases with special emphasis on endometriosis-associated infertility.
Keywords: Zinc finger protein, Oocyte, Oxidative stress, Reactive oxygen species, Nitric oxide synthase, Infertility
Introduction
Zinc, a transition metal and the second most abundant trace element in the body, is required for the catalytic activity of hundreds of enzymes. An estimated 3000 proteins bind zinc to maintain their structural integrity and function [1]. Trace amounts of zinc are crucial for living organisms as they participate in normal growth and development, gene transcription, and protein, lipid, and nucleic acid metabolism [2]. Recent research has identified the detrimental role of zinc in the human body in immune function, wound repair, and reproduction [3–5]. Fluctuations of intracellular zinc are critical to create a fertilizable egg and the process of fertilization in mammals requires a fine-tuned and well-regulated process including progression of the developing oocyte through meiotic arrest and resumption. Several factors can impact this natural development process, leading to decreased oocyte quality, anovulation, spontaneous oocyte arrest, and apoptosis, namely chronological aging, inflammation, environmental factors, and gynecological disorders such as endometriosis and polycystic ovarian syndrome (PCOS) [6–8].
As sufficient levels of zinc are required in an abundance of physiological processes, the human body requires constant intake of zinc either through diet or supplementation; thus, assessment of zinc status is significantly hindered by the tight regulation of zinc homeostasis. Serum zinc is commonly utilized as a marker of zinc status but free zinc in the serum is found mainly bound to proteins such as albumin (low affinity), α2-macroglobulin (A2M, moderate affinity), and transferrin (high affinity) [9]. Although blood collection is a typically easy process, zinc levels are affected by several alternative factors including anemia and pregnancy [10]. It has been suggested that a more elusive method to measure zinc is through biomarkers such as the expression of metallothionein and/or zinc transporters in leukocytes [11].
Recent discussion has highlighted the importance of zinc as an antioxidant due to zinc deficiency being correlated with an increase in reactive oxygen species (ROS) and reactive nitrogen species (RNS), although the exact mechanism is poorly understood [12–15]. It is speculated that the link between low zinc and increased ROS activity is due to decreased activity of antioxidant enzymes that require zinc, such as Cu/Zn SOD [14]. Zinc participates in antioxidant activity, yet it is not redox active nor directly interacts with ROS; however, zinc-binding proteins can be targeted by ROS/RNS. In biological processes, zinc finger proteins play important roles in transcription and DNA repair, as their zinc finger motif mediates protein-DNA, protein-RNA, and protein–protein interactions [16, 17]. The method by which certain zinc finger proteins are specifically targeted by ROS is not understood, as there is an abundant number of these proteins and cysteine residues. Upon constant exposure to overwhelming ROS may result in irreversible loss of protein function, abnormal modification, and physiological dysfunction [18, 19].
The maintenance of zinc concentrations at the cellular level can be accomplished through binding to proteins/enzymes or as labile zinc in intracellular compartments [1]. At the cellular level, zinc is mostly found in zinc-storing vesicles known as zincosomes, the nucleus, and distributed between the cytoplasm (bound to metallothioneins) and organelles [20]. Metallothioneins are zinc-chelating proteins that complex roughly 20% of intracellular zinc helping to protect against high cytotoxic metal concentrations and acting as mechanisms of zinc compartmentalization and zinc sequestration [20]. The distribution of zinc between the cytosol and organelles is accomplished through membrane channels or by zinc transporting proteins, keeping the overall cytosolic free zinc concentration to picomolar or nanomolar levels [9, 20]. The coordination of zinc release within the cell increasing cytosolic concentrations can impact gene expression, enzymatic activity, and cell signaling.
The disturbance of key zinc-binding proteins, including zinc finger proteins and nitric oxide synthases (NOSs), due to overwhelming ROS in the oocyte microenvironment may adversely impact oocyte quality through diminishing the ability of the oocyte to develop and be fertilized as described in Fig. 1 [21, 22]. The disrupted oxidant to antioxidant ratio in inflammatory diseases such as endometriosis or with the introduction of zinc-chelating agents by environmental toxins such as glyphosate may interfere with zinc bioavailability and protein functionality, thereby increasing oxidative stress and decreasing oocyte quality [8, 22–24]. As there are gaps within the field to understand the source, contribution, and importance of zinc in the oocyte, this review aims to summarize the current understanding of zinc in oocyte development and connect research regarding oxidative stress and zinc-binding proteins and enzymes (i.e., zinc finger proteins and NOS) to oocyte quality and fertility in inflammatory disorders.
Fig. 1.
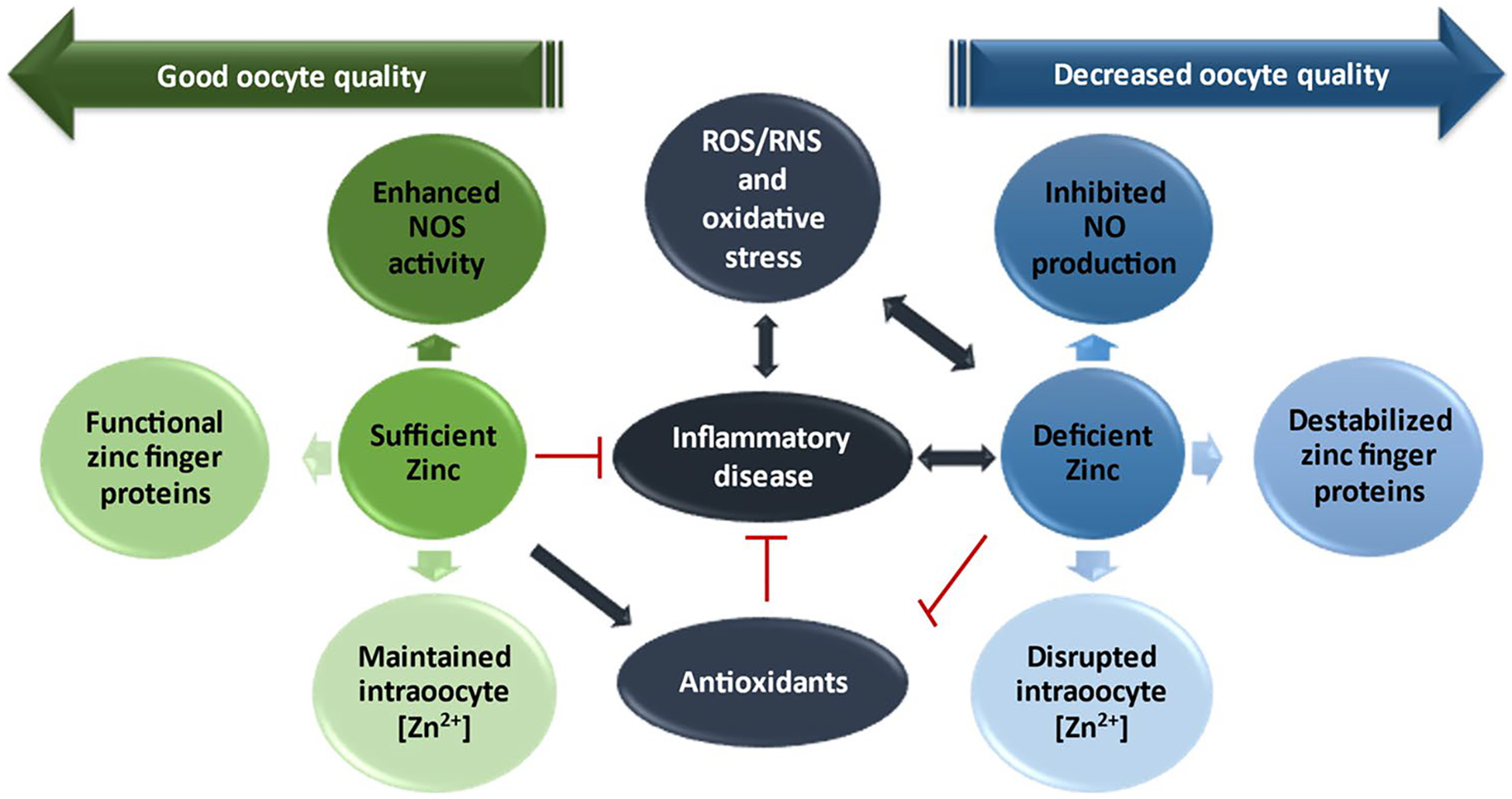
The implications of oxidative stress on zinc availability contributing to decreased oocyte quality. In the presence of sufficient levels of zinc and antioxidants, inflammation and the production of oxidative stress are decreased allowing for (1) appropriate levels of nitric oxide (NO) from nitric oxide synthase (NOS) activity and (2) appropriate levels of zinc to bind to zinc finger proteins and participate in physiological processes requiring zinc such as zinc sparks. When this pathway is followed, oocyte quality is maintained through metaphase II arrest allowing for fertilization. In the event of overwhelming oxidative stress where zinc is deficient and antioxidant function goes down, i.e., in SOD where zinc is required for activity, resulting in destruction of NOS and NO consumption, inadequate zinc for zinc finger proteins, and disruption in zinc ion concentration ([Zn2+]) and homeostasis. In this detrimental pathway, the oocyte may not develop properly and/or prematurely exits MII arrest resulting in decreased maturation competence and inability to be fertilized
Methodology
An extensive literature review was conducted through the online databases PubMed, Science Direct, and Springer Link using the keywords zinc, oocyte development, oocyte maturation, control of meiotic arrest, zinc finger proteins/zinc-binding proteins, EMI2, reactive oxygen species/ROS, infertility, and antioxidants. Of the results found, included references are peer reviewed articles written in the English language. Individual hand searches of the references of retrieved literature was conducted. Ninety-two original articles presenting an overview of oocyte meiotic development, reactive oxygen species, inflammatory conditions and infertility, and zinc in the female reproductive process were included in this review.
Overview of Oocyte Development and the Role of Zinc
Humans are born with all the eggs they will ever have arrested in the diplotene stage of prophase I of the cell cycle, characterized by a large nucleus covered by a nuclear envelope known as the germinal vesicle (GV) [25]. A select group of follicles are stimulated each ovarian cycle by a surge in follicle-stimulating hormone (FSH) in which one of the recruited follicles acquires an increasing number of granulosa cells with enough FSH receptors to respond to stimulation from the falling FSH levels, becoming the dominant follicle. The oocyte in the dominant follicle is stimulated by a rise in luteinizing hormone (LH) before ovulation to resume meiosis. During this time, a rise in intracellular zinc is crucial to allow for the progression from prophase I to metaphase II without an intervening interphase [1, 25–27].
Prophase I Through Fertilization
Cyclic nucleotides cyclic adenosine monophosphate (cAMP) and cyclic guanosine monophosphate (cGMP) have been determined as critical for maintaining meiotic arrest [28]. Before the LH-surge occurs, cGMP from granulosa cells is transferred through gap junctions to the oocyte and antagonizes the activity of phosphodiesterase 3A (PDE3A) [28, 29]. The surge in LH results in a decrease in follicular cGMP induced by LH with increased phosphorylation of connexin 43 in the granulosa cells [30, 31]. Connexin 43 has been determined to be involved in the permeability of the gap junctions between the granulosa cells and oocytes, and phosphorylation of connexin 43 decreases the permeability [31]. It is thus speculated a closure in the gap junctions, in part, may contribute to the decrease in cGMP. The decrease in cGMP results in an increase in PDE3A activity [32, 33] which, when active, hydrolyzes cAMP allowing for decreased cAMP activity and progression through meiosis [27]. The degradation of cAMP signals the breakdown of the germinal vesicle seen in prophase I, known as germinal vesicle breakdown (GVBD), and formation of the first meiotic spindle in metaphase I [27]. From there the oocyte resumes meiosis I, asymmetrically divides giving off the first polar body, and enters meiosis II where it is again arrested until fertilization [25, 34].
Metaphase II (MII) arrest depends on the “cytostatic factor” (CSF) activity referring to not a single molecule or gene, but a series of mechanisms that arrest the oocyte in metaphase [35, 36]. Notably, the maturation-promoting factor (MPF), a heterodimer made up of cyclin B1 and CDK1, sustains MII arrest. MPF activity is eventually inactivated through the anaphase-promoting complex/cyclosome (APC/C) that facilitates the degradation of cyclin B1 through ubiquitin-mediated proteolysis [35]. Additionally, Wee1, a tyrosine kinase, destabilizes MPF through phosphorylation of Cdk1, triggering the dissociation of cyclin B1 that allows for degradation by APC/C [36]. APC/C activity is inhibited by early meiotic-inhibitor 1-related protein 1 (Emi2), a zinc finger protein that was first identified in xenopus laevis oocytes [26, 35, 37]. At its zinc-binding region (ZBR), Emi2 binds APC/C thereby inhibiting it from degrading MPF keeping the oocyte arrested. Emi2 therefore preserves CDK activity during the metaphase arrest.
At fertilization, the oocyte undergoes egg activation triggered by the spermatozoa, followed by an observed rapid efflux of zinc ions, termed “zinc sparks,” regulated by calcium oscillations are observed in oocytes from several species including mouse, bovine, human, and even in the roundworm Caenorhabditis elegans and are associated with progression of the cell cycle [38–44]. The release of zinc from the oocyte is mediated by exocytosis of zinc on loaded vesicles containing around 106 zinc atoms [45]. Emi2 is degraded through the rise in intracellular calcium (Ca2+) that activates calmodulin kinase II (CaMKII) to phosphorylate Emi2, destabilizing the complex and releasing APC/C [37]. The now activated APC/C then is free to promote anaphase by degrading many substrates including cyclin B1, decreasing MPF activity and allowing the oocyte to complete the meiotic division [36]. Completion of meiosis involves sequential events including extrusion of a second polar body, formation of pronuclei (male and female), and the transition to mitotic embryonic divisions [36].
The Importance of Zinc
The process of meiotic maturation from prophase I to MII (after meiotic resumption) in the mouse model occurs in about 12–14 h, with the total cellular content of zinc increasing by more than 50% [26, 46, 47]. The oocyte remains arrested in prophase I in the antral follicle due in part to high cAMP/PKA preventing MPF activity, and, in vitro, oocytes will mature spontaneously without treatment with a PDE3A inhibitor. It was found in a study by Tian and Diaz that treatment of mouse oocytes with a zinc-specific chelator, TPEN, resulted in GVBD even in the presence of a PDE3A inhibitor in more than 90% of oocytes treated [48]. Moreover, another study found that chelation of zinc at prophase I resulted in premature arrest in telophase I, signifying the importance of zinc as an inorganic signal for oocyte maturation [38]. Recently, our lab has shown that zinc decreases in the oocyte as a function of increasing maternal age, which is correlated with decreasing oocyte quality (Camp et. al., in press). In this study, we reported that MII oocytes from young mice contained around 10.05 picograms (pg) of zinc/oocyte with around 100% good quality oocytes while old animals on average had around 5.82 pg of zinc/oocyte and produced no good quality oocytes as shown through cortical granule status and microtubule dynamics. Similarly, a study by Kim et al. investigated intraoocyte zinc concentrations required for meiotic progression and found MII mouse oocytes have approximately 6.0 × 1010 atoms of Zn2+/oocyte, which is equivalent to a concentration of 6.5 pg/oocyte [46]. The authors of this study determined zinc-insufficient oocytes treated with TPEN all showed the spindle in a telophase configuration signifying an unusual block in meiosis [46].
The rise in intracellular zinc and meiotic progression are dependent on maternally derived ZIP6 and ZIP10 genes that encode SLC39 family zinc transporters that import zinc ions across the plasma or luminal membrane into the cytosol [49]. At fertilization, the zinc spark begins with thousands of cortically enriched, zinc-loaded secretory compartments releasing around 15% of the oocytes total zinc content at activation [45]. At the time of activation, these vesicles undergo dynamic movement allowing for the rapid exocytosis of zinc. Recently discovered phenomenon of zinc sparks or zinc exocytosis at fertilization also results in about 300% increase in zinc in the zona pellucida and the resultant “Zn2+ shield” prevents penetration of supernumerary spermatozoa [50]. Tian and Diaz demonstrated that acute zinc deficiency in mice before conception led to a dramatic disruption of oocyte and global DNA methylation and decreased in transcription of critical factors involved in oocyte development such as Gdf9, ZP3, and Figla [51]. On the other hand, Jeon et al. showed improved development of preimplantation embryos and parthenotes derived from in vitro matured oocytes supplemented with zinc during maturation [52]. This was accompanied with increased intracellular GSH and transcription factor expression concomitant with decreased ROS [52]. Additionally, Zn2+ can also “decapacitate” and prevent additional spermatozoa from undergoing hyperactivation, and thus prevent polyspermy. The spatiotemporal characteristics of fertilization-induced zinc spark profile is also recently proposed as parameter for the assessment of oocyte/embryo quality, as the zinc spark profile was positively correlated with total number of cells in the resulting morulae and blastocysts [50].
Nitric Oxide, Zinc, and the Oocyte
Virtually every step of oocyte development involves nitric oxide (NO), including meiotic maturation, fertilization, embryonic cleavage, and implantation, as biological concentrations are important for maintaining oocyte quality and delaying oocyte aging [53–57]. NO is generated through NOS which requires both Zn2+ and Ca2+ to regulate activity [58, 59]. Zinc is required for the stability of the enzyme’s active dimeric form in all three isoforms: inducible, iNOS; endothelial, eNOS; and neuronal, nNOS. The zinc atom is coordinated in a tetrahedral arrangement between two Cys-Cys motifs from each monomer [59]. Here, Zn2+ serves a structural function rather than catalytic. NOSs require adequate amounts of the substrate L-arginine (L-Arg) and the cofactor tetrahydrobiopterin (H4B) for heme coupling and O2 reduction in NOS dimers to produce NO and L-citrulline [60]. When these are not present in sufficient amounts, the enzyme utilizes O2 to instead produce O2•−.
NO can improve oocyte quality and delay aging through NO-mediated activation of soluble guanylyl cyclase and cGMP-dependent protein kinase [61, 62]. NO is a ligand of soluble guanylyl cyclase, which catalyzes the conversion of GTP to cGMP; therefore, sufficient amounts of NO are required to allow adequate production of cGMP and maintenance of oocyte arrest [57, 61, 63, 64]. Both ONOO− and HOCl can downregulate NO production through destruction of NOS, as these molecules may target cysteine and histidine residues binding zinc and flavins to NOS [65]. This disturbance of zinc binding to proteins is of particular importance in the context of the developing oocyte, decreasing the production of NO and altering oocyte quality. The loss of NO production at the MII stage will alter the production of cGMP, leading to decreased cAMP levels, dysregulated calcium channels (i.e., InsP3 and RYR), and oocyte activation.
Although there is increased production, NO can be rapidly consumed by O2•− and myeloperoxidase (MPO). MPO, generated in inflammation from activated neutrophils and macrophages, compound I and II can consume NO as a physiological 1e− substrate generating nitrosonium cation that then hydrolyzes into NO2−. Under conditions of enhanced oxidative stress, the activation of inflammatory cytokines results in NO synthesis and subsequent accumulation of several NO and RNS species that can react with dioxygen producing nitrogen dioxide, nitrite, and nitrous acid. The NO intracellular carrier S-nitrosoglutathione (GSNO), formed during transnitrosylation of glutathione (GSH) with NO, can facilitate irreversible damage to tissues when produced at high levels [19, 66]. Also, GSNO can s-nitrosylate the zinc cysteine core, and it has been noted that the mechanism of zinc release under low concentrations of GSNO is reversible [19].
Oxidative Stress and Zinc
The delicate balance between ROS/RNS and antioxidants is important in the maintenance of homeostasis and regulation of physiological signaling. In the event that ROS production is increased contributing to enhanced oxidative stress, the present antioxidant/oxidant ratio is disrupted resulting in an imbalance and potential oxidative damage to the oocyte and its microenvironment. ROS can either directly damage the oocyte or deplete oocyte quality through disruption of cumulus cells, decreasing the ability of the oocyte to mature [67–70]. Cumulus cells synthesize glutathione (GSH) as well as other antioxidants to protect the oocyte against oxidative stress [71, 72]. ROS such as superoxide (O2•−), hydrogen peroxide (H2O2), hypochlorous acid (HOCl), peroxynitrite (ONOO−), and hydroxyl radical (•OH) can disturb the cumulus cells and connexin proteins, leading to a disruption of cellular communication in the cumulus oocyte complex, potentially creating an imbalance in antioxidant machinery and oocyte activation due to depletion of cGMP [29, 68, 73].
Moreover, O2•− produced from overexpression of NADPH oxidase can result in increased production of ONOO−, the product of the reaction between O2•− and NO, that can further cause substantial damage through thiol oxidation, lipid peroxidation, inactivation of enzymes and ion channels via protein oxidation and nitration, and inhibition of mitochondrial respiration [74]. Enhanced NADPH oxidase activity and the subsequent production of ROS has been linked to zinc deficiency [75, 76]. NADPH oxidase generated O2•− overproduction and metabolism results in the activation of caspace-3 contributing to DNA breaks and damage and apoptosis [77]. Several studies have indicated zinc can inhibit caspase activity through binding to the cysteine-histidine catalytic dyad on the caspases [78, 79]. Specifically, zinc at biologically relevant low concentrations, as low as 1.7 nM, have been shown to inhibit caspase-3 activity, which can bind three zinc ions [80, 81]. Additionally, zinc functions to inhibit NADPH oxidases and is a cofactor for SOD which can catalyze the dismutation of O2− to H2O2, thus assisting the antioxidant defense. When H2O2 is generated, it can react with redox-active transition metals through the Fenton reaction producing •OH, which can induce lipid peroxidation events and is the most damaging ROS. Zinc can competitively antagonize the participating redox-active transition metals, such as iron, cobalt, copper, and nickel, and thus may aid in preventing against site-specific peroxidative injury [82–84]. In another way, zinc can protect select proteins and enzymes from oxidation by stabilizing sulfhydryl’s either through direct binding, nearby binding producing steric hindrance, or causing conformational changes to the protein [83]. However, ROS may play an important role in disturbing the binding of Zn2+ to essential proteins and enzymes, leading to decreased oocyte quality.
Zinc-binding Proteins and ROS
As discussed, Emi2 is an important regulator of APC/C and miotic arrest at MII prior to fertilization, with the rise of intracellular calcium brought about by joining of a sperm cell leading to degeneration of Emi2 and thus resumption of meiosis [26, 85]. The structure of Emi2 is known and includes a destruction box (D-Box), zinc-binding region (ZBR), and an RL-tail at the C terminus [85]. A crystal structure of the ZBR fragment was determined by NMR spectroscopy showing the coordination of two zinc ions [85, 86]. The N-terminus contains two successive β-hairpins (β1 and β2 on the first and β3 and β4 on the second). The first zinc ion is coordinated between four cysteine residues (C4) at the tips of the first and second β-hairpins, which are tightly bound forming a scissor-like structure. The second zinc ion is coordinated at the C-terminal which forms a GAG-knuckle zinc-binding site with three cysteine residues and one histidine residue (C3H). The β4 strand associates with a β5 strand in an anti-parallel manner forming the C-terminal [85]. Interestingly, a study by Suzuki et al., investigated the role of zinc in the MII oocyte independent of calcium and found that Zn2+ chelation induces exit from MII with subsequent Cyclin B degradation without degradation of Emi2. A study by Bernhardt et al. determined that zinc insufficiency at the end of MI causes meiotic arrest, decreased CCNB1, and reduced MPF activity and concluded that a reduction in intracellular zinc availability is both necessary and sufficient to cause activation of the MII-arrested oocyte [19, 26]. Another report determined that reducing zinc content in the MII oocyte using the zinc-chelating agent TPEN resulted in release of CGs and spindle depolarization in MII porcine oocytes as well as a significant reduction in MPF activity, evidenced by p34cdc2 activity [87]. Together, these studies suggest the necessity of zinc in oocyte arrest and in Emi2 regulation of the APC/C allowing for meiotic arrest [88].
Zinc binding in zinc finger proteins and enzymes reduces the thiol oxidation potential which can be disturbed by ROS/RNS releasing zinc from the coordination sphere and generating loss of protein function [89]. Previously, our lab has shown that treating oocytes with agents capable of chelating zinc result in a decrease in oocyte zinc with an increase in ROS [22, 24]. Increased ROS can diminish MII oocyte quality either indirectly through disruption to the cumulus cells and connexin proteins or directly through direct disturbance to the oocyte. Moreover, this enhancement in ROS and decrease in zinc may promote mitochondrial damage subsequently increasing NADPH oxidase activity and the production of O2•−. O2•− can then react with existing NO to generate ONOO−; however, extracellularly generated ONOO− can freely diffuse through the phospholipid membrane, as the calculated permeability constant of ONOO− is ≈8.0 × 10−4 cm⋅s−1, comparable to that of water [90]. ONOO− may induce the destruction of zinc-binding domains through S-nitrosylation resulting in zinc dissociation. HOCl is produced during inflammation when MPO reacts with H2O2 in the presence of Cl− [91], and can freely diffuse through the cell membrane facilitating oocyte deterioration and aging as evidenced by significant increases in zona pellucida dissolution time, altered spindle and chromosomal alignment, and cortical granule loss with the potential to oxidize zinc-binding domains [70, 92]. This resultant loss of function of zinc-binding proteins due to oxidation of the cysteinyl thiol residues, resulting in formation of a S–S disulfide bridge, may be reversible in the presence of a reducing agent [19]. Thus, the ability of ROS to disturb zinc binding essentially chelates zinc from the zinc-binding domain, rendering the protein or enzyme dysfunctional. This may mimic the results seen when using zinc-specific chelators, as described above, resulting in loss of MII arrest and oocyte aging phenomena.
The Potential Role of Zinc and ROS in Endometriosis-associated Infertility
The role of zinc and the oxidant to antioxidant balance may serve as guidance for understanding inflammatory diseases and related sub-fertility. For example, the discussed interaction of oxidative stress and zinc can be applied to endometriosis. Endometriosis, an enigmatic disorder of the female reproductive tract, includes several biologically active proinflammatory mediators [8, 93]. Of these mediators, nuclear factor kappa B (NF-κB) has been noted to play a substantial role in endometriosis [94–97], as its signaling is activated in endometriotic cells by tumor necrosis factor α (TNF-α) and interleukin-1β (IL-1β), also noted to be prominent in endometriosis [98–100]. NF-κB activates transcriptional activity of proinflammatory cytokines/chemokines including IL1, IL,6, IL8, and TNF-α [95, 100–103]. Interestingly, it has been reported that inflammatory cytokines including TNF-α and IL-1β are associated with increased ROS and decreased zinc [104–106], while zinc has been shown to inhibit NF-κB activation in several different disorders [107–110]. Thus, the investigation of zinc in the inflammatory response of endometriosis as well as endometriotic cell proliferation and development may be of increasing interest.
Recently, we found immature oocytes from women with endometriosis had decreased maturation competence, increased cortical granule loss, zona pellucida hardening, and spindle/chromosome disruption signifying a decrease in oocyte quality [111]. The mechanistic action of endometriosis in alerting oocyte quality can be determined by examining the role of ROS on cumulus cells, NO, and zinc in the oocyte. First, we previously observed cumulus cells from patients with endometriosis presented an increased incidence of apoptosis and high protein nitration as evidence by high measured NO2− [111]. Second, in isolated oocytes without cumulus cells, there is high protein nitration, observed premature cortical granule loss accompanied by alteration in oocyte size, shape, and microtubule spindle formation [111]. These phenomena can be attributed to the above presented model, as studies have found alterations in mitochondrial function and cumulus cells [112]. Disruptions to the COC by ROS can alter the cGMP and cAMP levels contributing to disturbed maintenance of meiotic arrest explaining the zona pellucida hardening, granulosa cell apoptosis, and spindle/chromosome disruptions. Additionally, ROS-mediated destruction of NOSs can decrease the liable NO used in the guanylyl cyclase system, producing similar results as evidenced by our previous work reporting NO supplementation can delay oocyte aging [57, 61, 62]. In regard to the isolated oocyte, decreased zinc can generate an influx of Ca2+ into the oocyte activating PKC and increasing NADPH oxidase expression. In turn, this will increase the production of ONOO− explaining the increased protein nitration [113]. Furthermore, this occurrence is supported by the finding of increased abnormal mitochondria and decreased mitochondrial mass in oocytes from patients with endometriosis [114].
The elevated production of ROS in endometriosis, independent of the calcium mediated mechanisms above, may also disturb zinc-binding regions of Emi2 and other zinc finger proteins in the oocyte explaining the disruption to the pericentrin, spindles, and chromosomal alignment. Thus, the effects of ROS on oocytes may be acting through removing zinc from the ZBR of Emi2, thereby inhibiting its functionality, releasing APC/C, and allowing for meiotic resumption.
Conclusion
The functional role of zinc as an antioxidant may be key to understanding oxidative stress in inflammatory diseases that impact fertility, with insufficiencies potentially resulting in dysfunctional antioxidant enzymes and increased ROS accumulation. The overproduction of ROS in this way may function to disrupt cumulus cell functioning, deplete NO production, and/or remove the bound zinc of critical zinc finger proteins such as Emi2 or those present in the microtubule organizing center producing similar effects as zinc-chelating agents, i.e., anomalous pericentrin formation, disappearance of spindle fibers, altered chromosomal alignment, and subsequent meiotic resumption without fertilization [22, 115, 116]. More research is needed to investigate the potential of zinc in improving the antioxidant defense and oocyte quality.
Abbreviations
- LH
Luteinizing hormone
- ZFP
Zinc finger protein
- ROS
Reactive oxygen species
- NO
Nitric oxide
- iNOS
Inducible nitric oxide synthase
- HOCl
Hypochlorous acid
- O2•−
Superoxide
- H2O2
Hydrogen peroxide
- •OH
Hydroxyl radical
- ONOO−
Peroxynitrite
- Cl−
Chloride
- H4B
Tetrahydrobiopterin
- CAT
Catalase
- RNS
Reactive nitrogen species
Footnotes
Conflicts of Interest The authors declare no competing interests.
References
- 1.Garner TB, Hester JM, Carothers A, Diaz FJ. Role of zinc in female reproduction. Biol Reprod. 2021;104(5):976–94. 10.1093/biolre/ioab023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.McClung JP. Iron, zinc, and physical performance. Biol Trace Elem Res. 2019;188(1):135–9. 10.1007/s12011-018-1479-7. [DOI] [PubMed] [Google Scholar]
- 3.Fallah A, Mohammad-Hasani A, Colagar AH. Zinc is an essential element for male fertility: a review of Zn roles in men’s health, germination, sperm quality, and fertilization. J Reprod Infertil. 2018;19(2):69–81. [PMC free article] [PubMed] [Google Scholar]
- 4.Sanna A, Firinu D, Zavattari P, Valera P. Zinc status and auto-immunity: a systematic review and meta-analysis. Nutrients. 2018;10(1):68 10.3390/nu10010068. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Skalny AV, Aschner M, Tinkov AA. Zinc. Adv Food Nutr Res. 2021;96:251–310. 10.1016/bs.afnr.2021.01.003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Li J, Chen H, Gou M, Tian C, Wang H, Song X, et al. Molecular features of polycystic ovary syndrome revealed by transcriptome analysis of oocytes and cumulus cells. Front Cell Dev Biol. 2021;9:735684. 10.3389/fcell.2021.735684. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Qiao J, Feng HL. Extra- and intra-ovarian factors in polycystic ovary syndrome: impact on oocyte maturation and embryo developmental competence. Hum Reprod Update. 2011;17(1):17–33. 10.1093/humupd/dmq032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Camp OG, Bai D, Goud PT, Diamond MP, Abu-Soud HM. A novel theory implicating hypochlorous acid as the primary generator of angiogenesis, infertility, and free iron in endometriosis. F&S Reviews. 2022;3(2):146–56. 10.1016/j.xfnr.2022.02.001. [DOI] [Google Scholar]
- 9.Wessels I, Maywald M, Rink L. Zinc as a gatekeeper of immune function. Nutrients. 2017;9(12):1286. 10.3390/nu9121286. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Hennigar SR, Lieberman HR, Fulgoni VL 3rd, McClung JP. Serum zinc concentrations in the US population are related to sex, age, and time of blood draw but not dietary or supplemental zinc. J Nutr. 2018;148(8):1341–51. 10.1093/jn/nxy105. [DOI] [PubMed] [Google Scholar]
- 11.Hennigar SR, Kelley AM, McClung JP. Metallothionein and zinc transporter expression in circulating human blood cells as biomarkers of zinc status: a systematic review. Adv Nutr. 2016;7(4):735–46. 10.3945/an.116.012518. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Eide DJ. The oxidative stress of zinc deficiency. Metallomics. 2011;3(11):1124–9. 10.1039/c1mt00064k. [DOI] [PubMed] [Google Scholar]
- 13.Kloubert V, Rink L. Zinc as a micronutrient and its preventive role of oxidative damage in cells. Food & Function. 2015;6(10):3195–204. Epub 2015/08/20. 10.1039/c5fo00630a. [DOI] [PubMed] [Google Scholar]
- 14.Lee SR. Critical role of zinc as either an antioxidant or a prooxidant in cellular systems. Oxid Med Cell Longev. 2018;2018:9156285. 10.1155/2018/9156285. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Sethuram R, Bai D, Abu-Soud HM. Potential role of zinc in the COVID-19 disease process and its probable impact on reproduction. Reprod Sci. 2022;29(1):1–6. 10.1007/s43032-020-00400-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Cassandri M, Smirnov A, Novelli F, Pitolli C, Agostini M, Malewicz M, et al. Zinc-finger proteins in health and disease. Cell Death Discov. 2017;3:17071. 10.1038/cddiscovery.2017.71. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Krishna SS, Majumdar I, Grishin NV. Structural classification of zinc fingers: survey and summary. Nucleic Acids Res. 2003;31(2):532–50. 10.1093/nar/gkg161. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Miloch A, Krezel A. Metal binding properties of the zinc finger metallome–insights into variations in stability. Metallomics. 2014;6(11):2015–24. 10.1039/c4mt00149d. [DOI] [PubMed] [Google Scholar]
- 19.Kluska K, Adamczyk J, Krężel A. Metal binding properties, stability and reactivity of zinc fingers. Coord Chem Rev. 2018;367:18–64. 10.1016/j.ccr.2018.04.009. [DOI] [Google Scholar]
- 20.Colvin RA, Holmes WR, Fontaine CP, Maret W. Cytosolic zinc buffering and muffling: their role in intracellular zinc homeostasis. Metallomics. 2010;2(5):306–17. 10.1039/b926662c. [DOI] [PubMed] [Google Scholar]
- 21.Singh AK, Chattopadhyay R, Chakravarty B, Chaudhury K. Markers of oxidative stress in follicular fluid of women with endometriosis and tubal infertility undergoing IVF. Reprod Toxicol. 2013;42:116–24. 10.1016/j.reprotox.2013.08.005. [DOI] [PubMed] [Google Scholar]
- 22.Yahfoufi ZA, Bai D, Khan SN, Chatzicharalampous C, Kohan-Ghadr HR, Morris RT, et al. Glyphosate induces metaphase II oocyte deterioration and embryo damage by zinc depletion and overproduction of reactive oxygen species. Toxicology. 2020;439:152466. 10.1016/j.tox.2020.152466. [DOI] [PubMed] [Google Scholar]
- 23.Nikbakht R, Mohammadjafari R, Rajabalipour M, Moghadam MT. Evaluation of oocyte quality in Polycystic ovary syndrome patients undergoing ART cycles. Fertil Res Pract. 2021;7(1):2. 10.1186/s40738-020-00094-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Aldhaheri SR, Jeelani R, Kohan-Ghadr HR, Khan SN, Mikhael S, Washington C, et al. Dimercapto-1-propanesulfonic acid (DMPS) induces metaphase II mouse oocyte deterioration. Free Radic Biol Med. 2017;112:445–51. 10.1016/j.freeradbiomed.2017.08.015. [DOI] [PubMed] [Google Scholar]
- 25.He M, Zhang T, Yang Y, Wang C. Mechanisms of oocyte maturation and related epigenetic regulation. Front Cell Dev Biol. 2021;9:654028. 10.3389/fcell.2021.654028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Bernhardt ML, Kong BY, Kim AM, O’Halloran TV, Woodruff TK. A zinc-dependent mechanism regulates meiotic progression in mammalian oocytes. Biol Reprod. 2012;86(4):114. 10.1095/biolreprod.111.097253. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Mehlmann LM. Stops and starts in mammalian oocytes: recent advances in understanding the regulation of meiotic arrest and oocyte maturation. Reproduction. 2005;130(6):791–9. 10.1530/rep.1.00793. [DOI] [PubMed] [Google Scholar]
- 28.Gershon E, Maimon I, Galiani D, Elbaz M, Karasenti S, Dekel N. High cGMP and low PDE3A activity are associated with oocyte meiotic incompetence. Cell Cycle. 2019;18(20):2629–40. 10.1080/15384101.2019.1652472. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Shuhaibar LC, Egbert JR, Norris RP, Lampe PD, Nikolaev VO, Thunemann M, et al. Intercellular signaling via cyclic GMP diffusion through gap junctions restarts meiosis in mouse ovarian follicles. Proc Natl Acad Sci U S A. 2015;112(17):5527–32. 10.1073/pnas.1423598112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Granot I, Dekel N. Phosphorylation and expression of connexin-43 ovarian gap junction protein are regulated by luteinizing hormone. J Biol Chem. 1994;269(48):30502–9. [PubMed] [Google Scholar]
- 31.Norris RP, Freudzon M, Mehlmann LM, Cowan AE, Simon AM, Paul DL, et al. Luteinizing hormone causes MAP kinase-dependent phosphorylation and closure of connexin 43 gap junctions in mouse ovarian follicles: one of two paths to meiotic resumption. Development. 2008;135(19):3229–38. 10.1242/dev.025494. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Zhang W, Colman RW. Conserved amino acids in metal-binding motifs of PDE3A are involved in substrate and inhibitor binding. Blood. 2000;95(11):3380–6. [PubMed] [Google Scholar]
- 33.Vaccari S, Weeks JL 2nd, Hsieh M, Menniti FS, Conti M. Cyclic GMP signaling is involved in the luteinizing hormone-dependent meiotic maturation of mouse oocytes. Biol Reprod. 2009;81(3):595–604. 10.1095/biolreprod.109.077768. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Fan H-Y, Sun Q-Y. Chapter 12 Oocyte meiotic maturation. Ovary. 2019:181–203. 10.1016/B978-0-12-813209-8.00012-1 [DOI] [Google Scholar]
- 35.Tripathi A, Kumar KV, Chaube SK. Meiotic cell cycle arrest in mammalian oocytes. J Cell Physiol. 2010;223(3):592–600. 10.1002/jcp.22108. [DOI] [PubMed] [Google Scholar]
- 36.Prasad S, Tiwari M, Koch B, Chaube SK. Morphological, cellular and molecular changes during postovulatory egg aging in mammals. J Biomed Sci. 2015;22:36. 10.1186/s12929-015-0143-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Madgwick S, Jones KT. How eggs arrest at metaphase II: MPF stabilisation plus APC/C inhibition equals Cytostatic Factor. Cell Div. 2007;2:4. 10.1186/1747-1028-2-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Kim AM, Bernhardt ML, Kong BY, Ahn RW, Vogt S, Woodruff TK, et al. Zinc sparks are triggered by fertilization and facilitate cell cycle resumption in mammalian eggs. ACS Chem Biol. 2011;6(7):716–23. 10.1021/cb200084y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Wu JM, Zelinski MB, Ingram DK, Ottinger MA. Ovarian aging and menopause: current theories, hypotheses, and research models. Exp Biol Med (Maywood). 2005;230(11):818–28. 10.1177/153537020523001106. [DOI] [PubMed] [Google Scholar]
- 40.Xi X, Zou Q, Wei Y, Chen Y, Wang X, Lv D, et al. Dynamic changes of DNA methylation and transcriptome expression in porcine ovaries during aging. BioMed Res Int. 2019;2019:8732023. 10.1155/2019/8732023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Malhi PS, Adams GP, Mapletoft RJ, Singh J. Superovulatory response in a bovine model of reproductive aging. Anim Reprod Sci. 2008;109(1–4):100–9. 10.1016/j.anireprosci.2007.12.002. [DOI] [PubMed] [Google Scholar]
- 42.Uliani RC, Conley AJ, Corbin CJ, Friso AM, Maciel LFS, Alvarenga MA. Anti-Müllerian hormone and ovarian aging in mares. J Endocrinol. 2019;240(2):147–56. 10.1530/joe-18-0391. [DOI] [PubMed] [Google Scholar]
- 43.Nichols SM, Bavister BD, Brenner CA, Didier PJ, Harrison RM, Kubisch HM. Ovarian senescence in the rhesus monkey (Macaca mulatta). Hum Reprod (Oxford, England). 2005;20(1):79–83. 10.1093/humrep/deh576. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Mendoza AD, Sue A, Antipova O, Vogt S, Woodruff TK, Wig-nall SM, et al. Dynamic zinc fluxes regulate meiotic progression in Caenorhabditis elegans†. Biol Reprod. 2022;107(2):406–18. 10.1093/biolre/ioac064. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Que EL, Bleher R, Duncan FE, Kong BY, Gleber SC, Vogt S, et al. Quantitative mapping of zinc fluxes in the mammalian egg reveals the origin of fertilization-induced zinc sparks. Nat Chem. 2015;7(2):130–9. 10.1038/nchem.2133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Kim AM, Vogt S, O’Halloran TV, Woodruff TK. Zinc availability regulates exit from meiosis in maturing mammalian oocytes. Nat Chem Biol. 2010;6(9):674–81. 10.1038/nchembio.419. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Celik O, Celik N, Gungor S, Haberal ET, Aydin S. Selective regulation of oocyte meiotic events enhances progress in fertility preservation methods. Biochem Insights. 2015;8:11–21. 10.4137/BCI.S28596. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Tian X, Diaz FJ. Zinc depletion causes multiple defects in ovarian function during the periovulatory period in mice. Endocrinology. 2012;153(2):873–86. 10.1210/en.2011-1599. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Kong BY, Duncan FE, Que EL, Kim AM, O’Halloran TV, Woodruff TK. Maternally-derived zinc transporters ZIP6 and ZIP10 drive the mammalian oocyte-to-egg transition. Mol Hum Reprod. 2014;20(11):1077–89. 10.1093/molehr/gau066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Zhang N, Duncan FE, Que EL, O’Halloran TV, Woodruff TK. The fertilization-induced zinc spark is a novel biomarker of mouse embryo quality and early development. Sci Rep. 2016;6:22772. 10.1038/srep22772. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Tian X, Diaz FJ. Acute dietary zinc deficiency before conception compromises oocyte epigenetic programming and disrupts embryonic development. Dev Biol. 2013;376(1):51–61. 10.1016/j.ydbio.2013.01.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Jeon Y, Yoon JD, Cai L, Hwang SU, Kim E, Zheng Z, et al. Supplementation of zinc on oocyte in vitro maturation improves preimplatation embryonic development in pigs. Theriogenology. 2014;82(6):866–74. 10.1016/j.theriogenology.2014.06.021. [DOI] [PubMed] [Google Scholar]
- 53.Jablonka-Shariff A, Olson LM. The role of nitric oxide in oocyte meiotic maturation and ovulation: meiotic abnormalities of endothelial nitric oxide synthase knock-out mouse oocytes. Endocrinology. 1998;139(6):2944–54. 10.1210/endo.139.6.6054. [DOI] [PubMed] [Google Scholar]
- 54.Tranguch S, Steuerwald N, Huet-Hudson YM. Nitric oxide synthase production and nitric oxide regulation of preimplantation embryo development. Biol Reprod. 2003;68(5):1538–44. 10.1095/biolreprod.102.009282. [DOI] [PubMed] [Google Scholar]
- 55.Khorram O Nitric oxide and its role in blastocyst implantation. Rev Endocr Metab Disord. 2002;3(2):145–9. 10.1023/a:1015459029397. [DOI] [PubMed] [Google Scholar]
- 56.Nishikimi A, Matsukawa T, Hoshino K, Ikeda S, Kira Y, Sato EF, et al. Localization of nitric oxide synthase activity in unfertilized oocytes and fertilized embryos during preimplantation development in mice. Reproduction. 2001;122(6):957–63. 10.1530/rep.0.1220957. [DOI] [PubMed] [Google Scholar]
- 57.Goud AP, Goud PT, Diamond MP, Abu-Soud HM. Nitric oxide delays oocyte aging. Biochemistry. 2005;44(34):11361–8. 10.1021/bi050711f. [DOI] [PubMed] [Google Scholar]
- 58.Abu-Soud HM, Stuehr DJ. Nitric oxide synthases reveal a role for calmodulin in controlling electron transfer. Proc Natl Acad Sci U S A. 1993;90(22):10769–72. 10.1073/pnas.90.22.10769. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Raman CS, Li H, Martasek P, Kral V, Masters BS, Poulos TL. Crystal structure of constitutive endothelial nitric oxide synthase: a paradigm for pterin function involving a novel metal center. Cell. 1998;95(7):939–50. 10.1016/s0092-8674(00)81718-3. [DOI] [PubMed] [Google Scholar]
- 60.Stuehr DJ. Mammalian nitric oxide synthases. Biochim Biophys Acta. 1999;1411(2–3):217–30. 10.1016/s0005-2728(99)00016-x. [DOI] [PubMed] [Google Scholar]
- 61.Goud AP, Goud PT, Diamond MP, Gonik B, Abu-Soud HM. Activation of the cGMP signaling pathway is essential in delaying oocyte aging in diabetes mellitus. Biochemistry. 2006;45(38):11366–78. 10.1021/bi060910e. [DOI] [PubMed] [Google Scholar]
- 62.Goud PT, Goud AP, Diamond MP, Gonik B, Abu-Soud HM. Nitric oxide extends the oocyte temporal window for optimal fertilization. Free Radic Biol Med. 2008;45(4):453–9. 10.1016/j.freeradbiomed.2008.04.035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Ignarro LJ. Haem-dependent activation of guanylate cyclase and cyclic GMP formation by endogenous nitric oxide: a unique transduction mechanism for transcellular signaling. Pharmacol Toxicol. 1990;67(1):1–7. 10.1111/j.1600-0773.1990.tb00772.x. [DOI] [PubMed] [Google Scholar]
- 64.Denninger JW, Marletta MA. Guanylate cyclase and the .NO/cGMP signaling pathway. Biochim Biophys Acta. 1999;1411(2–3):334–50. 10.1016/s0005-2728(99)00024-9. [DOI] [PubMed] [Google Scholar]
- 65.Camp OG, Bai D, Awonuga A, Goud PT, Abu-Soud HM. Hypochlorous acid facilitates inducible nitric oxide synthase sub-unit dissociation: the link between heme destruction, disturbance of the zinc-tetrathiolate center, and the prevention by melatonin. Nitric Oxide. 2022;124:32–8. 10.1016/j.niox.2022.04.006. [DOI] [PubMed] [Google Scholar]
- 66.Foster MW, Hess DT, Stamler JS. Protein S-nitrosylation in health and disease: a current perspective. Trends Mol Med. 2009;15(9):391–404. 10.1016/j.molmed.2009.06.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Banerjee J, Shaeib F, Maitra D, Saed GM, Dai J, Diamond MP, et al. Peroxynitrite affects the cumulus cell defense of metaphase II mouse oocytes leading to disruption of the spindle structure in vitro. Fertil Steril. 2013;100(2):578–84.e1. 10.1016/j.fertnstert.2013.04.030. [DOI] [PubMed] [Google Scholar]
- 68.Shaeib F, Khan SN, Ali I, Thakur M, Saed MG, Dai J, et al. The defensive role of cumulus cells against reactive oxygen species insult in metaphase II mouse oocytes. Reprod Sci. 2016;23(4):498–507. 10.1177/1933719115607993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Thakur M, Shaeib F, Khan SN, Kohan-Ghadr HR, Jeelani R, Aldhaheri SR, et al. Galactose and its metabolites deteriorate metaphase II mouse oocyte quality and subsequent embryo development by disrupting the spindle structure. Sci Rep. 2017;7(1):231. 10.1038/s41598-017-00159-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Goud AP, Goud PT, Diamond MP, Gonik B, Abu-Soud HM. Reactive oxygen species and oocyte aging: role of superoxide, hydrogen peroxide, and hypochlorous acid. Free Radic Biol Med. 2008;44(7):1295–304. 10.1016/j.freeradbiomed.2007.11.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Cetica PD, Pintos LN, Dalvit GC, Beconi MT. Antioxidant enzyme activity and oxidative stress in bovine oocyte in vitro maturation. IUBMB Life. 2001;51(1):57–64. 10.1080/15216540119253. [DOI] [PubMed] [Google Scholar]
- 72.Cetica PD, Pintos LN, Dalvit GC, Beconi MT. Effect of lactate dehydrogenase activity and isoenzyme localization in bovine oocytes and utilization of oxidative substrates on in vitro maturation. Theriogenology. 1999;51(3):541–50. 10.1016/s0093-691x(99)00008-4. [DOI] [PubMed] [Google Scholar]
- 73.Ferre-Pujol P, Nguyen XK, Nagahara T, Bui TTM, Wakai T, Funahashi H. Removal of cumulus cells around 20 h after the start of in vitro maturation improves the meiotic competence of porcine oocytes via reduction in cAMP and cGMP levels. J Reprod Dev. 2019;65(2):177–82. 10.1262/jrd.2018-130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Beckman JS, Chen J, Ischiropoulos H, Crow JP. Oxidative chemistry of peroxynitrite. Methods Enzymol. 1994;233:229–40. 10.1016/s0076-6879(94)33026-3. [DOI] [PubMed] [Google Scholar]
- 75.Li MS, Adesina SE, Ellis CL, Gooch JL, Hoover RS, Williams CR. NADPH oxidase-2 mediates zinc deficiency-induced oxidative stress and kidney damage. Am J Physiol Cell Physiol. 2017;312(1):C47–55. 10.1152/ajpcell.00208.2016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Olechnowicz J, Tinkov A, Skalny A, Suliburska J. Zinc status is associated with inflammation, oxidative stress, lipid, and glucose metabolism. J Physiol Sci. 2018;68(1):19–31. 10.1007/s12576-017-0571-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Goud PT, Goud AP, Najafi T, Gonik B, Diamond MP, Saed GM, et al. Direct real-time measurement of intra-oocyte nitric oxide concentration in vivo. PLoS One. 2014;9(6):e98720. 10.1371/journal.pone.0098720. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Huber KL, Hardy JA. Mechanism of zinc-mediated inhibition of caspase-9. Protein Sci. 2012;21(7):1056–65. 10.1002/pro.2090. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Velazquez-Delgado EM, Hardy JA. Zinc-mediated allosteric inhibition of caspase-6. J Biol Chem. 2012;287(43):36000–11. 10.1074/jbc.M112.397752. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Maret W, Jacob C, Vallee BL, Fischer EH. Inhibitory sites in enzymes: zinc removal and reactivation by thionein. Proc Natl Acad Sci U S A. 1999;96(5):1936–40. 10.1073/pnas.96.5.1936. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Eron SJ, MacPherson DJ, Dagbay KB, Hardy JA. Multiple mechanisms of zinc-mediated inhibition for the apoptotic caspases-3, -6, -7, and -8. ACS Chem Biol. 2018;13(5):1279–90. 10.1021/acschembio.8b00064. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Searle AJF, Tomasi A. Hydroxyl free radical production in iron-cysteine solutions and protection by zinc. J Inorg Biochem. 1982;17(2):161–6. 10.1016/S0162-0134(00)80085-9. [DOI] [Google Scholar]
- 83.Powell SR. The antioxidant properties of zinc. J Nutr. 2000;130(5S Suppl):1447S–S1454. 10.1093/jn/130.5.1447S. [DOI] [PubMed] [Google Scholar]
- 84.Girotti AW, Thomas JP, Jordan JE. Inhibitory effect of zinc(II) on free radical lipid peroxidation in erythrocyte membranes. J Free Radic Biol Med. 1985;1(5–6):395–401. 10.1016/0748-5514(85)90152-7. [DOI] [PubMed] [Google Scholar]
- 85.Shoji S, Muto Y, Ikeda M, He F, Tsuda K, Ohsawa N, et al. The zinc-binding region (ZBR) fragment of Emi2 can inhibit APC/C by targeting its association with the coactivator Cdc20 and UBE2C-mediated ubiquitylation. FEBS Open Bio. 2014;4:689–703. 10.1016/j.fob.2014.06.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Jia JL, Han YH, Kim HC, Ahn M, Kwon JW, Luo Y, et al. Structural basis for recognition of Emi2 by Polo-like kinase 1 and development of peptidomimetics blocking oocyte maturation and fertilization. Sci Rep. 2015;5:14626. 10.1038/srep14626. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Zhao MH, Kim NH, Cui XS. Zinc depletion activates porcine metaphase II oocytes independently of the protein kinase C pathway. In Vitro Cell Dev Biol Anim. 2014;50(10):945–51. 10.1007/s11626-014-9784-8. [DOI] [PubMed] [Google Scholar]
- 88.Suzuki T, Yoshida N, Suzuki E, Okuda E, Perry AC. Full-term mouse development by abolishing Zn2+-dependent metaphase II arrest without Ca2+ release. Development. 2010;137(16):2659–69. 10.1242/dev.049791. [DOI] [PubMed] [Google Scholar]
- 89.Hubner C, Haase H. Interactions of zinc- and redox-signaling pathways. Redox Biol. 2021;41:101916. 10.1016/j.redox.2021.101916. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Marla SS, Lee J, Groves JT. Peroxynitrite rapidly permeates phospholipid membranes. Proc Natl Acad Sci U S A. 1997;94(26):14243–8. 10.1073/pnas.94.26.14243. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Podrez EA, Abu-Soud HM, Hazen SL. Myeloperoxidase-generated oxidants and atherosclerosis. Free Radic Biol Med. 2000;28(12):1717–25. 10.1016/s0891-5849(00)00229-x. [DOI] [PubMed] [Google Scholar]
- 92.Banerjee J, Maitra D, Diamond MP, Abu-Soud HM. Melatonin prevents hypochlorous acid-induced alterations in microtubule and chromosomal structure in metaphase-II mouse oocytes. J Pineal Res. 2012;53(2):122–8. 10.1111/j.1600-079X.2012.00977.x. [DOI] [PubMed] [Google Scholar]
- 93.Zondervan KT, Becker CM, Koga K, Missmer SA, Taylor RN, Vigano P. Endometriosis Nat Rev Dis Primers. 2018;4(1):9. 10.1038/s41572-018-0008-5. [DOI] [PubMed] [Google Scholar]
- 94.Alvarado-Diaz CP, Nunez MT, Devoto L, Gonzalez-Ramos R. Iron overload-modulated nuclear factor kappa-B activation in human endometrial stromal cells as a mechanism postulated in endometriosis pathogenesis. Fertil Steril. 2015;103(2):439–47. 10.1016/j.fertnstert.2014.10.046. [DOI] [PubMed] [Google Scholar]
- 95.Xiu-li W, Su-ping H, Hui-hua D, Zhi-xue Y, Shi-long F, Pin-hong L. NF-kappaB decoy oligonucleotides suppress RANTES expression and monocyte chemotactic activity via NF-kappaB inactivation in stromal cells of ectopic endometrium. J Clin Immunol. 2009;29(3):387–95. 10.1007/s10875-009-9274-z. [DOI] [PubMed] [Google Scholar]
- 96.Taniguchi F, Harada T, Miyakoda H, Iwabe T, Deura I, Tagashira Y, et al. TAK1 activation for cytokine synthesis and proliferation of endometriotic cells. Mol Cell Endocrinol. 2009;307(1–2):196–204. 10.1016/j.mce.2009.04.012. [DOI] [PubMed] [Google Scholar]
- 97.Kim KH, Lee EN, Park JK, Lee JR, Kim JH, Choi HJ, et al. Curcumin attenuates TNF-alpha-induced expression of intercellular adhesion molecule-1, vascular cell adhesion molecule-1 and proinflammatory cytokines in human endometriotic stromal cells. Phytother Res. 2012;26(7):1037–47. 10.1002/ptr.3694. [DOI] [PubMed] [Google Scholar]
- 98.Veillat V, Lavoie CH, Metz CN, Roger T, Labelle Y, Akoum A. Involvement of nuclear factor-kappaB in macrophage migration inhibitory factor gene transcription up-regulation induced by interleukin-1 beta in ectopic endometrial cells. Fertil Steril. 2009;91(5 Suppl):2148–56. 10.1016/j.fertnstert.2008.05.017. [DOI] [PubMed] [Google Scholar]
- 99.Cao WG, Morin M, Metz C, Maheux R, Akoum A. Stimulation of macrophage migration inhibitory factor expression in endometrial stromal cells by interleukin 1, beta involving the nuclear transcription factor NFkappaB. Biol Reprod. 2005;73(3):565–70. 10.1095/biolreprod.104.038331. [DOI] [PubMed] [Google Scholar]
- 100.Ohama Y, Harada T, Iwabe T, Taniguchi F, Takenaka Y, Terakawa N. Peroxisome proliferator-activated receptor-gamma ligand reduced tumor necrosis factor-alpha-induced interleukin-8 production and growth in endometriotic stromal cells. Fertil Steril. 2008;89(2):311–7. 10.1016/j.fertnstert.2007.03.061. [DOI] [PubMed] [Google Scholar]
- 101.Lebovic DI, Chao VA, Martini JF, Taylor RN. IL-1beta induction of RANTES (regulated upon activation, normal T cell expressed and secreted) chemokine gene expression in endometriotic stromal cells depends on a nuclear factor-kappaB site in the proximal promoter. J Clin Endocrinol Metab. 2001;86(10):4759–64. 10.1210/jcem.86.10.7890. [DOI] [PubMed] [Google Scholar]
- 102.Sakamoto Y, Harada T, Horie S, Iba Y, Taniguchi F, Yoshida S, et al. Tumor necrosis factor-alpha-induced interleukin-8 (IL-8) expression in endometriotic stromal cells, probably through nuclear factor-kappa B activation: gonadotropin-releasing hormone agonist treatment reduced IL-8 expression. J Clin Endocrinol Metab. 2003;88(2):730–5. 10.1210/jc.2002-020666. [DOI] [PubMed] [Google Scholar]
- 103.Liu Y, Wang J, Zhang X. An update on the multifaceted role of NF-kappaB in endometriosis. Int J Biol Sci. 2022;18(11):4400–13. 10.7150/ijbs.72707. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Ozaki Y, Ohashi T, Kume S. Potentiation of neutrophil function by recombinant DNA-produced interleukin 1a. J Leukoc Biol. 1987;42(6):621–7. 10.1002/jlb.42.6.621. [DOI] [PubMed] [Google Scholar]
- 105.Berkow RL, Wang D, Larrick JW, Dodson RW, Howard TH. Enhancement of neutrophil superoxide production by preincubation with recombinant human tumor necrosis factor. J Immunol. 1987;139(11):3783–91. [PubMed] [Google Scholar]
- 106.Kocyigit A, Gur S, Erel O, Gurel MS. Associations among plasma selenium, zinc, copper, and iron concentrations and immunoregulatory cytokine levels in patients with cutaneous leishmaniasis. Biol Trace Elem Res. 2002;90(1–3):47–55. 10.1385/BTER:90:1-3:47. [DOI] [PubMed] [Google Scholar]
- 107.Uzzo RG, Leavis P, Hatch W, Gabai VL, Dulin N, Zvartau N, et al. Zinc inhibits nuclear factor-kappa B activation and sensitizes prostate cancer cells to cytotoxic agents. Clin Cancer Res. 2002;8(11):3579–83. [PubMed] [Google Scholar]
- 108.Kim CH, Kim JH, Moon SJ, Chung KC, Hsu CY, Seo JT, et al. Pyrithione, a zinc ionophore, inhibits NF-kappaB activation. Biochem Biophys Res Commun. 1999;259(3):505–9. 10.1006/bbrc.1999.0814. [DOI] [PubMed] [Google Scholar]
- 109.Ho E, Quan N, Tsai YH, Lai W, Bray TM. Dietary zinc supplementation inhibits NFkappaB activation and protects against chemically induced diabetes in CD1 mice. Exp Biol Med (Maywood). 2001;226(2):103–11. 10.1177/153537020122600207. [DOI] [PubMed] [Google Scholar]
- 110.Otsu K, Ikeda Y, Fujii J. Accumulation of manganese superoxide dismutase under metal-depleted conditions: proposed role for zinc ions in cellular redox balance. Biochem J. 2004;377(Pt 1):241–8. 10.1042/BJ20030935. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Goud PT, Goud AP, Joshi N, Puscheck E, Diamond MP, Abu-Soud HM. Dynamics of nitric oxide, altered follicular microenvironment, and oocyte quality in women with endometriosis. Fertil Steril. 2014;102(1):151–9.e5. 10.1016/j.fertnstert.2014.03.053. [DOI] [PubMed] [Google Scholar]
- 112.Hsu AL, Townsend PM, Oehninger S, Castora FJ. Endometriosis may be associated with mitochondrial dysfunction in cumulus cells from subjects undergoing in vitro fertilization-intracyto-plasmic sperm injection, as reflected by decreased adenosine triphosphate production. Fertil Steril. 2015;103(2):347–52.e1. 10.1016/j.fertnstert.2014.11.002. [DOI] [PubMed] [Google Scholar]
- 113.Mate G, Bernstein LR, Torok AL. Endometriosis is a cause of infertility. Does reactive oxygen damage to gametes and embryos play a key role in the pathogenesis of infertility caused by endometriosis? Front Endocrinol (Lausanne). 2018;9:725. 10.3389/fendo.2018.00725. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Xu B, Guo N, Zhang XM, Shi W, Tong XH, Iqbal F, et al. Oocyte quality is decreased in women with minimal or mild endometriosis. Sci Rep. 2015;5:10779. 10.1038/srep10779. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Jiang H, He X, Wang S, Jia J, Wan Y, Wang Y, et al. A micro-tubule-associated zinc finger protein, BuGZ, regulates mitotic chromosome alignment by ensuring Bub3 stability and kinetochore targeting. Dev Cell. 2014;28(3):268–81. 10.1016/j.devcel.2013.12.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Soubry A, Staes K, Parthoens E, Noppen S, Stove C, Bogaert P, et al. The transcriptional repressor Kaiso localizes at the mitotic spindle and is a constituent of the pericentriolar material. PLoS One. 2010;5(2):e9203. 10.1371/journal.pone.0009203. [DOI] [PMC free article] [PubMed] [Google Scholar]