

Abstract
Avian photopic curves show energy peaks at different wavelengths than humans, particularly in the ultraviolet, yellow, and red range. Therefore, an illumination system with a spectrum tailored for laying hens, encompassing the entire visible spectrum, can enhance performance and welfare. The primary contribution of this study was the development of two Spectral Power Distributions (SPDs) specifically designed for laying hens, with different spectral proportions (S1 and S2), and the evaluation of their effects on productive performance, egg quality, and hormonal levels, compared to conventional white lighting at 3000 K (C). The SPD with a higher emission of red light to increased egg production and egg mass. It also had a lower melatonin concentration, suggesting an inverse relationship with the egg production rate. Regarding egg quality, SPDs specifically designed for laying hens resulted in eggs with greater weight (S1), shell strength (S2), and yolk height (S1 and S2) and diameter (S2). The study’s results indicate that lighting emitting wavelengths within the spectrum visible to poultry, with higher emissions at long wavelengths, appears to be more favorable for laying hens than conventional lighting. A spectrum with higher emissions at shorter wavelengths appears to impair the productive performance of laying hens.
Keywords: Lighting, Melatonin, LED, Light spectrum
Subject terms: Engineering, Animal physiology
Introduction
In poultry farming, the strategic use of lighting is crucial for raising laying hens. Beyond providing necessary ambient light, it significantly affects the hens’ physiological processes1. Light influences bird physiology through ocular and transcranial mechanisms. Light stimulation via the eyes activates the optic nerve, which in turn stimulates specific hypothalamic nuclei. Alternatively, light can penetrate the skull bones (transcranially) and activate hypothalamic photoreceptors that transform light energy into neural signals. These signals prompt hypothalamic neurons to synthesize and release gonadotropin-releasing hormone (GnRH) into the bloodstream. GnRH travels to the anterior pituitary gland through a vascular system, the portal venous system, and prompts the release of gonadotropins (luteinizing hormone – LH and follicle-stimulating hormone – FSH)2–4. These hormones facilitate gonadal growth and the production of steroid hormones, such as progesterone and estradiol, from ovarian follicles. Conversely, gonadotropin inhibitory hormone (GnIH) acts as a physiological brake, inhibiting the reproductive axis by reducing hypothalamic GnRH activity and suppressing gonadotropin release in the pituitary5. Research also indicates that GnIH expression may be regulated by the secretion of melatonin6–8. Studies have also demonstrated the relationship between lighting characteristics (photoperiod, intensity, and wavelength) and melatonin synthesis9–11.
The laying hens are significantly affected by the spectrum of light emitted by various sources. Avian photoreceptors in the retinal cones respond to a wider range of the light spectrum (320–700 nm) than human photoreceptors, including sensitivity to ultraviolet rays12. Consequently, hens exhibit peak responses to light stimuli in different energy ranges—primarily in the ultraviolet, yellow, and red spectrums—highlighting their distinct spectral requirements compared to humans. Retinal photoreceptors in hens show heightened sensitivity within the green-yellow wavelengths of the light spectrum, specifically between 545 and 575 nm13.
The presence of an extra cone in the retinal structure of poultry’ eyes enables the transmission of radiation at wavelengths shorter than 400 nm, allowing poultry to perceive part of the UVA spectrum14. This ability allows poultry to locate food that reflects UVA and can impact mate recognition and social interactions, as well as influence mating behavior12,15,16. Therefore, the inclusion of UVA wavelengths in light sources is essential for poultry to express their natural behaviors. Additionally, UV wavelengths induce the synthesis of cholecalciferol (vitamin D3) from 7-dehydrocholesterol in the skin, which can be beneficial for eggshell quality14,15. However, excessive exposure to ultraviolet radiation can damage collagen fibers, destroy vitamin A in the skin, and cause sunburn15. Thus, careful balancing of emitted UVA levels is necessary to avoid harm and ensure poultry’ well-being.
Regarding the productive performance of laying hens, research indicates that long-wavelengths light modulates egg production and the onset of sexual maturity in hens during the laying period3,17–22. Long-wavelength radiation, ranging from 620 to 750 nm, plays a crucial neurovegetative role due to its increased capacity for transcranial penetration13,17. However, the inability of short-wavelength light to stimulate a photosexual response may be due to suboptimal intensity levels. This suggests that light spectra providing higher energy levels at these wavelengths may be more photostimulatory20. Conversely, short-wavelength lighting is associated with improved egg quality23–25 and has been observed to affect growth rates and weight gain in laying hens22,26. Consequently, these specific light colors are increasingly utilized during the rearing and growing phases due to their beneficial effects.
Most research on lighting in poultry production focuses solely on the effects of monochromatic lights or light sources that emit within the human-visible spectrum. Poultry, however, have spectral requirements that differ from those of humans, rendering conventional light sources unsuitable for poultry production. Therefore, there is a need for lighting technologies tailored to the specific spectral requirements of laying hens. Unlike monochromatic lights, broad-spectrum lighting combines various wavelengths of electromagnetic radiation emitted from a light source27. Consequently, the effects of broad spectral power distributions — encompassing the wavelengths of the avian photopic sensitivity curve — on laying hens remain unknown. Furthermore, the effects of different emission proportions within the avian visible spectrum have yet to be investigated. Thus, the appropriate design of light spectral composition according to the requirements of each species and strain, along with practical outcome validation, is crucial1,28.
The literature supports the hypothesis that a lighting system with a spectral power distribution specifically designed to meet the visual needs of laying hens — covering all wavelengths of the avian visible spectrum — benefits the hens and enhances both productivity and egg quality.
This research was designed to integrate the beneficial impacts of diverse electromagnetic radiation wavelengths. Its purpose was to evaluate the effects of two Spectral Power Distributions (SPDs), specifically designed for laying hens and incorporating different spectral proportions, on productive performance, egg quality parameters, and hormonal levels such as progesterone, estradiol, melatonin, and corticosterone of the hens. Furthermore, it aimed to clarify the relationship between hormonal variations and productive outcomes in laying hens.
Results
Productive performance
Laying hens exposed to the S2 spectrum demonstrated superior productive performance compared to other treatments. This group exhibited a higher egg production rate (p = 0.0002), and increased egg mass (p = 0.0003, Table 1). The S1 spectrum did not differ significantly from the control (C) in these parameters. The number of eggs per housed hen was also higher in the S2 group, with respective values of 132.1, 112.1, and 121.1 eggs per hen for the S2, S1, and the control (C) groups. Laying hens under the C spectrum had higher feed consumption.
Table 1.
Average productive performance of laying hens aged 20 to 40 weeks, subjected to spectra S1, S2, and C.
| Variables | Treatments | p-value | ||
|---|---|---|---|---|
| S1 | S2 | C | ||
| Hen-day Egg Production (%) | 79.7 ± 4.59b | 93.5 ± 2.7a | 85.5 ± 3.7b | 0.0002 |
| Egg Mass (g.ave− 1.dia− 1) | 54.1 ± 1.5b | 60.9 ± 0.6a | 56.4 ± 1.3b | 0.0003 |
| Feed Consumption (g.ave− 1.dia− 1) | 135.5 ± 1.9b | 145.5 ± 1.6ab | 149.0 ± 3.0a | 0.0402 |
| Feed Conversion | 2.56 ± 0.10 | 2.45 ± 0.05 | 2.83 ± 0.12 | 0.0500 |
Means ± SE followed by different letters differ by the Tukey’s test at 5% probability.
S1: spectrum developed with color temperature of 4082 K.
S2: spectrum developed with color temperature of 2193 K.
C: spectrum with color temperature 3000 K.
Additionally, weight uniformity among hens remained above 80% throughout the experimental period, with specific values of 93% (average body weight of 1.698 kg), 83% (average body weight of 1.788 kg), and 90% (average body weight of 1.956 kg) recorded at 21, 30, and 40 weeks of age, respectively.
Hormonal analyses
Melatonin concentration was lower in the S2 treatment (p = 0.0135, Table 2). There were no significant differences in the sex hormones progesterone and estradiol among the treatments (p = 0.3040 and p = 0.7603, respectively). Progesterone levels peaked at 26 and 30 weeks of age (10259.5 ± 523.1 pg.mL− 1 and 10914.0 ± 876.2 pg.mL− 1, respectively), coinciding with the peak laying period, and significantly decreased by 34 weeks (6194.9 ± 461.6 pg.mL− 1, p = 0.0000, Fig. 1). The interaction between treatments and age was significant for estradiol (p = 0.0197), though it did not show significance in the post-hoc test. In the S1 and S2 treatments, estradiol levels increased linearly from 26 to 34 weeks, whereas in the C treatment, estradiol levels declined over this period. No significant differences were found among the treatments regarding the stress-related hormone corticosterone (p > 0.05).
Table 2.
Average concentrations of the hormones progesterone, estradiol, melatonin, and corticosterone in laying hens at 26, 30, and 34 weeks of age, subjected to spectra S1, S2, and C.
| Variables | Treatments | p-value | ||||
|---|---|---|---|---|---|---|
| S1 | S2 | C | T | A | TxA | |
| Progesterone (pg/mL) | 9889.1 ± 999.5 | 8654.2 ± 791.2 | 8756.7 ± 717.3 | 0.3040 | 0.0000 | 0.2010 |
| Estradiol (pg/mL) | 11895.3 ± 1108.8 | 11836.6 ± 1183.4 | 10950.3 ± 1103.4 | 0.7603 | 0.1768 | 0.0197 |
| Melatonin (pg/mL) | 150.0 ± 25.6a | 48.2 ± 13.8b | 162.9 ± 31.4a | 0.0135 | 0.2480 | 0.8994 |
| Corticosterone (pg/mL) | 4888.8 ± 778.5 | 4625.2 ± 1660.9 | 4288.7 ± 713.8 | 0.40931 | 0.17341 | 0.62411 |
Means ± SE followed by different letters differ from each other according to the Tukey’s test at 5% probability.
1p-value from the non-parametric Scheirer–Ray–Hare test at 5% probability.
S1: spectrum developed with color temperature of 4082 K.
S2: spectrum developed with color temperature of 2193 K.
C: spectrum with color temperature 3000 K.
T: Treatment. A: Age. TxA: interaction between the Treatment and Age factors.
Fig. 1.

Average concentration of the hormones progesterone, estradiol, melatonin, and corticosterone as a function of the hens’ age. Values are presented as means ± standard error. Different letters indicate significant differences between the ages, as determined by the Tukey’s test at 5% probability.
Egg quality
Laying hens in S1 treatment produced eggs with higher weight (p = 0.0009) compared to the control group (C) (Table 3). In S2 treatment, the hens produced eggs with greater shell strength (p = 0.0120) and larger yolk diameter (p = 0.0188), differing from the control group. Yolk height was higher in eggs from both S1 and S2 compared to those observed in the control group (p = 0.0000). The yolk index, an internal egg quality indicator calculated as the ratio of yolk height to yolk diameter, was higher in the S1 treatment (p = 0.0152). The other parameters did not differ significantly between treatments (p > 0.05).
Table 3.
Average egg quality parameters for laying hens aged 24 to 40 weeks, subjected to spectra S1, S2, and C.
| Variables | Treatments | p-value | ||
|---|---|---|---|---|
| S1 | S2 | C | ||
| Egg Weight (g) | 63.1 ± 0.43a | 62.3 ± 0.29ab | 61.0 ± 0.32b | 0.0009 |
| Albumen Height (mm) | 9.9 ± 0,17 | 9.6 ± 0,15 | 9.6 ± 0,16 | 0.3180 |
| Haugh Unit | 97.3 ± 1,00 | 95.9 ± 0.86 | 96.3 ± 0.94 | 0.5740 |
| Shell Strenght (kgf)2 | 4.3 ± 0.09ab | 4.5 ± 0.07a | 4.2 ± 0.08b | 0.01201 |
| Shell Thickness (mm) | 0.35 ± 0.003 | 0.34 ± 0.002 | 0.34 ± 0.003 | 0.1555 |
| Yolk Height (mm) | 19.0 ± 0.06a | 18.8 ± 0.07a | 18.4 ± 0.08b | 0.0000 |
| Yolk Diameter (mm) | 46.8 ± 1.50ab | 50.9 ± 1.80a | 45.2 ± 1.20b | 0.0188 |
| Yolk Index | 0.43 ± 0,007a | 0.40 ± 0.008b | 0.42 ± 0.006ab | 0.0152 |
Means ± SE followed by different letters differ from each other according to the Tukey test at 5% probability.
1p-value from the non-parametric Scheirer–Ray–Hare test.
2Means ± SE followed by different letters differ from each other according to the Dunn test at 5% probability.
S1: spectrum developed with color temperature of 4082 K.
S2: spectrum developed with color temperature of 2193 K.
C: spectrum with color temperature 3000 K.
Discussion
Research on the effects of different proportions of electromagnetic radiation within a broad light spectrum is crucial for enhancing the efficiency, production, and welfare of laying hens. Despite considerable research on other aspects of lighting, studies on the effects of full-spectrum light based on the photopic spectrum curve of poultry (encompassing UVA to red wavelengths) are limited.
Laying hens subjected to the S2 spectrum showed better productive performance than those exposed to the other treatments evaluated in the study. This result may be related to the S2 spectrum’s higher light emission in the orange and red wavelengths (21.3% and 43.7%, respectively) and reduced emission in the blue, cyan, and green wavelengths (4.4%, 1.6%, and 14.7%, respectively). In contrast, the S1 spectrum has a higher emission in the green wavelength (28.8%) and lower emission in the orange and red wavelengths (17.8% and 19.7%, respectively). Studies have shown that long-wavelength radiation (orange and red) plays a role in neurovegetative function, influences the sexual maturity of laying hens5,29, affects egg production22,30,31, and improves feed consumption efficiency5. Long-wavelength radiation has a greater capacity to penetrate the bones of the transcranial region and the brain tissue of hens, thereby stimulating the photoreceptors in the hypothalamus. According to Baxter et al.3, for reproduction purposes, the most important pathway for light transduction in poultry is the transcranial pathway.
Furthermore, the results of this study suggest that higher light emission in the blue and green wavelengths may have contributed to the lower productive performance of the laying hens in S1. Short-wavelength light, such as blue and green, can delay sexual maturation1,20,32,33 and appears to inhibit reproductive activity17,34. These findings align with those observed by Wichman et al.28, who found that laying hens exposed to light with more energy in the blue and green spectrum and less in the red tended to start laying later. Raziq et al.31 found a higher egg production rate and better feed conversion in laying hens exposed to red monochromatic lighting compared to dichromatic lighting composed of green and red wavelengths.
Regarding ultraviolet radiation (UVA) in treatments S1 and S2, it is not possible to confirm a beneficial effect on egg production, as it comprises short wavelengths (320 to 400 nm). Furthermore, egg production in treatment S1 did not differ from the control treatment (C). However, Rana et al.35 observed a trend toward increased egg production and accelerated sexual maturity in laying hens subjected to daily UVA and UVA/B light supplementation. Wei et al.36 reported an increase in egg production rate in laying hens during the later phase of the laying cycle when exposed to UVB light supplementation. According to Wichman et al.28, the ideal amount of UV exposure for laying hens remains unclear, but proportions similar to natural light may be beneficial.
Light sources can elicit different responses in laying hens. Research on this topic shows divergent or limited results, particularly for light-emitting diode (LED) sources37. In this study, the higher concentration of energy in the long-wavelength light spectrum (red) may have significantly influenced the melatonin levels observed in laying hens in the S2 treatment compared to the S1 treatment, which has a higher energy concentration in short wavelengths (blue, cyan, and green). Additionally, the spectrum with higher emission in the green wavelength range (S1) was associated with higher melatonin levels and lower egg production rates. Some studies indicate that melatonin, synthesized in the pineal gland and retina, acts directly on the hypothalamus and induces the expression of GnIH (gonadotropin-inhibitory hormone), which inhibits the secretion of GnRH6 and consequently compromises the secretion of LH and FSH7,38. The results of this study are consistent with those reported by Liu et al.10, who found a negative correlation between egg production and plasma melatonin levels. Furthermore, the low melatonin level of 48.2 pg/mL observed in S2 is compatible with the 86 pg/mL levels found by Poudel et al.39, who evaluated the effect of red LED lighting on egg production. Studies conducted on broiler chickens have also shown that the green light spectrum can stimulate melatonin secretion40,41. Thus, melatonin may represent a potential indicator of the influence of light on physiological responses related to the reproductive axis of laying hens.
A linear increasing trend in estradiol secretion was observed in the S2 treatment between 26 and 34 weeks of age, suggesting that lighting systems with higher proportions of red spectrum light can increase estradiol concentrations, a phenomenon reported in other studies5,30. However, a similar effect was observed in the S1 group, which has a higher green light emission in its spectrum. The metabolic processes through which estradiol stimulation occurred in this treatment, whether via the hypothalamus or directly at the ovarian level, remain uncertain. After sexual maturation, increasing levels of estradiol and progesterone act negatively on the mRNA expression of the GnIH receptor in the gonads, reducing the inhibitory effect of GnIH at the ovarian level5,42. However, melatonin concentrations directly regulate GnIH secretion in the hypothalamus of laying hens. Therefore, the higher melatonin levels observed in S1 may have stimulated the secretion of the neuropeptide GnIH at the hypothalamic level, which could be associated with the reduced egg production rate observed in S1 compared to S2. The ovaries of laying hens contain several pre-hierarchical follicles that, after maturation, enter the hierarchy of pre-ovulatory follicles. Pre-hierarchical follicles predominantly secrete estrogen and androgen42. We hypothesize that the presence of GnIH in the bloodstream may have activated the expression of the GnIH receptor in the laying hens’ ovaries, inhibiting pre-hierarchical follicles from maturing and entering the hierarchy. This could explain the similar levels of estradiol and the differences in egg production rates. Furthermore, in addition to reproductive hormones, other physiological factors, including energy metabolism and the action of intraovarian and epidermal growth factors, tightly regulate egg production in laying hens43.
Progesterone levels peaked at 26 and 30 weeks of age, coinciding with the peak laying period, and then significantly declined by 34 weeks, regardless of the spectrum applied. No association was observed between progesterone levels and egg production in this study. This result agrees with findings by Onagbesan et al.43, who also observed maximum progesterone levels at peak laying age and found no relationship between this hormone and differences in egg production rates among the treatments evaluated, suggesting that progesterone may be a predictor of peak egg production. Other researchers likewise found no relationship between progesterone levels and differences in egg production rates44,45.
Corticosterone, a hormone produced by the adrenal glands and the main indicator of the stress axis in poultry, can vary daily between 7 and 11 ng/mL in laying hens3,46. The average levels obtained in this study were below these limits, indicating that the laying hens were not under physiological stress during the experimental period. No difference in corticosterone levels was observed between treatments. Archer et al.47 found lower corticosterone concentrations in laying hens exposed to light with a higher concentration of energy in the red spectrum. Research has shown that poultry exposed to UV radiation have lower plasma corticosterone levels. Sobotik et al.48 found that raising laying hens under UV light as part of the lighting spectrum reduced their stress and fear response. The authors emphasized the importance of using appropriate lighting to optimize poultry welfare. Results from House et al.49 suggest that broilers raised under ultraviolet light have lower corticosterone levels, reduced susceptibility to stress, and fewer fear responses.
Regarding egg quality, eggs from the S1 and S2 treatments had greater weight (S1), stronger shell strength (S2), and increased yolk height (S1 and S2) and diameter (S2). This result may be related to estradiol, which stimulates hepatic synthesis of the main yolk components and shell calcification2. It may also be associated with ultraviolet radiation in the S1 and S2 spectra. According to England and Ruhnke14, the benefits of UV light are mediated by its ability to activate cholecalciferol from 7-dehydrocholesterol in the skin, resulting in improved eggshell quality. However, Sobotik et al.48 and Wei et al.36 did not observe an effect of ultraviolet radiation on egg quality. Li et al.25, however, observed increased egg weight in laying hens raised under red light and reported greater shell thickness and strength under green light. Other studies have also found benefits of green light on egg quality24,31.
In conclusion, the study highlights the importance of tailoring the light spectrum to match the visible electromagnetic radiation perceived by poultry, particularly with a higher emission of long wavelengths such as orange and red, which have been shown to enhance productive performance in laying hens. UVA radiation within the light spectrum is beneficial, contributing to improved egg quality measurements. Moreover, plasma melatonin levels have emerged as a potential marker for assessing the impact of light on physiological responses within the avian reproductive axis. Progesterone levels serve as predictors for peak egg production. Conversely, an increase in estradiol levels may not necessarily correlate with higher laying productivity. It is also important to note that higher light emission in the blue and green wavelengths may negatively impact laying hen productivity. This body of evidence shows the relationship between light exposure and avian physiology, suggesting that strategic lighting could be a powerful tool for optimizing laying hens’ performance. This study was conducted in a controlled environment, allowing for an understanding of the evaluated effects without external interferences. However, future studies on a commercial poultry production scale could further assess the generalizability of these findings.
Methods
Ethical approval
This research was approved by the Animal Use Ethics Committee of the State University of Campinas – CEUA/UNICAMP, Process # 5960-1/2022. All methods were carried out in accordance with relevant guidelines, regulations, and the ARRIVE guidelines. No anesthesia or euthanasia procedures were performed on the animals during the study.
Trial
In this study, two Spectral Power Distributions (SPDs) with different spectral proportions optimized for laying hens were developed. These specialized SPDs were compared to conventional lighting at a 3000 K color temperature. The installation and trials were conducted within the controlled environment of the FEAGRI/UNICAMP Thermal Comfort Laboratory, located in Campinas, São Paulo (as shown in Fig. 2).
Fig. 2.

Laboratory with three compartments where the experiment was conducted.
The laboratory setup consisted of three compartments measuring 1.4 × 1.6 m each. These compartments were separated by dual insulated blackout panels, 1.5 cm thick, with a front wall made of dual-sided expanded polystyrene, 7 cm thick, for thermal insulation. Each compartment had a doorway (0.7 × 2.0 m) that allowed access from the main passageway, facilitating the independent operation of each unit. All rooms were equipped with environmental controls to regulate ammonia levels, temperature, and humidity, and were continuously monitored via video surveillance. Each room included a feeder, drinkers, a perch, and a nesting area. The flooring substrate was composed of a 5 cm layer of sterile wood shavings.
Spectral power distributions for laying hens
Two SPDs were designed to cover the full range of avian photopic sensitivity, spanning electromagnetic wavelengths from 380 to 740 nm, each with distinct spectral distributions (Table 4; Fig. 3). Prototype lamps were developed to emit these SPDs using a combination of light-emitting diodes (LEDs) capable of producing white, ultraviolet-A (UVA), blue, cyan, and green light. The flicker especification was developed in accordance with IEEE 1789 (Recommended Practices for Modulating Current in High-Brightness LEDs to Mitigating Health Risks to Viewers), specifying that flicker percentage should be less than 8% at a frequency of 120 Hz.
Table 4.
Characteristics of the spectral power distribution developed for laying hens.
| Description | |
|---|---|
| S1 | The spectral power distribution was configured to include wavelengths ranging from 380 to 740 nm, with its composition featuring 3.2% UVA emission (380–400 nm), 12.6% in the blue band (400–485 nm), 5.1% in the cyan band (485–500 nm), 28.8% in the green band (500–565 nm), 12.8% in the yellow band (565–590 nm), 17.8% in the orange band (590–625 nm), and 19.7% in the red band (625–740 nm). This configuration yields a color temperature of 4082 K |
| S2 | The spectral power distribution spans wavelengths from 380 to 740 nm. Its composition includes 4.5% emission in the UVA band (380–400 nm), 4.4% in the blue band (400–485 nm), 1.6% in the cyan band (485–500 nm), 14.7% in the green band (500–565 nm), 9.9% in the yellow band (565–590 nm), 21.3% in the orange band (590–625 nm), and a dominant 43.7% in the red band (625–740 nm). This combination produces a color temperature of 2193 K |
| C | The control spectral power distribution, with a color temperature of 3000 K, spans wavelengths from 400 nm to 740 nm. Its spectral composition includes 11.5% blue light (400–485 nm), 3.0% cyan (485–500 nm), 23.3% green (500–565 nm), 14.1% yellow (565–590 nm), 23.0% orange (590–625 nm), and 25.0% red (625–740 nm). This spectrum serves as the control treatment |
Fig. 3.

Spectral power distribution of S1 (a), the S2 (b), and C (c).
Laying hens and management
The study was conducted on laying hens aged 20 to 40 weeks. The strain of laying hens used in the experiment was Hisex Brown (Gallus gallus domesticus). Lighting was adjusted to provide an illuminance level between 5 and 10 lx at the hens’ height. Weekly 30-minute increments were added to the light exposure until the photoperiod reached 16 h of light and 8 h of darkness (16 L:8D). The laying hens in each group were identified according to treatment they received and remained marked throughout the experimental period. All hens were kept under the same temperature and relative humidity conditions in each room. The air conditioning system was set to maintain temperatures between 20 and 22 °C. Commercial feed was provided to the hens, with feed and water replenished daily in the morning, both offered ad libitum. The flooring substrate was replaced periodically as needed. The hens were weighed weekly to monitor body weight, and eggs were counted and collected daily in the morning period.
Experimental design
The experimental design followed a 3 × 3 Latin square, featuring three treatments (S1, S2, and C) and three trials (E1 = 20th to 26th week of age; E2 = 27th to 33rd week of age; E3 = 34th and 40th week of age) (Table 4). Ten laying hens were randomly assigned to each treatment (totaling 30 hens in the experiment), with a housing density of 5 hens per m2 of total area. The number of hens in each treatment was determined based on housing density recommendations by the European Commission50, which advises a stocking density not exceeding 9 hens per m² of usable area for cage-free systems and 6 hens per m² for organic production systems, allowing the hens to move freely. Treatments were randomly assigned to different rooms in each trial to ensure that each treatment was applied in all rooms, mitigating potential positional effects. The assignment of treatments to compartments is detailed in Table 5.
Table 5.
Distribution of treatments (S1, S2, and C) in a latin square design within the rooms of the environmental chamber for each trial (E1, E2, and E3).
| Trials | Rooms | ||
|---|---|---|---|
| R1 | R2 | R3 | |
| E1 | S2 | S1 | C |
| E2 | C | S2 | S1 |
| E3 | S1 | C | S2 |
E1 = 20th to 26th week of age; E2 = 27th to 33rd week of age; E3 = 34th and 40th week of age.
Data collection and measurements
Production
The number of eggs produced over 24 h was recorded. Hen-day egg production (%) was calculated weekly based on the hens’ age.
Egg quality
Egg quality was assessed using various metrics, including egg weight (g), albumen height (mm), Haugh unit, shell thickness (mm), shell strength (kgf), yolk height (mm), yolk diameter (mm), and yolk index. All eggs were analyzed within 6 h of collection. These measurements were performed using a digital egg analyzer (model NABEL DET 6000, NABEL Co., Ltd., Kyoto, Japan).
Egg mass
Egg mass was calculated by multiplying egg production by egg weight, with the result expressed in g.hens− 1.day− 1.
Zootechnical performance
The laying hens’ body weight (kg), feed consumption (g), and feed conversion were measured. Hens were weighed at 21, 30, and 40 weeks of age. Average hen weights and weight uniformity were calculated for each treatment. Feed consumption was determined by calculating the difference between the amount of feed supplied and the remaining feed in the feeder. Feed conversion was calculated by dividing feed consumption (g.hens− 1.day− 1) by egg mass (g.hens− 1.day− 1).
Hormonal analyses
Blood samples (2 mL) were drawn via brachial vein puncture, and fecal samples were collected from five randomly selected hens from each treatment (n = 5)10. Sampling occurred when the hens were 26, 30, and 34 weeks of age. Blood was collected between 9 a.m. and 10 a.m. using Heparin-EDTA, after which plasma was immediately separated by centrifugation at 3000 rpm for 15 min and stored at -80 °C for subsequent hormonal analysis. Fecal samples were dried in an oven at 60 °C. After drying, 0.2 g of solid fecal material was subjected to ethanol extraction followed by overnight agitation. The sample was then centrifuged at 5000 rpm for 15 min at 4 °C, and the supernatant was stored at -20 °C for hormonal analysis. Hormonal concentrations of estradiol, progesterone, melatonin (related to the reproductive axis), and corticosterone (a stress indicator) were measured using commercial ELISA kits (DetectX® Arbor Assays and ELK Biotechnology) at the Institute of Biology at UNICAMP. Corticosterone and melatonin concentrations were measured in blood plasma samples, while estradiol and progesterone were measured in fecal samples. All analyses were conducted according to the manufacturers’ guidelines.
Statistical analyses
The data were analyzed using generalized linear models (GLM). Descriptive analyses were conducted on the data, along with skewness and kurtosis analyses51.
The model outlined in Eq. 1 was applied to the productive performance and egg quality variables.
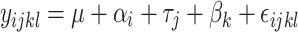 |
1 |
where µ represents the overall mean, 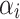 is the effect of the trials,
is the effect of the trials, 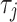 is the effect of the rooms,
is the effect of the rooms, 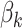 is the effect of the treatments, and
is the effect of the treatments, and 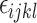 is the random error. This analyses included 63 observations (3 treatments x 3 trials x 7 weeks of age within each trial – E1 = 20th to 26th week of age; E2 = 27th to 33rd week of age; E3 = 34th and 40th week of age) (Table 5). The group of 10 hens represented the experimental unit.
is the random error. This analyses included 63 observations (3 treatments x 3 trials x 7 weeks of age within each trial – E1 = 20th to 26th week of age; E2 = 27th to 33rd week of age; E3 = 34th and 40th week of age) (Table 5). The group of 10 hens represented the experimental unit.
The model described in Eq. 2 was applied to the variables related to hormonal levels.
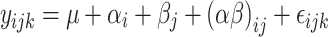 |
2 |
where µ is the overall mean, 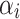 is the effect of the treatments,
is the effect of the treatments, 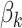 is the effect of the hens’ age,
is the effect of the hens’ age, 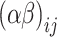 is the interaction effect between the treatments and the ages evaluated (26, 30, and 34 weeks of age), and
is the interaction effect between the treatments and the ages evaluated (26, 30, and 34 weeks of age), and 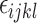 represents the random error. This analysis included 45 observations (3 treatments x 3 ages x 5 hens). Each hen in the sample represented an experimental unit.
represents the random error. This analysis included 45 observations (3 treatments x 3 ages x 5 hens). Each hen in the sample represented an experimental unit.
The data were subjected to analysis of variance (ANOVA), followed by Tukey’s multiple comparison test at a 5% significance level. The assumptions of normality and homogeneity of variances were verified through residual analysis using the Shapiro-Francia test and the Breusch-Pagan test, respectively. Variables that did not meet the normality assumption were transformed using the Box-Cox52 method. Variables that remained non-normal after transformation were analyzed using the nonparametric Scheirer–Ray–Hare test, followed by the Dunn test for mean comparisons. RStudio software was used for these analyses.
Acknowledgements
The author is grateful to the grant #2020/09523-0, São Paulo Research Foundation (FAPESP) for the financial support given to the current study, the Ministry of Science, Technology and to the Innovation and National Council for Scientific and Technological Development (CNPq), process n. 424186/2021-7, for the payment of scholarships. To Audax Electronics LTDA for their collaboration in the development of spectrum curves.
Author contributions
J.S.G.B. designed and participated in the entire experiment and data collection and wrote the manuscript. K.S. participated in the experiment and data collection. J.R.B. participated in the blood collection. T.F.P. and M.J.S. collected and prepared blood and fecal samples from the birds and carried out all hormonal analyses. H.V. and V.A.S. participated in developing spectrum curves and lamp prototypes. R.G.S. participated in preparing data and writing the article. D.J.M. directed all experiments. All authors reviewed the manuscript.
Data availability
All data analyzed in the study will be shared, as well as statistical analysis plans. The data will become available immediately following publication and with no end date. Access criteria data will be shared (including with anyone who wishes to access the data for any purpose), and proposal should be directed to jgbarros@unicamp.br.
Declarations
Competing interests
The authors declare no competing interests.
Footnotes
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Sun, Y. et al. Effects of LED lights with defined spectral proportion on growth and reproduction of indigenous Beijing-You chickens. Animals13, 616. 10.3390/ani13040616 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Etches, R. J. Calcium logistics in the laying hen. J. Nutr.117, 619–628. 10.1093/jn/117.3.619 (1987). [DOI] [PubMed] [Google Scholar]
- 3.Baxter, M., Joseph, N., Osborne, V. R. & Bédécarrats, G. Y. Red light is necessary to activate the reproductive axis in chickens independently of the retina of the eye. Poult. Sci.93, 1289–1297. 10.3382/ps.2013-03799 (2014). [DOI] [PubMed] [Google Scholar]
- 4.Han, S. et al. Influence of three lighting regimes during ten weeks growth phase on laying performance, plasma levels- and tissue specific gene expression- of reproductive hormones in Pengxian yellow pullets. Plos One12, 1–11. 10.1371/journal.pone.0177358 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Baxter, M. & Bédécarrats, G. Y. Evaluation of the impact of light source on reproductive parameters in laying hens housed in individual cages. J. Poult. Sci.56, 148–158. 10.2141/jpsa.0180054 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Ubuka, T., Bentley, G. E., Ukena, K., Wingfield, J. C. & Tsutsui, K. Melatonin induces the expression of gonadotropin-inhibitory hormone in the avian brain. Proc. Natl. Acad. Sci. USA102, 3052–3057. 10.1073/pnas.0403840102 (2005). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Chowdhury, V. S. et al. Melatonin stimulates the release of gonadotropin-inhibitory hormone by the avian hypothalamus. Endocrinology151, 271–280. 10.1210/en.2009-0908 (2010). [DOI] [PubMed] [Google Scholar]
- 8.Chowdhury, V. S., Ubuka, T. & Tsutsui, K. Review: Melatonin stimulates the synthesis and release of gonadotropin-inhibitory hormone in birds. Gen. Comp. Endocrinol.181, 175–178. 10.1016/j.ygcen.2012.08.005 (2013). [DOI] [PubMed] [Google Scholar]
- 9.Jin, E. et al. Effect of monochromatic light on melatonin secretion and arylalkylamine N-acetyltransferase mRNA expression in the retina and pineal gland of broilers. Anat. Rec. Adv. Integr. Anat. Evol. Biol.294, 1233–1241. 10.1002/ar.21408 (2011). [DOI] [PubMed] [Google Scholar]
- 10.Liu, L. et al. Effect of monochromatic light on expression of estrogen receptor (ER) and progesterone receptor (PR) in ovarian follicles of chicken. Plos One10, 1–14. 10.1371/journal.pone.0144102 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Li, S. et al. Melatonin mediates monochromatic Yght-induced insulin-like growth factor 1 secretion of chick liver: Involvement of membrane receptors. Photochem. Photobiol.92, 595–603. 10.1111/php.12594 (2016). [DOI] [PubMed] [Google Scholar]
- 12.Prescott, N. B. & Wathes, C. M. Spectral sensitivity of the domestic fowl (Gallus g. domesticus). Br. Poult. Sci.40, 332–339. 10.1080/00071669987412 (1999). [DOI] [PubMed] [Google Scholar]
- 13.Lewis, P. D. & Morris, T. R. Poultry and coloured light. Worlds. Poult. Sci. J.56, 189–207. 10.1079/WPS20000015 (2000). [Google Scholar]
- 14.England, A. & Ruhnke, I. The influence of light of different wavelengths on laying hen production and egg quality. Worlds. Poult. Sci. J.76, 443–458. 10.1080/00439339.2020.1789023 (2020). [Google Scholar]
- 15.Lewis, P. D. & Gous, R. M. Responses of poultry to ultraviolet radiation. World’s Poultry Science Journal65, 499–510 (2009). [Google Scholar]
- 16.Jones, E. K. M., Prescott, N. B., Cook, P., White, R. P. & Wathes, C. M. Ultraviolet light and mating behaviour in domestic broiler breeders. British Poultry Science42, 23–32 (2001). [DOI] [PubMed] [Google Scholar]
- 17.Mobarkey, N., Avital, N., Heiblum, R. & Rozenboim, I. The role of retinal and extra-retinal photostimulation in reproductive activity in broiler breeder hens. Domest. Anim. Endocrinol.38, 235–243. 10.1016/j.domaniend.2009.11.002 (2010). [DOI] [PubMed] [Google Scholar]
- 18.Gongruttananun, N. Influence of red light on reproductive performance, eggshell ultrastructure, and eye morphology in Thai-native hens. Poult. Sci.90, 2855–2863. 10.3382/ps.2011-01652 (2011). [DOI] [PubMed] [Google Scholar]
- 19.Gongruttananun, N. & Guntapa, P. Effects of red light illumination on productivity, fertility, hatchability and energy efficiency of Thai indigenous hens. Kasetsart J. - Nat. Sci.46, 51–63 (2012). [Google Scholar]
- 20.Min, J. K. et al. Effect of monochromatic light on sexual maturity, production performance and egg quality of laying hens. Avian Biol. Res.5, 69–74. 10.3184/175815512X13350270679453 (2012). [Google Scholar]
- 21.Huber-Eicher, B., Suter, A. & Spring-Stahli, P. Effects of colored light-emitting diode illumination on behavior and performance of laying hens. Poult. Sci.92, 869–873. 10.3382/ps.2012-02679 (2013). [DOI] [PubMed] [Google Scholar]
- 22.Yang, Y. F. et al. The relationship of spectral sensitivity with growth and reproductive response in avian breeders (Gallus gallus). Sci. Rep.6, 19291. 10.1038/srep19291 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Er, D., Wang, Z., Cao, J. & Chen, Y. Effect of monochromatic light on the egg quality of laying hens. J. Appl. Poult. Res.16, 605–612. 10.3382/japr.2006-00096 (2007). [Google Scholar]
- 24.Hassan, M. R., Sultana, S., Choe, H. S. & Ryu, K. S. Effect of combinations of monochromatic LED light color on the performance and behavior of laying hens. J. Poult. Sci.51, 321–326. 10.2141/jpsa.0130105 (2014). [Google Scholar]
- 25.Li, D. et al. The effect of monochromatic light-emitting diode light on reproductive traits of laying hens. J. Appl. Poult. Res.23, 367–375. 10.3382/japr.2013-00746 (2014). [Google Scholar]
- 26.Astuti, P. et al. Corticosterone levels, heterophil/lymphocyte ratios and growth rates in Lohmann Indian river chickens raised under monochromatic blue light. Int. J. Poult. Sci.14, 639–643. 10.3923/ijps.2015.639.643 (2015). [Google Scholar]
- 27.Archer, G. S. Color temperature of light-emitting diode lighting matters for optimum growth and welfare of broiler chickens. Animal12, 1015–1021. 10.1017/S1751731117002361 (2018). [DOI] [PubMed] [Google Scholar]
- 28.Wichman, A., De Groot, R., Håstad, O., Wall, H. & Rubene, D. Influence of different light spectrums on behaviour and welfare in laying hens. Animals11, 1–15. 10.3390/ani11040924 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Wei, Y., Zheng, W., Li, B., Tong, Q. & Shi, H. Effects of a two-phase mixed color lighting program using light-emitting diode lights on layer chickens during brooding and rearing periods. Poult. Sci.99, 4695–4703. 10.1016/j.psj.2020.06.072 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Takeshima, K., Hanlon, C., Sparling, B., Korver, D. R. & Bédécarrats, G. Y. Spectrum lighting during pullet rearing and its impact on subsequent production performance in layers. J. Appl. Poult. Res.28, 1262–1278. 10.3382/japr/pfz094 (2019). [Google Scholar]
- 31.Raziq, F., Hussain, J., Mahmud, A. & Javed, K. Effect of light-emitting diode (LED)-based colors on production performance and welfare of commercial layers. Turkish J. Vet. Anim. Sci.44, 1269–1278. 10.3906/vet-2007-30 (2020). [Google Scholar]
- 32.Hassan, M. R., Sultana, S., Choe, H. S. & Ryu, K. S. Effect of monochromatic and combined light colour on performance, blood parameters, ovarian morphology and reproductive hormones in laying hens. Ital. J. Anim. Sci.12, 359–364. 10.4081/ijas.2013.e56 (2013). [Google Scholar]
- 33.Elkomy, H. E., Taha, A. E., Basha, H. A., Abo-Samaha, M. I. & Sharaf, M. M. Growth and reproduction performance of Japanese quails (Coturnix coturnix japonica) under various environments of light colors. Slov. Vet. Res.56, 119–127. 10.26873/SVR-749-2019 (2019). [Google Scholar]
- 34.Mobarkey, N., Avital, N., Heiblum, R. & Rozenboim, I. The effect of parachlorophenylalanine and active immunization against vasoactive intestinal peptide on reproductive activities of broiler breeder hens photostimulated with green light. Biol. Reprod.88, 1–7. 10.1095/biolreprod.112.103697 (2013). [DOI] [PubMed] [Google Scholar]
- 35.Rana, M. S., Clay, J., Regmi, P. & Campbell, D. L. M. Minimal effects of ultraviolet light supplementation on egg production, egg and bone quality, and health during early lay of laying hens. PeerJ11, e14997. 10.7717/peerj.14997 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Wei, Y. et al. Effects of B-wave ultraviolet supplementation using light-emitting diodes on caged laying hens during the later phase of the laying cycle. Animals10, 15. 10.3390/ani10010015 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Bahuti, M. et al. Evaluation of different light intensities on the well-being, productivity, and eggs quality of laying hens. Comput. Electron. Agric.10.1016/j.compag.2023.108423 (2023). [Google Scholar]
- 38.Martin, J. E., McKellar, S. & Klein, D. C. Melatonin inhibition of the in vivo pituitary response to luteinizing hormone-releasing hormone in the neonatal rat. Neuroendocrinology31, 13–17. 10.1159/000123044 (1980). [DOI] [PubMed] [Google Scholar]
- 39.Poudel, I., Beck, M. M., Kiess, A. S. & Adhikari, P. The effect of blue and red LED light on the growth, egg production, egg quality, behavior, and hormone concentration of Hy-Line W-36 laying hens. J. Appl. Poult. Res.31, 100248. 10.1016/j.japr.2022.100248 (2022). [Google Scholar]
- 40.Zhang, L., Cao, J., Wang, Z., Dong, Y. & Chen, Y. Melatonin modulates monochromatic light-induced GHRH expression in the hypothalamus and GH secretion in chicks. Acta Histochemica118, 286–292 (2016). [DOI] [PubMed] [Google Scholar]
- 41.Yue, L. et al. Melatonin receptor Mel1b- and Mel1c-mediated green light induced the secretion of growth hormone in anterior pituitary of chicks. Photochemistry and Photobiology95, 1387–1394 (2019). [DOI] [PubMed] [Google Scholar]
- 42.Maddineni, S. R., Ocón-Grove, O. M., Krzysik-Walker, S. M., Hendricks, G. L. & Ramachandran, R. Gonadotropin-inhibitory hormone (GnIH) receptor gene is expressed in the chicken ovary: Potential role of GnIH in follicular maturation. Reproduction135, 267–274. 10.1530/REP-07-0369 (2008). [DOI] [PubMed] [Google Scholar]
- 43.Onagbesan, O. M. et al. Effects of genotype and feed allowance on plasma luteinizing hormones, follicle-stimulating hormones, progesterone, estradiol levels, follicle differentiation, and egg production rates of broiler breeder hens. Poult. Sci.85, 1245–1258. 10.1093/ps/85.7.1245 (2006). [DOI] [PubMed] [Google Scholar]
- 44.Jia, Y. et al. Melatonin implantation improved the egg-laying rate and quality in hens past their peak egg-laying age. Sci. Rep.6, 1–8. 10.1038/srep39799 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Su, C. H. et al. The effect of different colored light emitting diode illumination on egg laying performance, egg qualities, blood hormone levels and behavior patterns in Brown Tsaiya duck. Anim. Biosci.34, 1870–1878. 10.5713/ab.20.0657 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Beuving, G. & Vonder, G. M. A. Effect of stressing factors on corticosterone levels in the plasma of laying hens. Gen. Comp. Endocrinol.35, 153–159. 10.1016/0016-6480(78)90157-0 (1978). [DOI] [PubMed] [Google Scholar]
- 47.Archer, G. S. How does red light affect layer production, fear, and stress?. Poult. Sci.98, 3–8. 10.3382/ps/pey302 (2019). [DOI] [PubMed] [Google Scholar]
- 48.Sobotik, E. B., Nelson, J. R. & Archer, G. S. How does ultraviolet light affect layer production, fear, and stress. Appl. Anim. Behav. Sci.223, 104926. 10.1016/j.applanim.2019.104926 (2020). [DOI] [PubMed] [Google Scholar]
- 49.House, G. M., Sobotik, E. B., Nelson, J. R. & Archer, G. S. Effect of the addition of ultraviolet light on broiler growth, fear, and stress response. J. Appl. Poult. Res.29, 402–408. 10.1016/j.japr.2020.01.003 (2020). [Google Scholar]
- 50.European Commission. Overview report: Protection of the welfare of laying hens at all stages of production.https://food.ec.europa.eu/document/download/4be5d6ba-0a12-416f-8207-b8a524f89ce9_en?filename=aw_report-offical-controls.pdf (2023).
- 51.Hair, J., Black, W. C., Babin, B. J. & Anderson, R. E. Multivariate Data Analysis 7th ed. 72 (Pearson Educational International, 2010).
- 52.Box, G. E. P. & Cox, D. R. An analysis of transformations. Journal of the Royal Statisistical Society, Series B.26, 211–246. 10.1111/j.2517-6161.1964.tb00553.x (1964). [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
All data analyzed in the study will be shared, as well as statistical analysis plans. The data will become available immediately following publication and with no end date. Access criteria data will be shared (including with anyone who wishes to access the data for any purpose), and proposal should be directed to jgbarros@unicamp.br.