
Table 1.
Dietary anti-aging polyphenols and potential mechanisms.
| Polyphenols | Source | Age-Related Conditions | Model | Dosage | Anti-Aging Activity and Proposed Anti-Aging Mechanism |
|---|---|---|---|---|---|
Resveratrol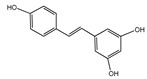
|
Grapes, red wine, peanuts, and blueberries | Age-related motor deficits | Older adults | 500 mg/day, 1000 mg/day | A pilot randomized controlled trial indicates that the combined application of exercise training and resveratrol in elderly people with functional limitations could improve skeletal muscle mitochondrial function and exercise-related physical function indicators [58]. |
| Sarcopenic obesity | Sprague-Dawley (SD) rats | 0.4% of diet | Resveratrol could ameliorate mitochondrial dysfunction and oxidative stress, thereby improving protein metabolism and helping to prevent sarcopenic obesity in the elderly [59]. | ||
| Aging | ddY mice (derived from a non-inbred strain carrying a retrovirus causing significant mortality with age) | 0.4 g/kg of diet | Resveratrol could reduce the level of acetylated proteins in muscles and restore autophagic activity, thereby alleviating age-related sarcopenia and cardiomyocyte hypertrophy [60]. | ||
| Aging | HtrA2 KO mice (the absence of serine/threonine protease HtrA2 causes a PD phenotype) | 25 mg/kg body weight (BW) | Resveratrol treatment can extend the lifespan of HtrA2 KO mice and delay the deterioration of the motor phenotype by attenuating apoptosis at the level of Bax [61]. | ||
| Aging | Caenorhabditis elegans (C. elegans) | 100 mM | Resveratrol, as a Sirtuin activator, can simulate calorie restriction and extend the lifespan of C. elegans [62]. | ||
| Aging | Drosophila melanogaster | 200 μM | Supplementation of resveratrol to larval can effectively eliminate ROS, thereby extending the adult lifespan of fruit flies and not reducing their reproductive capacity [63]. | ||
| Ovarian aging | N. guentheri | 200 μg/g food | Resveratrol could delay ovarian aging by alleviating inflammation and ER stress through the SIRT1/NRF2 pathway [64]. | ||
| Aging | S. cerevisiae | 10, 100, 500 μM | Resveratrol could mimic calorie restriction by stimulating Sir2, increase DNA stability, and extend lifespan by 70% in yeast [65]. | ||
| Aging | Human mononuclear cells (PBMCs) | 5 µM | Compared with the elderly, resveratrol exerts better antioxidant and anti-inflammatory effects in PBMCs of middle-aged individuals [66]. | ||
| Brain aging | Hippocampal astrocytes | 10 μM | Resveratrol could increase the antioxidant defense capacity and reduce pro-inflammatory cytokines in hippocampal astrocyte cultures of rats of all ages, improving NDs [67]. | ||
| Age-related motor deficits | SH-SY5Y cells | 1 or 5 μM | Resveratrol alleviates age-related motor deficits by promoting the survival of dopaminergic neurons and the activation of the ERK1/2 pathway [68]. | ||
Ellagic acid 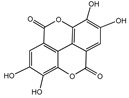
|
Berries, nuts, tea, and medicinal plants [69] | Aging | D-gal-induced aging rats | 120 mg/kg BW | EA improved oxidative damage and inflammation in D-gal-induced aging rats [70]. |
| Hepatic lipid metabolism disorders induced by aging | 25-month-old rats | 30 mg/kg BW | EA affected lipid metabolism in the aged liver via the sirt1/AMPK/sreBP-1c/PPAr-α pathway for age-related metabolic disorders [71]. | ||
| Brain aging | SD rats | 10, 30, 90 mg/kg BW | EA could increase the proliferation of brain neural stem cells and protect brain cell activity through the Wnt/β-catenin signaling pathway to help treat nerve dysfunction, NDs, and aging [72]. | ||
| Osteoporosis | C57BL/6 mice, bone mesenchymal stem cells | Mice: 10, 50 mg/kg BW; cell: 0 μM–30 μM | EA could promote bone formation by activating the SMAD2/3 signaling pathway to ameliorate aging-induced osteoporosis [73]. | ||
| Skin aging | Male SKH-1 hairless mice, human dermal fibroblasts | Mice: 10 μmol/L, cell: 1–10 μmol/L | EA could reduce wrinkles and UV-induced skin inflammation to improve photoaging by decreasing UV-B-induced collagen degradation and inflammatory reactions [74]. | ||
| Aging | C. elegans | 50 μM | EA reduced the injury caused by UV radiation and enhanced stress resistance to extend the lifespan of C. elegans [75]. | ||
| Aging | Drosophila melanogaster | 100 μM and 200 μM | EA can up-regulate the expression of dFOXO, CAT, and SOD, thereby extending lifespan [76]. | ||
| Aging | SH-SY5Y cells | 0.1–1 μM | Lower concentrations of EA could provide better anti-aging benefits than higher concentrations of EA and metformin, presumably via the PPAR-γ/ HO-1 signaling pathway [77]. | ||
| Skin sagging and wrinkling | Human dermal fibroblasts | 2 μg/mL | EA improved skin extracellular matrix production of elastin and collagen and improved skin fine wrinkles [78]. | ||
Urolithin A 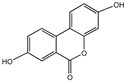
|
Gut microbiota metabolite of EA | Aging | Healthy elderly male and female | 250, 500, 1000 and 2000 mg | UA could promote mitochondrial autophagy and improve muscle health in old animals and in preclinical models of aging [79]. |
| Aging | D-gal-induced aging mice | 3, 15 mg/kg BW | Uro-A from the colon can prevent D-gal-induced aging in mice by blocking NF-κB and mTOR targets, improving motor and cognitive abilities [80]. | ||
| Alzheimer’s disease (AD) | 3xTg-AD mice, B6129SF2/J mice | 5 mg/kg BW | UA could improve the cognition of 3xTg-AD mice and prolong longevity in normal aging mice by inducing autophagy and increasing amyloid-β (Aβ) clearance in neuronal cells [81]. | ||
| Aging | C. elegans, C57BL/6 mice | C. elegans: 50 μM, mice: 50 mg/kg BW | Urolithin A could induce mitophagy and improve mitochondrial and muscle function in C. elegans and rodents, leading to life extension [82]. | ||
| Brain aging | H2O2-induced PC12 cell, D-gal-induced aging mice | Cell: 10, 30, 50 μg/mL, Mice: 50, 100, 150 mg/kg body weight | Urolithin A can exert neuroprotective effects and delay brain aging by activating miR-34a-mediated SIRT1/mTOR signaling pathway [83]. | ||
| Aging | D-gal-induced aging mice | 50, 100, 150 mg/kg BW | UA could attenuate D-gal-induced liver injury in aged mice via antioxidant, anti-inflammatory, and anti-apoptotic properties [84]. | ||
Gallic acid 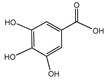
|
Rheum palmatum, Eucalyptus robusta, Cornus officinalis | Age-associated thymic involution | D-gal-treated mice | 200, 250, 500 mg/kg body weight | GA administration may ameliorate age-related thymic degeneration and enhance immune function in the elderly by stimulating FoxN1 expression, increasing proliferating cells, and decreasing apoptotic cells [85]. |
| Aging | H2O2-induced rat’s embryonic fibroblast cells | 1000 μM | GA could postpone aging through its antioxidative stress potential and modulation of mitochondrial complexes’ activities [86]. | ||
| AD | APP/PS1 transgenic AD mouse model | 20 mg/kg body weight | EA could inhibit neuroinflammation and stabilize brain oxidative stress [87]. | ||
Rutin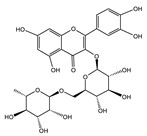
|
Fagopyrum esculentum, Ruta graveolens, and Sophora japonica [88] | Aging | Drosophila melanogaster | 200 μM and 400 μM | Rutin could improve the resistance of male and female Drosophila melanogaster fed with a high-fat diet, increase the expression of age-related genes, and extend longevity [89]. |
| Huntington’s disease | C. elegans | 15, 30, 60, 120 μM | Rutin could inhibit polyglutamine protein aggregation in muscle, decrease neuronal death, and extend longevity through antioxidant, autophagy, and insulin/IGF1 signaling pathways [90]. | ||
| Aging | D-gal-treated mice | 50 mg/kg body weight | Rutin could enhance the biochemical indicators of aging rats by exerting antioxidant effects and regulating apoptosis-related proteins to inhibit cell apoptosis [91]. | ||
| Age-related metabolic dysfunction | Twenty-month-old rats | 25, 50 mg/kg body weight | Rutin suppresses aging-associated mitochondrial dysfunction, endoplasmic reticulum stress, and oxidative stress, thus improving the response to age-related metabolic dysfunction [92]. | ||
| AD | TgAPP mice (a model for AD) | 30 mg/kg body weight | Rutin could raise GSH/GSSG levels, reduce MDA levels, and inhibit APP expression and BACE1 activity, resulting in anti-AD and anti-aging benefits [93]. | ||
Quercetin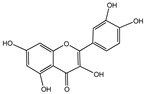
|
Grapes, peaches, onions, garlic [94] | Age-related metabolic dysfunction | Twenty-one-month-old mice | Dasatinib (5 mg/kg BW) and quercetin (50 mg/kg BW) | Dasatinib + quercetin (D + Q) lowers inflammation in adipose tissue and enhances systemic metabolic function in older people [95]. |
| Age-dependent progression of disc degeneration | C57BL/6 mice | Dasatinib (5 mg/kg BW) and quercetin (50 mg/kg BW) | D + Q can target senescent cells non-invasively and lessen the impact of age-dependent degeneration [96]. | ||
| Aging | Macaca fascicularis | Dasatinib (5 mg/kg BW) and quercetin (50 mg/kg BW) | D + Q could prevent aging by strengthening the gut barrier, boosting immunity, and combating inflammation [97]. | ||
| Aging | C57BL/6 mice | Dasatinib (5 mg/kg BW) and quercetin (50 mg/kg BW) | D + Q could lead to selective elimination of senescent cells and the release of senescence-associated pro-inflammatory cytokines [98]. | ||
| Intestinal senescence | BALB/c mice | Dasatinib (5 mg/kg BW) and quercetin (50 mg/kg BW) | Long-term D + Q treatment could decrease the gene expression of senescence and inflammation to alleviate intestinal senescence [9]. | ||
Naringenin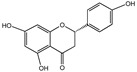
|
Ribes meyeri | Aging | C57BL/6 mice | 25, 50, 100 mg/kg BW | Naringenin could reduce neuronal damage and mitigate systemic inflammation induced by LPS by modulating the expression of NF-κB/TNFα/COX-2/iNOS/TLR4/GFAP [99]. |
| Retinal degeneration | C57BL/6 mice | 100 mg/kg BW | Oral administration of naringenin could regulate mitochondrial dynamics and autophagy to counteract aging-related retinal degeneration [100]. | ||
| Aging | C. elegans, 6-month-old C57BL/6J mice | C. elegans: 100 μM, mice: 100 mg/kg BW | Naringenin could extend the lifespan of C. elegans and slow brain aging in mice by increasing the expression of SIRT enzymes, promoting the activity of metabolic enzymes, and upregulating the expression of anti-aging markers [101]. | ||
Cyanidin-3-O-glucoside (Cy3G)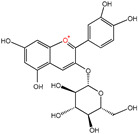
|
Vegetables and berries [102] | AD | HMC3 cell, APPswe/PS1ΔE9 mice (a model for AD) | cell: 50 μM, mice: 30 mg/kg BW | Cy3G could eliminate accumulated β-amyloid and modulate microglial polarization by activating PPARγ and promoting Aβ42 phagocytosis through TREM2 overexpression [103]. |
| AD | APPswe/PS1ΔE9 mice | 30 mg/kg BW | Cy3G could reduce lysosome-associated protein expression, increase autophagy, and modulate the PI3K/Akt/GSK3β signaling pathway to protect neurons and enhance cognitive performance in AD mice [104]. | ||
| AD | SD rats received Aβ in the hippocampus | 10 mg/kg BW | Cy3G could attenuate Aβ-induced tau protein hyperphosphorylation and GSK-3β hyperactivation, possibly rescuing Aβ-induced cognitive deficits through GSK-3β/tau variation [105]. | ||
| AD | APPswe/PS1ΔE9 mice | 30 mg/kg BW | Cy3G may exert therapeutic effects on AD through antioxidant and immunomodulatory mechanisms [106]. | ||
| Parkinson’s disease (PD) | MPTP-induced C57BL/6J | 10, 20, 40 mg/kg BW | Cy3G could play a role in the treatment of PD by modulating the structure and metabolism of gut microbiota [107]. | ||
| Aging | C. elegans | 12.5, 25, 50 μg/mL | Cy3G could enhance resistance and extend the lifespan of polystyrene-exposed C. elegans through the DAF-16 pathway [108]. | ||
| Aging | H9c2 cells | 1 mM | Cy3G could decrease CD38 expression, increase Sirt6 expression in tissues, and restore NAD+ and NK cell levels to exert anti-aging effects through CD38-Sirt6 signaling [109]. | ||
Procyanidin C1 (PCC1)
|
Grape seed extract | Aging | PSC27 cells, WI38 cells, HUVEC cells; C57BL/6J mice | Cell: 1, 5, 10 μg/mL Mice: 20 mg/kg BW |
Low concentrations of PCC1 can inhibit the formation of SASP, while at higher concentrations, it may selectively kill senescent cells by promoting the production of ROS and mitochondrial dysfunction [110]. |
| Aged retina | Aged mice | 8 mg/kg of diet | Long-term PCC1 treatment could relieve function and structural impairment in the aged retina and reduce the accumulation of senescent cells and secretion of SASP [111]. | ||
Nordihydroguaiaretic acid
|
Larrea tridentata | Aging | UM-HET3 mice (designed to model the aging process and age-related diseases) | 2.5, 5 g/kg of diet | Nordihydroguaiaretic acid could increase the survival rate of male mice but does not change the survival rate of female mice, which might be explained by gender differences in steady-state levels or drug metabolism [112]. |
Pterostilbene
|
Blueberries | AD | Senescence accelerated mouse prone 8 (SAMP8) mice | 120 mg/kg of diet | Pterostilbene could increase the expression of peroxisome proliferator-activated receptor (PPAR) α, improve cognitive function and cellular stress regulation ability, and alleviate AD symptoms [113]. |
| AD | 18-month-old rats | 22.5 mg/kg BW | Pterostilbene could increase the expression of postsynaptic density protein 95 and improve the cognitive ability of elderly rats with mild cognitive impairment AD [114]. | ||
| AD | 19-month-old male Fischer | 40, 160 mg/kg of diet | Pterostilbene can effectively reverse cognitive behavioral deficits and dopamine release and improve age-related cognitive degeneration [115]. |