
Fig. 3 |. Modular structure of metazoan Mediator.
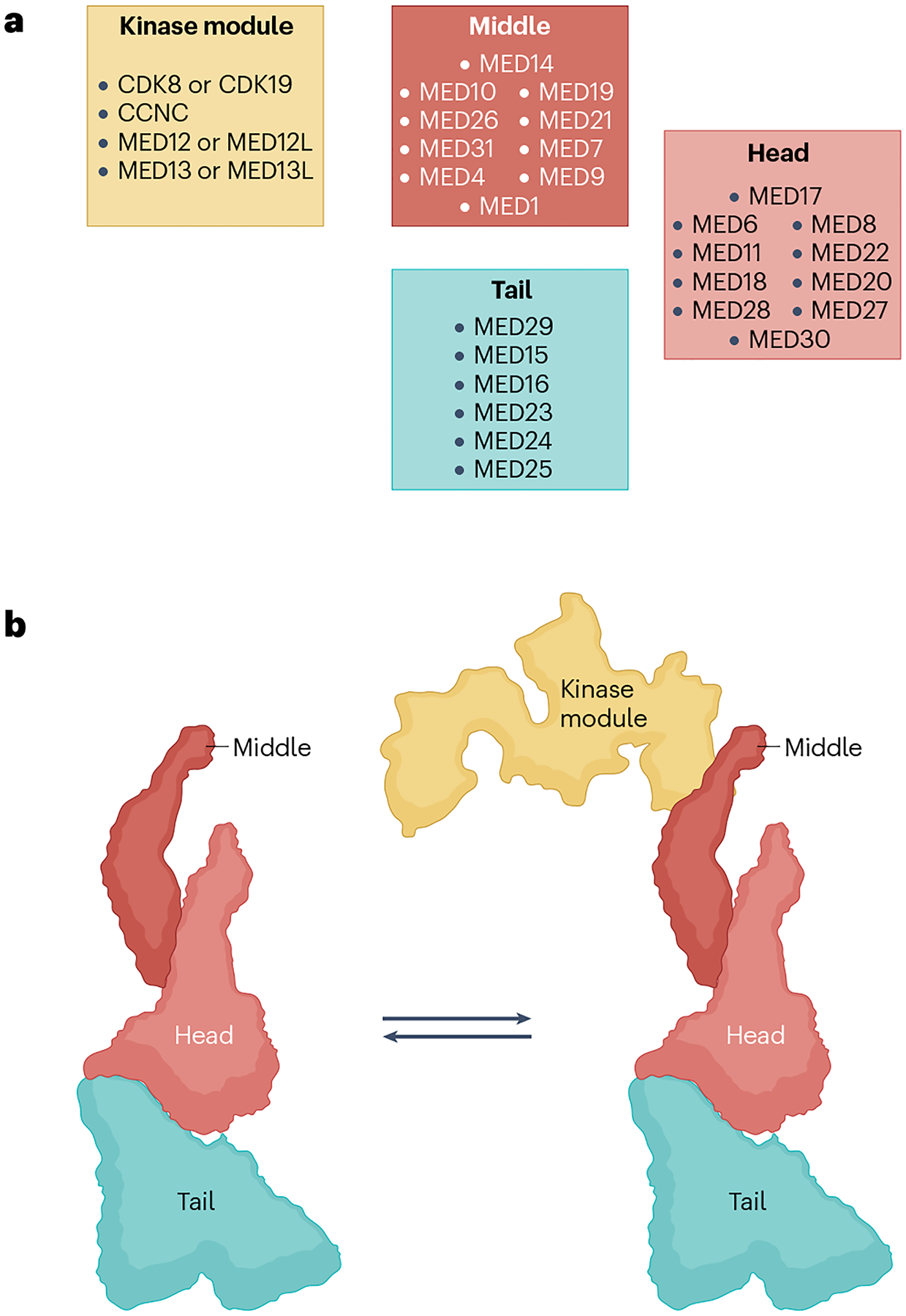
a, Human Mediator structural modules with conventional subunit assignment to the head, middle and tail modules (right). However, cryogenic electron microscopy (cryo-EM) and cross-linking–mass spectrometry studies35–46 reveal that several key subunits straddle more than one module. In particular, MED14 forms a structural backbone around which the head, middle and tail modules are organized. Similarly, whereas the bulk of MED17 resides in the head, its amino terminus is embedded in the middle. Metazoan-specific subunits MED27, MED28 and MED30 also straddle the head and tail. Also note the location of MED1 in the middle. This large subunit has not yet been visualized in its entirety, but the tail-proximal location of this activator target hot spot in Mediator structure is intriguing for models of how activation signals may be processed. The kinase module, which reversibly associates with the Mediator, is also depicted (left). As shown, three of its four constituent subunits have paralogues that can give rise to multiple permutations. b, Left: a model of the Mediator complex showing the relative location of the head, middle and tail modules. The modules can move relative to each other, resulting in several conformers of the complex (not shown). The model is based on ref. 43, which produced high-resolution structures showing detailed subunit architecture of the complex. Right: the Mediator complex in association with the kinase module. Note that this form of the Mediator can interact with activators but not with RNA polymerase II (Pol II). In metazoans, association of the kinase module with the complex is also mutually exclusive with association of the metazoan-specific MED26 middle subunit. Precisely how the two forms of the Mediator interchange is not yet known (see also Fig. 6). The kinase module is modelled after the structure in ref. 147. CDK8, cyclin-dependent kinase 8.