
Fig. 3 |. Fork dynamics regulation by the ATR pathway.
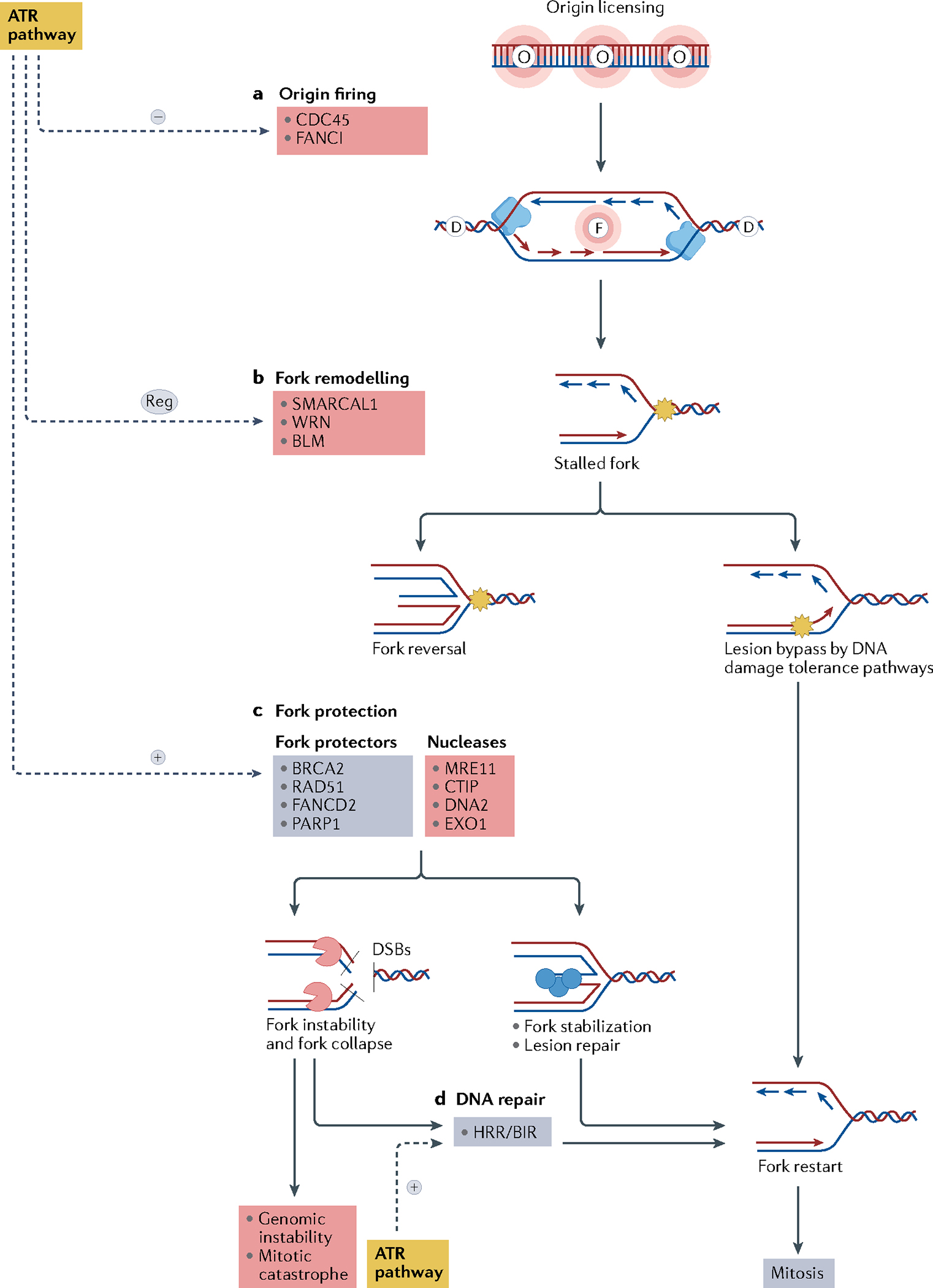
a | The ataxia telangiectasia mutated (ATM) and Rad3-related (ATR) pathway inhibits replication fork origin firing through inhibition of the minichromosome maintenance 2–7 complex (MCM2–7) and Fanconi anaemia group I protein (FANCI). Replication origin firing is indicated by the letter F, and dormant replication origins — licensed but not activated — are indicated by the letter D. Replisome (blue), is a protein complex with helicase and polymerase activities that progresses bidirectionally, starting from the origin firing and forming the replication forks. b | Stalled forks owing to an obstacle to fork progression (shown by the yellow star) may be rescued directly by DNA damage tolerance pathways or may be reversed by fork remodelling proteins. Fork reversal may result in fork protection or fork degradation depending on the balance of activity of various remodelling factors and on the DNA repair proficiency of the cell. c | The ATR pathway modulates fork reversal and activates fork protectors, promoting fork stabilization. Reversed forks are substrates for nucleases that cleave the DNA to form double-strand breaks (DSBs). Fork protectors such as BRCA2, RAD51 and poly[ADP-ribose]polymerase 1 (PARP1) prevent fork degradation by nucleases, allowing lesion repair and fork restart. d | Collapsed replication forks may be repaired by the homologous recombination repair (HRR) and the break-induced replication (BIR) pathways, allowing fork restart and normal mitosis. If DNA repair is not effective, fork collapse may lead to genomic instability and mitotic catastrophe. Mechanisms that lead to fork stabilization and restart and normal mitosis are shown in blue, mechanisms that lead to fork instability, fork collapse and genomic instability are shown in red. BLM, Bloom syndrome protein; CDC45, cell division cycle 45; WRN, Werner syndrome ATP-dependent helicase.