

Abstract
Recently, there has been increasing interest in the complex relationship between transcription and genome stability, with specific attention directed toward the physiological significance of molecular structures known as R‐loops. These structures arise when an RNA strand invades into the DNA duplex, and their formation is involved in a wide range of regulatory functions affecting gene expression, DNA repair processes or cell homeostasis. The persistent presence of R‐loops, if not effectively removed, contributes to genome instability, underscoring the significance of the factors responsible for their resolution and modification. In this review, we provide a comprehensive overview of how R‐loop processing can drive either a beneficial or a harmful outcome. Additionally, we explore the potential for manipulating such structures to devise rationalized therapeutic strategies targeting the aberrant accumulation of R‐loops.
Keywords: DNA damage, DNA repair, genome instability, R‐loops, transcription
R‐loops function as both regulators of gene expression, DNA repair, telomere maintenance or chromosome segregation, and as drivers of genome instability. This dual nature of these structures is coordinated by RNA/DNA/chromatin processing factors that safeguard the genome from detrimental R‐loop‐instigated DNA damage or cellular dysfunction. Understanding R‐loop biology can lead to context‐specific therapeutic interventions targeting their aberrant accumulation in disease.

Abbreviations
AGS, Aicardi Goutières syndrome
AID, activation‐induced cytidine deaminase
ALS, amyotrophic lateral aclerosis
ALS4, amyotrophic lateral sclerosis type 4
ALT, alternative telomere lengthening
Alt‐NHEJ, alternative‐non‐homologous end joining
AML, acute myeloid leukemia
AOA2, ataxia with oculomotor apraxia 2
BER, base excision repair
BRD4, bromodomain‐containing protein 4
CD, co‐directional
cen R‐loops, RNA–DNA hybrids at the centromere
cenRNAs, centromeric long non‐coding RNAs
CPEO, chronic progressive external ophthalmoplegia
CPT, camptothecin
CSR, class‐switch recombination
DDR, DNA damage response
DDRNAs, DNA damage response RNAs
dilncRNAs, damage‐induced long non‐coding RNAs
DSB, double‐strand break
FTD, frontotemporal dementia
FXS, fragile X syndrome
G4, G‐quadruplex
H3K27ac, H3 acetylation
H3K36me3, tri‐methylation of lysine 36 of histone H3
H3K4me1, mono‐methylation of lysine 4 of histone H3
H3K4me3, tri‐methylation of lysine 4 of histone H3
HDR, homology‐directed repair
HO, head‐on
HR, homologous recombination
ICF, immunodeficiency‐centromeric instability‐facial anomalies syndrome
lncRNA, long non‐coding RNA
m 6 A, N6‐methyladenosine
MDS, myelodysplastic syndrome
mESC, mouse embryonic stem cell
MRN, MRE11‐RAD50‐NBS1
NER, nucleotide excision repair
NHEJ, non‐homologous end joining
PRC2, polycomb repression complex 2
ROS, reactive oxygen species
RPA, replication protein A
SETX, senataxin
SMA, spinal muscular atrophy
SSB, single‐strand break
ssDNA, single‐stranded DNA
TC, transcription‐coupled
TERRA, telomere repeat‐containing RNA
TOP1, topoisomerase 1
TRCs, transcription‐replication conflicts
TSSs, transcription start sites
VIM , vimentin gene
XP, xeroderma pigmentosum
Transcription is far from a neutral coordination of proteins toward the mechanical synthesis of RNA; the enzymatic activities involved and the mechanisms that ensure DNA accessibility paradoxically contribute to genome instability, making gene expression a risky endeavor. Besides the inherent mutagenesis associated with transcription [1, 2], DNA breaks induced to relieve RNA polymerase‐associated torsional stress or the risk of transcription‐replication conflicts, a necessary evil that accompanies mRNA synthesis arises from the formation of non‐B DNA structures, such as R‐loops. R‐loops are generated when a transcribed RNA strand invades the DNA duplex and hybridizes with the template DNA, forming an RNA–DNA hybrid and, at the same time, displacing the non‐template DNA as a single‐stranded DNA (ssDNA). Such three‐stranded structures were originally thought to be merely by‐products of transcription, generated exclusively in cis [3]; there is emerging evidence to suggest that they can be formed in trans [4, 5], while they also exhibit regulatory functions. In addition, although R‐loops are instinctively considered to form behind RNA polymerases, there is evidence to suggest that, during backtracking (the backward shift of RNA polymerase along DNA), R‐loops are also generated anterior to the RNA polymerase complex [6]. While short, transient RNA–DNA hybrids form continuously during transcription or DNA synthesis of Okazaki fragments [7], R‐loops are distinct structures that form throughout the cell cycle, span about 100–2000 base pairs and occupy 5–10% of the genome [8, 9]. They have a tendency to form at sites with high GC‐content or GC‐skew [10, 11, 12], a preference which extends their influence beyond highly expressed genes, to encompass repetitive genomic regions, centromeres, telomeres and enhancers [12, 13, 14].
RNA–DNA hybrids and R‐loops have recently attracted attention for having roles in multiple physiological processes, including gene expression regulation and DNA repair [15, 16, 17, 18]. Nevertheless, the failure to promptly and effectively eliminate such structures results in DNA breaks leading to genome instability. For the detrimental effects of R‐loops on genome integrity, we refer the readers to the following reviews [19, 20]. Here, we discuss how processing of co‐transcriptional R‐loops affects the outcome of gene expression and cell homeostasis, with an emphasis on the factors involved in their recognition, modification or removal. We juxtapose the regulatory, beneficial roles of R‐loops, against the detrimental consequences of their misregulation, while also considering the possibility of R‐loop manipulation for therapeutic purposes.
Discerning the characteristics of physiological and harmful R‐loops
Since the earliest evidence of R‐loop formation in vitro, in 1976 [21], there have been numerous studies mapping R‐loops and investigating their biological functions in different organisms [18, 22]. Paradoxically, besides the multiple cases where R‐loop formation is beneficial for fundamental cellular processes, they have also been proven to be deleterious to cells. The distinction between R‐loops with physiological (regulatory) functions, formed either in specific genome loci or at scheduled time‐points (programmed) and spontaneous (unscheduled) R‐loops, which can have detrimental consequences to the cell homeostasis, has raised many questions on their biology [17, 20].
R‐loops occur naturally across the genomes of bacteria, yeast and all higher eukaryotes and are now considered to be more than byproducts of transcription [9, 12]. Comparison of results generated with different R‐loop mapping strategies has revealed major discrepancies, especially in terms of size, sequence characteristics and distribution of R‐loops in the genome. For example, depending on the technique [23, 24, 25, 26, 27, 28], R‐loops are either mapped predominantly along transcribed gene bodies and 3′ gene terminal regions and at hotspots of GC‐skewed promoters, or identified at G‐rich promoters and Transcription Start Sites (TSSs), possibly constricted by the presence of auxiliary endogenous protein factors, such as the ssDNA‐binding protein Replication Protein A (RPA).
Nevertheless, a general consensus emerging from R‐loop mapping approaches is their preference for GC‐rich or ‐skewed genic regions and their transient nature. This dynamic characteristic of R‐loops, besides making the identification of their interacting factors and occupancy sites (e.g. enhancer RNAs have short half‐lives) very challenging, also highlights detrimental implications for genome stability, in the case of other more stable or persistent hybrids, or R‐loops occurring at non‐scheduled times or at aberrant sites [29]. It has been suggested that the threat to genome stability is derived from a secondary event and not the R‐loop itself [20]. This is supported by evidence that describe specific histone mutants that accumulate R‐loops without concurrent DNA damage accumulation [30]. Current research only describes R‐loops as harmful in the context of aberrant genetic modifications and disorders or in the presence of exogenously added agents that influence the transcription process. Although any discernible molecular features of such harmful R‐loops have not been defined yet, a subset of partially RNase H‐resistant RNA–DNA hybrids were recently identified, whose characteristics may well explain the secondary features of an intrinsically (un)stable R‐loop [31].
In principle, the structure of an R‐loop poses a significant threat for genome stability, as the displaced ssDNA strand is open to exogenous and endogenous DNA damaging agents and can be a substrate for nucleases and DNA modifying enzymes. A nick in the DNA can be further processed into a DNA double‐strand break (DSB), making R‐loops a source of DNA damage, while, at the same time, they have the potential to block replication fork progression, causing collisions with the transcription machinery. To this end, cells have developed protective measures to secure the fine balance of R‐loop formation and resolution; RNA export/processing factors [32] and DNA topoisomerases prevent their formation, RNA helicases [33] and RNases remove them, and nucleases [15] promote DNA repair (Table 1).
Table 1.
Protein factors involved in R‐loop regulation and function.
| Molecular function | Protein factor | Associated disease | Proposed mechanism/process | References |
|---|---|---|---|---|
| Topoisomerases | ||||
| Topoisomerase | TOP1, TOP2 | Cancer | Prevention of negative supercoiling, replication stress | [42, 84, 202, 203, 204, 205] |
| Topoisomerase | TOP3B/TDRD3 | Multiple Myeloma Burkhitt's Lymphoma | R‐loop resolution | [179, 206] |
| Nucleases | ||||
| Ribonuclease | RNase H1 | Cancer, Mitochondrial Encephalomyopathy, Chronic Progressive External Ophthalmoplegia (CPEO) | R‐loop resolution | [207, 208, 209] |
| Ribonuclease | RNase H2 | Aicardi‐Goutières Syndrome (AGS) | R‐loop resolution | [154, 210, 211, 212] |
| Ribonuclease | DICER | Cancer, DICER1 syndrome | R‐loop resolution | [57, 213] |
| Ribonuclease | XRN2 | R‐loop resolution, transcription termination | [56, 60, 214, 215] | |
| Endonuclease | XPF, XPG | Xeroderma Pigmentosum (XP), chronic pancreatitis | R‐loop processing | [42, 51, 53] |
| Endonuclease | FEN1 | Lung and gastrointestinal Cancer | Trinucleotide repeat deletion in R‐loops | [216] |
| ssDNA nuclease | SAMHD1 | Aicardi‐Goutières Syndrome (AGS) | R‐loop resolution, tumor suppressor | [155, 212] |
| ssDNA nuclease | TREX1 | Aicardi‐Goutières Syndrome (AGS) | R‐loop resolution | [212, 217] |
| Helicases | ||||
| Helicase | SETX | Amyotrophic Lateral Sclerosis type 4 (ALS4), Ataxia with Oculomotor Apraxia type 2 (AOA2) | R‐loop resolution | [37, 56, 122, 218] |
| Helicase/mRNA splicing | AQR | R‐loop resolution, DSB repair by HR | [53, 219] | |
| Helicase | DHX9 | R‐loop formation/resolution, transcription termination | [58, 220] | |
| Helicase | DDX5 | Cancer | R‐loop resolution, transcription termination | [83, 221] |
| Helicase | DDX21 | Spinal Muscular Atrophy (SMA) | R‐loop resolution | [222, 223] |
| Helicase | DDX17 | Cancer | R‐loop formation/resolution | [33, 224] |
| Helicase | DDX1 | Retinoblastoma, neuroblastoma | R‐loop formation/resolution | [225, 226] |
| Helicase | DDX41 | Myelodysplastic syndrome (MDS), Acute myeloid leukemia (AML) | R‐loop suppression, tumor suppressor | [227] |
| Helicase, DNA repair | BLM | Bloom's syndrome, cancer | R‐loop resolution | [228] |
| Helicase, transcription | RECQL5 | Transcription elongation, R‐loop suppression | [229] | |
| Helicase, exonuclease | WRN | Werner syndrome | R‐loop resolution | [230, 231] |
| Translocase/DNA repair | FANCM | ALT‐dependent cancer | R‐loop resolution | [232] |
| Helicase | RTEL1 | G4 binding on TERRA for R‐loop resolution | [233, 234] | |
| Nucleic acid binding | ||||
| ssDNA binding | AID | Immunodeficiency, cancer | Deaminates cytosine residues in the ssDNA | [235, 236] |
| ssDNA binding | RPA | R‐loop formation/resolution | [46, 47] | |
| ssDNA binding | RAD51 | R‐loop formation | [237, 238] | |
| RNA binding | EWS‐FLI1/EWSR1 | Ewing's sarcoma | R‐loop formation/suppression | [137] |
| DNA/RNA binding | TDP‐43/FUS | Amyotrophic Lateral Sclerosis (ALS), Frontotemporal Dementia (FTD) | R‐loop suppression, replication stress | [239, 240, 241] |
| DNA binding | WASp | Wiskott‐Aldrich syndrome | R‐loop suppression, regulates RPA ssDNA binding | [242, 243] |
| DNA binding, repair | RAD52 | R‐loop processing for DNA repair | [72] | |
| DNA binding, repair | BRCA1/2 | Breast and ovarian Cancer | R‐loop formation/suppression, recruited on R‐loops for DNA repair | [244, 245] |
| DNA binding, repair | FANCA‐W | Fanconi Anemia | R‐loop suppression by ssDNA/RNA binding | [196, 246] |
| RNA processing | ||||
| mRNA processing/export | THO/TREX‐2 complex | Hyperrecombination, sarcoma | Transcription elongation, mRNA export | [35, 244, 247, 248] |
| Splicing factor | U2AF1/2 | Myelodysplastic syndrome (MDS) | Recruited by R‐loops for co‐transcriptional pre‐mRNA splicing | [64, 249] |
| Splicing factor | SRSF1/2 | NASH, Myelodysplastic syndrome (MDS) | R‐loop suppression | [250, 251] |
| RNA editing | ADAR1 | Aicardi‐Goutières syndrome (AGS), cancer | R‐loop regulation by editing A‐C mismatches | [252] |
| Telomere binding, RNA/DNA binding | Shelterin complex | R‐loop formation/suppression, G4 binding on TERRA | [253, 254] | |
| Chromatin remodeling | ||||
| Chromatin remodeler, helicase | ATRX | ALT‐dependent cancer, ATRX syndrome | R‐loop suppression at telomeres | [47, 255] |
| Chromatin remodeler, histone chaperone | FACT complex | Cancer | Nucleosome assembly at R‐loop TRCs | [256] |
| Chromatin remodeler | ARID1A | Cancer | TOP2A‐mediated R‐loop regulation at TRCs | [257] |
| Chromatin remodeler | INO80 | Cancer | R‐loop resolution, chromatin relaxation | [258, 259] |
| Chromatin remodeler | BRG1 | Cancer | R‐loop suppression, regulation of chromatin accessibility | [260] |
| Methyltransferases | ||||
| RNA methyltransferase | METTL3/14, TonEBP | DDR, immunoregulation | R‐loop formation/suppression by m6A methylation | [61, 63] |
| RNA methyltransferase | TRDMT1 | Cancer | R‐loop regulation by m5C methylation | [261] |
| DNA methyltransferase | DNMT3B | Immunodeficiency‐centromeric instability‐facial anomalies syndrome (ICF) | (peri‐)centromeric R‐loop suppression | [262] |
| Other | ||||
| snRNP biogenesis | SMN1 | Spinal Muscular Atrophy (SMA) | Promotes splicing to prevent R‐loops and instability | [263] |
| Poly [ADP‐ribose] polymerase, DNA repair | PARP1 | Cancer | R‐loop suppression | [160] |
| Transcription co‐activator | BRD4/CDK9 | Cancer | R‐loop suppression, transcription elongation at TSS, transcription termination | [164, 213] |
| Helicase‐exoribonuclease complex | SUV3‐PolA‐PNPase | Mitochondrial R‐loop resolution | [264] | |
Intriguingly, the very same structure of R‐loops that makes them a threat to genome integrity is also responsible for their regulatory functions. The first described example of biologically important regulatory R‐loop formation highlights their transcription‐induced contribution in immunoglobulin class‐switch recombination (CSR), during B cell activation [34]. Indeed, it is accepted that high transcription rates increase the likelihood of co‐transcriptional formation of R‐loops [9]. In CSR, R‐loops form co‐transcriptionally at the G‐rich switch region of the IgH locus. Cytosine residues in the displaced non‐template ssDNA are then deaminated by Activation‐Induced cytidine Deaminase (AID), converted to uracils and processed by factors of the Base Excision Repair (BER) and Mismatch Repair pathways, to generate DNA nicks and double‐strand breaks (DSBs), respectively. The production of the different Ig types, vital for the immune response, is only possible through the consequent end joining of these DSBs. Yet, although the connection of RNA polymerase elongation rates with R‐loops suggests that high gene expression levels promote R‐loop formation, in line with rRNA and tRNA loci and multiple context‐specific studies, it is widely established that R‐loops are also potent inhibitors of RNA polymerases, leading to stalling and transcription inhibition [35].
This contradictory dual nature of R‐loops is the focus of ongoing investigation [36], and a major challenge in the field is to dissect the phenotypes that result directly from R‐loops, or from impaired transcription elongation‐associated processes, like mRNA splicing, methylation or export. Besides the specific R‐loop processing events described here, the possibility of a context‐dependent R‐loop biology and structure would be intriguing and could explain their different functions.
Physiological roles of R‐loops during DNA‐templated transactions
Linking R‐loop processing to gene expression outcomes
In addition to their structure, the sequence specificity of RNA–DNA hybrids allows R‐loops to either attract and precisely target proteins on particular chromosomal regions or inhibit the binding of cofactors. For example, R‐loops have been linked to the protection of CpG islands against DNA methylation, both by repelling DNA methyltransferases and by recruiting H3K4 methyltransferases, which results in chromatin decondensation and transcription activation [12, 37, 38]. This renders R‐loops ideal candidates for gene expression and chromatin organization regulation. The fact that such structures occupy ~ 10% of the genome and are found primarily in genic regions, at promoters and termination sites, further supports this role.
R‐loop‐associated transcription activation has been linked to activating histone marks, such as mono‐ and tri‐methylation of lysine 4 of histone H3 (H3K4me1/3) and H3 acetylation (H3K27ac), or to elongation marks, like the tri‐methylation of lysine 36 of histone H3 (H3K36me3) [12, 39]. Moreover, in the case of the vimentin (VIM) gene, transcription activation occurs when the antisense long non‐coding RNA (lncRNA) VIM‐AS1 forms an R‐loop downstream of the VIM TSS, which attracts activators of the NF‐κB pathway [40]. In a similar fashion, the lncRNA TARID‐generated R‐loop recruits the stress response protein GADD45A and the methylcytosine dioxygenase TET1 on the promoter of the TCF21 tumor suppressor gene, for demethylation and transcription activation [41]. Additionally, R‐loop‐directed, CTCF‐mediated DNA looping has recently been shown to induce transcription in mouse embryonic fibroblasts [42]. On the other hand, R‐loops have also been shown to induce transcriptional repression. For instance, in Arabidopsis thaliana, the binding of the ATNDX transcription factor on the displaced ssDNA strand stabilizes an R‐loop on the COOLAIR promoter, resulting in transcription silencing [43]. Similarly, transcription repression of the RASSF1A promoter and a subset of mouse embryonic stem cell (mESC) developmental genes is achieved by the R‐loop‐directed recruitment of the Polycomb Repression Complex 2 (PRC2) [44]. In contrast, a different study has shown that the formation of R‐loops leads to the antagonistic recruitment of the histone acetyl‐transferase complex TIP60‐p400, over PRC2 binding, in promoter regions of mESCs [45]. These results strongly indicate that the role of R‐loops as effectors of epigenetic and transcription regulation depends on the specific context and could be differentially modulated by their stabilization or resolution. Proteins that bind the displaced ssDNA strand of R‐loops may either stabilize the R‐loop structure, as in the case of ATNDX [43], or they can facilitate the consequent recruitment of chromatin‐modifying enzymes or transcription factors. RPA, for example, is required for the stabilization of the ssDNA strand and also promotes R‐loop formation [46], but can also recruit RNase H1 for their resolution [47]. Likewise, AID initiates the resolution and repair of R‐loops in CSR [34]. Interestingly, however, AID deficiency leads to genome‐wide hypermethylation [48], suggesting an additional role in R‐loop‐directed DNA demethylation.
The two structure‐specific endonucleases involved in the transcription‐coupled (TC) nucleotide excision repair (NER) pathway [49, 50], XPF and XPG, have also been identified as R‐loop modifiers [51, 52, 53]. Independently of their primary function in removing transcription‐blocking DNA lesions, these factors are involved in R‐loop processing, leaving single‐strand breaks that can be converted into DSBs, through replication. Although the TC‐NER pathway functions mainly in gene bodies, the two nucleases have been recently shown to be recruited preferentially on promoters, termination regions or imprinted genomic loci, where they facilitate R‐loop‐dependent, CTCF‐mediated DNA looping for transcription activation or silencing [42, 54, 55]. Consistent with their enrichment in G‐rich RNAPII termination sites, especially in gene‐dense regions, R‐loops have also been suggested to assist in transcription termination by stalling RNAPII at the poly‐A sequence. Their resolution ensures the accurate processing and termination of transcripts, preventing transcriptional read‐through [56]. This could be achieved by the recruitment of Senataxin (SETX), an RNA/DNA helicase involved both in transcription termination, through the recruitment of the XRN2 exoribonuclease, and R‐loop resolution. DICER, the only other reported RNA–DNA hybrid resolvase, besides RNases H, has also been suggested to favor the release of RNAPII at termination sites, but it remains to be seen if this function is guided by replication [57]. Additionally, deficiency in the RNA helicases DHX9 and DDX5 has also been shown to impair termination of transcription by RNA polymerases [58, 59, 60]. Recently, the reversible RNA modification N6‐methyladenosine (m6A), which is generated on R‐loops by the METTL3 methyl‐transferase, was shown to facilitate transcription termination [61]. The m6A reader YTHDF2 was additionally demonstrated to be involved in their resolution [62, 63]. Thus, a unified model emerges, wherein R‐loop binding factors may regulate gene expression while, at the same time, ensuring that R‐loops are readily removed to avoid genome instability (Fig. 1A). However, it remains unclear in which genomic contexts these functions are exerted and how a balance in R‐loop numbers is achieved.
Fig. 1.
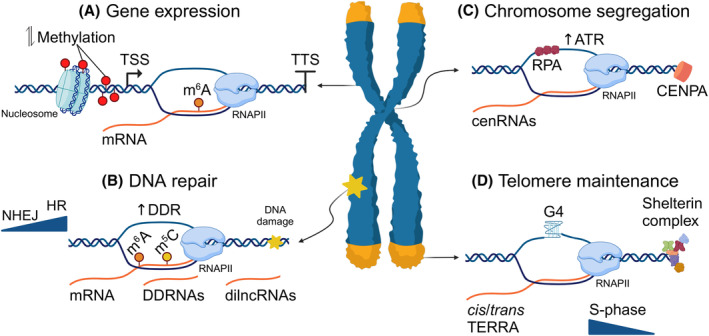
Functions of regulatory R‐loops. (A) R‐loops form at promoters and transcription termination sites and affect transcriptional programs by altering the histone/DNA methylation status or by recruiting chromatin remodelers or transcription factors. (B) R‐loops promote DNA repair by activating the DNA Damage Response, by recruiting protein factors that facilitate repair or by acting as repair intermediates. Pre‐existing or de novo synthesized RNAs (DDRNAs, dilncRNAs) form transient hybrids to guide DNA repair factors to the lesion, while RNA methyl‐modifications either direct the choice of repair pathway or stabilize them to promote repair. (C) Centromeric R‐loops are formed by long non‐coding RNAs transcribed from centromeres (cenRNAs) and facilitate chromosome segregation through an RPA‐dependent activation of ATR. Precise fine‐tuning of cen R‐loops is required for proper centromere assembly (CENPA recruitment) and DSB repair. (D) Telomeres are transcribed into telomere repeat‐containing RNA (TERRA), a G‐rich repetitive long non‐coding RNA which has the propensity for R‐loop formation, necessary for telomere maintenance. TERRA R‐loops are transient, cell cycle‐controlled and are regulated by the shelterin complex. Created with BioRender.com.
R‐loops influence DNA damage response and repair
Genome integrity is not only safeguarded by the timely dissolution of R‐loops; a large body of evidence links the presence of R‐loops and RNA–DNA hybrids directly to DNA damage signaling and repair. By stalling replication forks or promoting DSB formation, R‐loops activate the ATR and ATM protein kinases that orchestrate the response to DNA damage [64, 65, 66, 67]. Additionally, as in CSR, R‐loops act as DNA repair intermediates, activating a non‐canonical ATM‐induced DNA damage response (DDR), with the concurrent retrograde translocation of RNAPII, which allows accessibility for repair, even in the absence of DSBs [68].
Less is clear about the role of R‐loop processing (recognition, editing or resolution) in genome maintenance (Fig. 1B). For instance, the ssDNA‐binding proteins RPA and RAD51 can mediate R‐loop‐induced DNA repair. However, their role could be attributed both to R‐loop binding and to end resection for DSB repair, after R‐loop resolution [69, 70, 71]. Similarly, recruitment of RAD52 to DSBs was shown to be reduced in RNase H‐overexpressing cells [72], while the depletion of SETX favored Non‐Homologous End Joining (NHEJ)‐associated 53BP1 accumulation, over RAD51 recruitment. Yet, evidence that R‐loops enhance or favor Homologous Recombination (HR) is still missing. What seems critical, however, is the impact R‐loops have on DSB end resection, the step which guides the repair choice of HR or NHEJ; contradicting evidence shows R‐loops can either block [73] or enhance [74] DSB resection and, vice‐versa, resection‐associated proteins, such as CtIP, can promote their resolution [75]. Of note, HR is also selected over alternative (Alt‐) NHEJ, when the methyltransferase TRDMT1 preferentially inserts RNA m5C modifications on R‐loops at DSBs [76], while m6A‐modified R‐loops are stabilized to promote DNA repair upon UV irradiation or camptothecin treatment [63].
Another contradicting feature of the involvement of R‐loops in DNA repair is the transcription repression that ensues as a response to DNA breaks. This transcriptional silencing seems at odds with the formation of R‐loops that are generally correlated with high transcription rates. Nevertheless, there is ample evidence to suggest that pre‐existing, or de novo‐synthesized, RNA transcripts are involved in the transient formation of RNA–DNA hybrids, a key step in DSB repair (Fig. 1B). These RNAs (DDRNAs, dilncRNAs) exploit their sequence‐specificity to guide DNA repair factors, such as BRCA1, BRCA2, or RAD51, to the site of the damage [77, 78, 79]. Although some of these R‐loops might be formed by the potential stalling of an elongating RNAPII complex near the site of the damage, it is clear that repair of DNA nicks or DSBs favors R‐loop formation, especially at actively transcribed regions [80]. This seems counter‐intuitive, as one would expect that R‐loop resolution would be required to allow repair. Indeed, RNase H overexpression enhances resection [70, 77], while the helicase activity of several R‐loop processing factors, such as DDX5 or DHX9, has been shown to be promoted in HR [81, 82, 83].
R‐loops in telomere and centromere maintenance
RNA–DNA hybrids preferentially accumulate in telomeres and centromeres [9, 10, 12, 23, 84]. Despite their heterochromatinized nature, centromeres are transcribed by RNAPII into long non‐coding RNAs (cenRNAs) [85, 86, 87]. CenRNAs are synthesized at low levels, in a cell cycle‐dependent manner and can be found in single‐, double‐stranded RNA forms, or in RNA–DNA hybrids at the centromere (cen R‐loops) [39, 88, 89]. The main function of centromeres, the interaction of microtubules with the kinetochore during cell division, is facilitated by cen R‐loops, through the recruitment of RPA and the consequent ATR‐dependent activation of Aurora B [90]. Although cen R‐loops are necessary for chromosome segregation, they are involved in CENP‐A localization [91] and seem to promote end‐resection for DSB repair [92]; their accumulation can be detrimental to genome stability [89] (Fig. 1C). It is tempting to speculate that, from an evolutionary perspective, cen R‐loops offer a significant advantage to proper centromere functionality, especially if one considers that centromeres do not favor their formation; centromeric DNA sequences are mainly AT‐rich [93, 94], while CENP‐A, the centromere‐specific histone H3 variant, introduces positive DNA supercoiling, not suitable to promote R‐loop formation [95, 96, 97].
Similar to centromeres, telomeres are transcribed into telomere repeat‐containing RNA (TERRA), a G‐rich repetitive long non‐coding RNA which has the propensity for R‐loop formation, in cis or in trans (reviewed in [98, 99]). TERRA hybrids are degraded by RNase H enzymes in the S phase, to avoid replication conflicts, while they promote telomere maintenance by driving DNA repair and, as all R‐loops, they are unstable and dynamic. Their transient nature becomes more evident by the different conditions that either increase, reduce or stabilize them, especially considering that TERRA levels are also subject to multiple modifications (Fig. 1D). For example, co‐transcriptional R‐loop accumulation leads to telomere dysfunction [100], while in critically short telomeres, R‐loops are stable throughout the cell cycle and accumulate due to impaired RNase H degradation [101, 102, 103]. This, in turn, promotes HR to prevent premature senescence. Similarly, R‐loops promote telomere length in the tumor cells that maintain their telomeres by the alternative telomere lengthening (ALT) pathway. In fact, homology‐directed repair (HDR) can only be achieved with strictly regulated R‐loop levels, to avoid replication stress or insufficient HDR factor recruitment [13, 104, 105, 106]. The formation of TERRA R‐loops can be counteracted by proteins like the RNA processing THO complex, members of the shelterin complex that determine how TERRA interacts with the DNA, the SWI/SNF2 chromatin remodeler ATRX, or DNA repair factors, such as RAD51, BRCA1 and RAD27.
R‐loops as drivers of genome instability
R‐loops processing in transcription and replication stress
Within transcriptionally active regions, R‐loops can interfere with the DNA repair process by forming physical obstacles to the lesion [107, 108]. The contribution of R‐loop formation on genome instability has been extensively covered in reviews and shall not be discussed here [15, 19, 20, 29, 109]. It is noteworthy that persistent R‐loops can affect nearby gene transcription, altering the chromatin landscape, inducing global changes in transcription programs and contributing to multiple disorders, including neurodegenerative disease and cancer [18, 110, 111]. RNAPII stalling, due to unresolved R‐loops, can also lead to truncated transcripts which, if translated, can yield aberrant proteins [112]. Since RNase H overexpression can suppress these effects, it has been presumed that the source of DNA damage is the R‐loop structure. However, R‐loop formation might simply be an indicator of transcription stress, prompting the argument that stalled/arrested RNAPII compromises genome integrity. In line with this, R‐loops have been shown to be involved in the XRN2‐dependent premature termination of RNAPII, under conditions of transcription stress. This suggests that unresolved R‐loops might represent the actual cause of genome instability (Fig. 2A) [113, 114].
Fig. 2.

R‐loops as a source of genome stability. R‐loops, if left unresolved, can be processed into single‐strand or double‐strand breaks, yet their impact extends to multiple DNA‐templated processes, further contributing to genome instability. (A) R‐loop formation leads to RNAPII stalling, DNA repair inhibition or replication stress. Persistent R‐loop accumulation introduces global gene expression changes and leads to truncated transcripts and the aggregation of aberrant proteins, further adding to transcription stress. (B) Replication forks encountering the transcription machinery in a head‐on (HO) or co‐directional (CD) orientation lead to distinct outcomes. HO conflicts, being more damaging than CD, create favorable conditions for R‐loop formation and DNA damage. (C) R‐loops impede DNA repair and inhibit end resection at DSBs, by physically blocking access to or by interfering with recruitment of repair factors. Created with BioRender.com.
Transcription and replication share the same DNA template and, regardless of the mechanisms to separate them, a progressing replication fork can encounter R‐loops during the S phase [115]. Chromatin‐retained RNAPII due to transcription elongation defects can hinder replication [116], and the impact of R‐loops on replication stress and genome stability has been highlighted either as a source, or a consequence of transcription‐replication conflicts (TRCs) [117]. Evidence from depletion or overexpression of RNase H experiments suggests that R‐loops can interfere with replication fork progression, yet the mechanistic details remain obscure [118, 119, 120]. In line, SETX, the FANCM helicase and RTEL1 have also been associated with R‐loop resolution at TRCs [31, 56, 73, 121, 122, 123, 124, 125]. The complexity arises from the varying effects of replication fork orientation in relation to an R‐loop. It is suggested that conflicts of R‐loops associated with head‐on (HO)‐oriented genes are more harmful, while the co‐directional (CD) encounter of the transcription machinery by the replisome constitutes a smaller impediment [65]. When the replication fork encounters an R‐loop, molecular events diverge depending on the length of the R‐loop, the presence of secondary structures, like G4‐quadruplexes, on the ssDNA strand, or even due to the presence of RNAPII (Fig. 2B) [126, 127, 128]. For example, the convergence of transcription and replication machineries in the HO orientation may induce topological stress, hindering further progression, whereas CD encounters can pose issues if RNA polymerase is stabilized by backtracking. Of note, the RECQL5 helicase mitigates backtracking, thereby reducing TRCs and genome instability [129, 130].
R‐loops hinder DNA repair
As briefly mentioned before, the mechanisms that link R‐loops to DNA repair remain obscure. Nonetheless, there is evidence to suggest a role for R‐loops in DSB end resection. After recognition by the MRE11‐RAD50‐NBS1 (MRN) complex, together with CtIP [131], EXO1/DNA2‐mediated long‐range resection follows, a process requiring the BLM helicase [132, 133]. End resection at induced DSBs can be hindered by improper R‐loop resolution, in the absence of crucial resolution factors (e.g. DDX5, DHX9) that lead to R‐loop accumulation. This results in reduced RPA and RAD51 loading and nuclease recruitment [70, 77, 134], resulting in additional DNA damage [81, 82, 83, 135]. In support of the idea that unscheduled R‐loops inhibit end resection, RNase H/Sen1‐depleted cells accumulated DSBs inhibiting resection at the end of the R‐loop [73, 136]. Hybrid clearance appears crucial at DSB sites, and deregulated hybrids may interfere with resection, causing repair defects and genome instability (Fig. 2C).
Several factors that are implicated in R‐loop resolution also interact with HR proteins, indicating a connection between HR and R‐loop processing. For example, BRCA2 stimulates the helicase activity of DDX5, affecting R‐loop dissolution [82], while the USP42 deubiquitinase facilitates DHX9 and recruits BRCA1 at R‐loops located near DSBs, reinforcing the association between HR and hybrid resolution [81]. Additionally, R‐loops could indirectly hinder DNA repair by titrating out repair proteins, as observed in cells expressing EWS‐FLI, where unscheduled R‐loops prevent BRCA1 relocalization to DSB sites [137].
DNA breaks as a result of R‐loop processing
The NER endonucleases XPF and XPG contribute to DSBs induced by R‐loops, and their depletion results in an increase in cellular R‐loop levels [42, 51, 53, 72, 138]. The mechanism by which XPF and XPG act on R‐loops is not fully understood; they might cleave opposite strands, generating a DSB, or cleave the same strand, generating a single‐strand break (SSB), that is then processed to form a DSB. XPG and XPF can also be recruited to R‐loops by XAB2, indicating a potential role beyond classical NER [52]. Yet, one could consider these XPF/XPG‐induced DSBs not to be a mark of genome instability, but an R‐loop processing intermediate that escapes the repair control. Lastly, it has been described that XPF‐XPG are also involved in the generation of RNA–DNA hybrids or ssDNA fragments, that are actively exported to the cytoplasm of SETX/BRCA1/SAMHD1‐challenged or ERCC1‐deficient cells, respectively, where they induce an inflammatory response [51, 139].
Topoisomerase 1 (TOP1) inhibition also results in R‐loop‐associated DSBs. For instance, the TOP1 inhibitor camptothecin (CPT) induces R‐loops containing trapped TOP1 cleavage complexes, and the endonucleases XPF, XPG, FEN1, and MRE11 that contribute to break formation [138]. Additionally, the suppression of R‐loops by Sae2/CtIP leads to reduced DNA break formation in human cells, while the absence of CtIP and XPG leads to R‐loop accumulation, which is further exacerbated with the addition of CPT [53, 75].
Lastly, cellular metabolic byproducts, such as reactive oxygen species (ROS), and enzymatic activities can cause damage to the exposed single‐stranded DNA (ssDNA) of an R‐loop structure.
R‐loop formation or stabilization is increased specifically in actively transcribed regions, due to ROS‐induced DNA nicks at these sites, consequently resulting in DNA damage [71, 140, 141]. Additionally, cytidine deaminases like APOBEC proteins or AID can contribute to damage of the exposed ssDNA in R‐loops [142, 143, 144], resulting in uracil formation and subsequent single‐strand break formation, as observed in immunoglobulin CSR [143].
R‐loops as putative therapeutic targets
It becomes evident, from the above that R‐loops are an integral part of normal cellular processes. Yet, the need for nuanced regulation of R‐loop formation and resolution, for the efficient execution of their functions, as opposed to driving genome instability, is a recurrent theme in the study of their (patho‐)physiology. It thus seems imperative for cells, for example, to separate the role of R‐loops in facilitating the recruitment of transcription factors and chromatin remodelers, from their involvement in transcription inhibition through RNAPII stalling. Likewise, cells need to fine‐tune the timing of R‐loop formation and subsequent removal. This is in line, for example, with the cell cycle‐dependent regulation of telomere or centromere R‐loops, which fits the DNA replication schedule to avoid transcription‐replication collisions. To do so, cells must be able to determine the exact location or the chromatin environment that favors R‐loop formation, or select which processing factors will be guided to the structures and when. Harnessing this information would provide us with insights on R‐loop manipulation toward rationalized therapeutic interventions.
Indeed, R‐loops have been documented to be associated with multiple diseases, including cancer, neurological syndromes and autoimmune disorders, which have been extensively described in multiple reviews [19, 145, 146, 147, 148, 149, 150]. In cancer, the abnormal accumulation of R‐loops arises from global perturbations in transcription, replication, or RNA processing. The complex interplay between R‐loops and oncogenes or tumor suppressor genes further drives tumorigenesis. Disease progression in several neurodegenerative disorders, such as amyotrophic lateral sclerosis type 4 (ALS4) or ataxia with oculomotor apraxia 2 (AOA2), is also associated with alterations in R‐loop levels, while they also contribute to repeat instability, either through expansion or contraction [37, 110, 124, 151, 152, 153]. Lastly, an immunogenic role has been attributed to R‐loops, especially those associated with the Aicardi Goutières Syndrome (AGS), an autoimmune disorder that leads to progressive neurological damage driven by increased immune signaling [154, 155]. AGS is caused by mutations in a number of genes involved in RNA/DNA processing, including RNase H2, TREX1, SAMHD1 and ADAR1, which eventually lead to the cytoplasmic accumulation of nucleic acid fragments.
As an approach toward combination therapy in oncology, R‐loops have been targeted in order to sensitize cancer cells to chemotherapeutic treatments. Intercalating molecules, such as ethidium bromide [156] or actinomycin D [157] that directly interact with the RNA–DNA hybrids, have been used to inhibit telomerase activity or cell growth [158, 159]. So far, there have been several attempts to exploit R‐loop‐driven DNA damage, toward the induction of cancer cell death. ATR inhibition, for example, sensitizes cells to R‐loop‐induced DNA breaks [66], while PARP inhibition leads to synthetic lethality in cells with impaired HR repair [160]. Moreover, reduced levels of RNF168, the E3 ubiquitin ligase of the R‐loop helicase DHX9, are correlated with a lower tumor incidence and better survival outcomes in patients [161]. The inhibition of RPA has also been shown to induce cytotoxicity and increase the sensitivity of cancer cells to chemotherapy [162]. By impeding the transcription process, Bromodomain‐containing protein 4 (BRD4) inhibition leads to R‐loop‐dependent transcription‐replication collisions, DSBs and cell death [163, 164], while inhibition of pre‐mRNA release leads to elevated R‐loop levels and, interestingly, to the induction of apoptosis [165, 166]. Similarly, G‐quadruplex (G4) stabilizers [167, 168, 169, 170, 171, 172, 173, 174, 175], histone deacetylase inhibitors [176] or compounds that alter R‐loop homeostasis [64, 177, 178, 179, 180] induce R‐loop‐dependent DNA damage and are efficient anticancer treatments. Similarly, selective inhibitory molecules that affect R‐loop homeostasis (e.g. the CARM1 histone arginine methylase inhibitor EZM2302 [181], the PRMT1 histone arginine methylatransferase inhibitor GSK3368715 [179] or spliceosome inhibitors [182]) have been shown to induce cytotoxicity. Putative anticancer drug targets also include topoisomerase 1 [183], RNase H2 [184], SETX [185] or splicing factors [64, 186, 187] whose selective inhibition increases R‐loop levels. In neurodegenerative syndromes like fragile X syndrome (FXS), the therapeutic target is to inhibit R‐loop formation and trinucleotide‐repeat expansion that would lead to gene silencing [188]. Upregulation of R‐loop‐specific helicases, such as SETX, or endonucleases like RNase H1/H2, represents an efficient strategy to reduce R‐loop levels. Alternatively, targeting the resolution of G4 structures that stabilize R‐loops (e.g. in C9ORF72) or promoting the DSB repair processes to counteract the downstream R‐loop effects could prevent the pathology of disease. The RNA–DNA hybrid present in the polypurine tract of HIV‐1 has also been targeted with triple‐helix‐forming oligonucleotides [189] or small intercalating molecules [190], in order to inhibit reverse transcription of the virus RNA. Accordingly, G4 ligands have been used as antiviral therapy or to target telomeric R‐loops [191, 192, 193].
Exploring the therapeutic potential of R‐loop manipulation, while conceptually straightforward, faces certain pitfalls. Similarly to their regulatory functions, the adverse effects of R‐loops also seem context‐dependent. In this sense, any efforts to forcefully eliminate them would disrupt the tight balance of their context‐specific functions and lead to perturbations of cellular homeostasis. Of note, the indirect targeting of R‐loops through their associated factors brings more complexity to the end result. For example, RNase H1 overexpression, besides being toxic [194, 195, 196, 197, 198, 199], can alter the stability of a number of DDR factors [199], or impact the outcome of transcription programs [200, 201]. Presumably, the unwinding capacity of helicases could prove more beneficial over the loss of nascent RNA transcripts. Moreover, the cell‐cycle‐regulated delivery of RPA‐independent RNase H1 molecules could enhance efficiency. Utilizing the sequence specificity of R‐loops in specific contexts, such as employing guide RNAs, would offer a more targeted approach. Overall, the therapeutic potential of targeting R‐loops is promising, with strategies including inhibition of R‐loop formation and promotion of their resolution offering avenues for intervention (Fig. 3). However, the complex nature of R‐loop biology presents challenges, as their dual role as both regulators and sources of genome instability must be carefully considered. Context‐specific approaches will be essential to minimize off‐target effects and maintain cellular homeostasis while harnessing the therapeutic potential of R‐loop manipulation.
Fig. 3.

Unlocking the dual nature of R‐loops. R‐loops function as both regulators of gene expression, DNA repair, telomere maintenance or chromosome segregation, and as drivers of genome instability. This inherent dual nature of these structures is coordinated by RNA/DNA/chromatin processing factors that safeguard the genome from detrimental R‐loop‐instigated DNA damage or cellular dysfunction. The association of R‐loops with multiple diseases, including cancer, neurodegenerative or autoimmune disorders, underscores their clinical relevance. Therapeutic strategies targeting R‐loop manipulation show promise, with approaches ranging from promoting (e.g. fragile X syndrome) to inhibiting their resolution (e.g. cancer). However, the context‐dependent nature of R‐loops necessitates careful consideration to avoid disrupting cellular homeostasis. Future research aimed at understanding the intricacies of R‐loop biology and developing context‐specific interventions will be crucial for realizing their therapeutic potential while minimizing adverse effects. Created with BioRender.com.
Concluding remarks
Research on R‐loops, the three‐stranded nucleic acid structures formed by the hybridization of RNA to the DNA template, has seen significant interest due to the multitude of cellular processes that are affected by their presence. Inspiring work in the past years highlights their inherent duality as both regulators of gene expression, DNA repair, telomere maintenance or chromosome segregation, and as a potent source of DNA damage and genome instability (Fig. 3). Each role depends heavily on the genomic context and time of formation, but also, equally, on the distinct protein factors that mediate their resolution. Although our knowledge of the R‐loops' impact on genome stability is increasing, so much so that they could even be brazenly considered as a DNA damage marker, many questions remain on what makes an R‐loop to be harmful. Their DNA content/sequence definitely adds to this complexity, especially when it modifies their accessibility potential, while the surrounding torsional or epigenetic status of chromatin seems key to their functional outcome. Yet, from an evolutionary perspective, one can't help but ponder the causality of R‐loop formation. For example, is their transcription termination function general for all, or at least a specific subset of genes? Is the choice of HR over NHEJ repair affected by R‐loop presence and could this be a characteristic of actively transcribed loci? And, vice‐versa, could we extrapolate information from R‐loop features (e.g., RNA–DNA hybridization or secondary structures) to be necessary for all DNA looping events? Continuing research over such questions will introduce an exciting number of opportunities toward the understanding of R‐loop physiology and regulation. By navigating the complexities of R‐loop biology, we can unlock new possibilities for developing targeted therapeutic interventions, to address a wide range of diseases and improve human health.
Author contributions
KS and AS wrote the manuscript; GAG edited the manuscript.
Acknowledgements
We apologize to all colleagues whose work could not be discussed due to space constraints. The Horizon 2020 Marie Curie ITN “aDDRess” (GA 812829), and “HealthAge” (GA 812830), ELIDEK grants 15546; Fondation Sante; The “Research‐Create‐Innovate” actions (MIA‐RTDI) “Panther”‐00852; Uni‐Pharma Kleon Tsetis Pharmaceutical Laboratories S.A. (PAR00838) and Pharmathen S.A. (PAR00863) funds; and the European Union‐Next Generation EU, Greece 2.0 National Recovery and Resilience plan, National Flagship Initiative “Health and Pharmaceuticals” TAEDR‐0535850 supported this work.
Kalliopi Stratigi and Athanasios Siametis contributed equally to this article
Edited by Donata Orioli
References
- 1. Aguilera A (2002) The connection between transcription and genomic instability. EMBO J 21, 195–201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. Gaillard H and Aguilera A (2016) Transcription as a threat to genome integrity. Annu Rev Biochem 85, 291–317. [DOI] [PubMed] [Google Scholar]
- 3. Aguilera A and Garcia‐Muse T (2012) R loops: from transcription byproducts to threats to genome stability. Mol Cell 46, 115–124. [DOI] [PubMed] [Google Scholar]
- 4. Knott GJ and Doudna JA (2018) CRISPR‐Cas guides the future of genetic engineering. Science 361, 866–869. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Wahba L, Gore SK and Koshland D (2013) The homologous recombination machinery modulates the formation of RNA‐DNA hybrids and associated chromosome instability. Elife 2, e00505. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Zatreanu D, Han Z, Mitter R, Tumini E, Williams H, Gregersen L, Dirac‐Svejstrup AB, Roma S, Stewart A, Aguilera A et al. (2019) Elongation factor TFIIS prevents transcription stress and R‐loop accumulation to maintain genome stability. Mol Cell 76, 57–69.e9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Burgers PM (2019) Solution to the 50‐year‐old Okazaki‐fragment problem. Proc Natl Acad Sci USA 116, 3358–3360. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Sanz LA, Hartono SR, Lim YW, Steyaert S, Rajpurkar A, Ginno PA, Xu X and Chedin F (2016) Prevalent, dynamic, and conserved R‐loop structures associate with specific Epigenomic signatures in mammals. Mol Cell 63, 167–178. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Wahba L, Costantino L, Tan FJ, Zimmer A and Koshland D (2016) S1‐DRIP‐seq identifies high expression and polyA tracts as major contributors to R‐loop formation. Genes Dev 30, 1327–1338. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Chan YA, Aristizabal MJ, Lu PY, Luo Z, Hamza A, Kobor MS, Stirling PC and Hieter P (2014) Genome‐wide profiling of yeast DNA:RNA hybrid prone sites with DRIP‐chip. PLoS Genet 10, e1004288. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Ginno PA, Lim YW, Lott PL, Korf I and Chedin F (2013) GC skew at the 5′ and 3′ ends of human genes links R‐loop formation to epigenetic regulation and transcription termination. Genome Res 23, 1590–1600. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Ginno PA, Lott PL, Christensen HC, Korf I and Chedin F (2012) R‐loop formation is a distinctive characteristic of unmethylated human CpG Island promoters. Mol Cell 45, 814–825. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Arora R, Lee Y, Wischnewski H, Brun CM, Schwarz T and Azzalin CM (2014) RNaseH1 regulates TERRA‐telomeric DNA hybrids and telomere maintenance in ALT tumour cells. Nat Commun 5, 5220. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Nadel J, Athanasiadou R, Lemetre C, Wijetunga NA, Broin PÓ, Sato H, Zhang Z, Jeddeloh J, Montagna C, Golden A et al. (2015) RNA:DNA hybrids in the human genome have distinctive nucleotide characteristics, chromatin composition, and transcriptional relationships. Epigenetics Chromatin 8, 46. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Brickner JR, Garzon JL and Cimprich KA (2022) Walking a tightrope: the complex balancing act of R‐loops in genome stability. Mol Cell 82, 2267–2297. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Garcia‐Muse T and Aguilera A (2019) R loops: from physiological to pathological roles. Cell 179, 604–618. [DOI] [PubMed] [Google Scholar]
- 17. Niehrs C and Luke B (2020) Regulatory R‐loops as facilitators of gene expression and genome stability. Nat Rev Mol Cell Biol 21, 167–178. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Petermann E, Lan L and Zou L (2022) Sources, resolution and physiological relevance of R‐loops and RNA‐DNA hybrids. Nat Rev Mol Cell Biol 23, 521–540. [DOI] [PubMed] [Google Scholar]
- 19. Crossley MP, Bocek M and Cimprich KA (2019) R‐loops as cellular regulators and genomic threats. Mol Cell 73, 398–411. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Rondon AG and Aguilera A (2019) What causes an RNA‐DNA hybrid to compromise genome integrity? DNA Repair (Amst) 81, 102660. [DOI] [PubMed] [Google Scholar]
- 21. Thomas M, White RL and Davis RW (1976) Hybridization of RNA to double‐stranded DNA: formation of R‐loops. Proc Natl Acad Sci USA 73, 2294–2298. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Hegazy YA, Fernando CM and Tran EJ (2020) The balancing act of R‐loop biology: the good, the bad, and the ugly. J Biol Chem 295, 905–913. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Chen L, Chen JY, Zhang X, Gu Y, Xiao R, Shao C, Tang P, Qian H, Luo D, Li H et al. (2017) R‐ChIP using inactive RNase H reveals dynamic coupling of R‐loops with transcriptional pausing at gene promoters. Mol Cell 68, 745–757.e5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Crossley MP, Bocek MJ, Hamperl S, Swigut T and Cimprich KA (2020) qDRIP: a method to quantitatively assess RNA‐DNA hybrid formation genome‐wide. Nucleic Acids Res 48, e84. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Dumelie JG and Jaffrey SR (2017) Defining the location of promoter‐associated R‐loops at near‐nucleotide resolution using bisDRIP‐seq. Elife 6, e28306. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Malig M, Hartono SR, Giafaglione JM, Sanz LA and Chedin F (2020) Ultra‐deep coverage single‐molecule R‐loop Footprinting reveals principles of R‐loop formation. J Mol Biol 432, 2271–2288. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Wang K, Wang H, Li C, Yin Z, Xiao R, Li Q, Xiang Y, Wang W, Huang J, Chen L et al. (2021) Genomic profiling of native R loops with a DNA‐RNA hybrid recognition sensor. Sci Adv 7, eabe3516. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Yan Q, Shields EJ, Bonasio R and Sarma K (2019) Mapping native R‐loops genome‐wide using a targeted nuclease approach. Cell Rep 29, 1369–1380.e5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Rinaldi C, Pizzul P, Longhese MP and Bonetti D (2020) Sensing R‐loop‐associated DNA damage to safeguard genome stability. Front Cell Dev Biol 8, 618157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Garcia‐Pichardo D, Canas JC, Garcia‐Rubio ML, Gomez‐Gonzalez B, Rondon AG and Aguilera A (2017) Histone mutants separate R loop formation from genome instability induction. Mol Cell 66, 597–609.e5. [DOI] [PubMed] [Google Scholar]
- 31. Arnould C, Rocher V, Saur F, Bader AS, Muzzopappa F, Collins S, Lesage E, Le Bozec B, Puget N, Clouaire T et al. (2023) Chromatin compartmentalization regulates the response to DNA damage. Nature 623, 183–192. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Brown RE, Su XA, Fair S, Wu K, Verra L, Jong R, Andrykovich K and Freudenreich CH (2022) The RNA export and RNA decay complexes THO and TRAMP prevent transcription‐replication conflicts, DNA breaks, and CAG repeat contractions. PLoS Biol 20, e3001940. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Yang S, Winstone L, Mondal S and Wu Y (2023) Helicases in R‐loop formation and resolution. J Biol Chem 299, 105307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Chaudhuri J, Tian M, Khuong C, Chua K, Pinaud E and Alt FW (2003) Transcription‐targeted DNA deamination by the AID antibody diversification enzyme. Nature 422, 726–730. [DOI] [PubMed] [Google Scholar]
- 35. Huertas P and Aguilera A (2003) Cotranscriptionally formed DNA:RNA hybrids mediate transcription elongation impairment and transcription‐associated recombination. Mol Cell 12, 711–721. [DOI] [PubMed] [Google Scholar]
- 36. Bonnet A, Grosso AR, Elkaoutari A, Coleno E, Presle A, Sridhara SC, Janbon G, Geli V, de Almeida SF and Palancade B (2017) Introns protect eukaryotic genomes from transcription‐associated genetic instability. Mol Cell 67, 608–621.e6. [DOI] [PubMed] [Google Scholar]
- 37. Grunseich C, Wang IX, Watts JA, Burdick JT, Guber RD, Zhu Z, Bruzel A, Lanman T, Chen K, Schindler AB et al. (2018) Senataxin mutation reveals how R‐loops promote transcription by blocking DNA methylation at gene promoters. Mol Cell 69, 426–437.e7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Krajewski WA, Nakamura T, Mazo A and Canaani E (2005) A motif within SET‐domain proteins binds single‐stranded nucleic acids and transcribed and supercoiled DNAs and can interfere with assembly of nucleosomes. Mol Cell Biol 25, 1891–1899. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Castellano‐Pozo M, Santos‐Pereira JM, Rondon AG, Barroso S, Andujar E, Perez‐Alegre M, Garcia‐Muse T and Aguilera A (2013) R loops are linked to histone H3 S10 phosphorylation and chromatin condensation. Mol Cell 52, 583–590. [DOI] [PubMed] [Google Scholar]
- 40. Boque‐Sastre R, Soler M, Oliveira‐Mateos C, Portela A, Moutinho C, Sayols S, Villanueva A, Esteller M and Guil S (2015) Head‐to‐head antisense transcription and R‐loop formation promotes transcriptional activation. Proc Natl Acad Sci USA 112, 5785–5790. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Arab K, Karaulanov E, Musheev M, Trnka P, Schafer A, Grummt I and Niehrs C (2019) GADD45A binds R‐loops and recruits TET1 to CpG Island promoters. Nat Genet 51, 217–223. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Chatzinikolaou G, Stratigi K, Siametis A, Goulielmaki E, Akalestou‐Clocher A, Tsamardinos I, Topalis P, Austin C, Bouwman BAM, Crosetto N et al. (2023) XPF interacts with TOP2B for R‐loop processing and DNA looping on actively transcribed genes. Sci Adv 9, eadi2095. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Sun Q, Csorba T, Skourti‐Stathaki K, Proudfoot NJ and Dean C (2013) R‐loop stabilization represses antisense transcription at the Arabidopsis FLC locus. Science 340, 619–621. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44. Beckedorff FC, Ayupe AC, Crocci‐Souza R, Amaral MS, Nakaya HI, Soltys DT, Menck CF, Reis EM and Verjovski‐Almeida S (2013) The intronic long noncoding RNA ANRASSF1 recruits PRC2 to the RASSF1A promoter, reducing the expression of RASSF1A and increasing cell proliferation. PLoS Genet 9, e1003705. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. Chen PB, Chen HV, Acharya D, Rando OJ and Fazzio TG (2015) R loops regulate promoter‐proximal chromatin architecture and cellular differentiation. Nat Struct Mol Biol 22, 999–1007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Mazina OM, Somarowthu S, Kadyrova LY, Baranovskiy AG, Tahirov TH, Kadyrov FA and Mazin AV (2020) Replication protein a binds RNA and promotes R‐loop formation. J Biol Chem 295, 14203–14213. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47. Nguyen HD, Yadav T, Giri S, Saez B, Graubert TA and Zou L (2017) Functions of replication protein a as a sensor of R loops and a regulator of RNaseH1. Mol Cell 65, 832–847.e4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48. Popp C, Dean W, Feng S, Cokus SJ, Andrews S, Pellegrini M, Jacobsen SE and Reik W (2010) Genome‐wide erasure of DNA methylation in mouse primordial germ cells is affected by AID deficiency. Nature 463, 1101–1105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49. Kamileri I, Karakasilioti I and Garinis GA (2012) Nucleotide excision repair: new tricks with old bricks. Trends Genet 28, 566–573. [DOI] [PubMed] [Google Scholar]
- 50. Apostolou Z, Chatzinikolaou G, Stratigi K and Garinis GA (2019) Nucleotide excision repair and transcription‐associated genome instability. Bioessays 41, e1800201. [DOI] [PubMed] [Google Scholar]
- 51. Chatzidoukaki O, Stratigi K, Goulielmaki E, Niotis G, Akalestou‐Clocher A, Gkirtzimanaki K, Zafeiropoulos A, Altmuller J, Topalis P and Garinis GA (2021) R‐loops trigger the release of cytoplasmic ssDNAs leading to chronic inflammation upon DNA damage. Sci Adv 7, eabj5769. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52. Goulielmaki E, Tsekrekou M, Batsiotos N, Ascensao‐Ferreira M, Ledaki E, Stratigi K, Chatzinikolaou G, Topalis P, Kosteas T, Altmuller J et al. (2021) The splicing factor XAB2 interacts with ERCC1‐XPF and XPG for R‐loop processing. Nat Commun 12, 3153. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53. Sollier J, Stork CT, Garcia‐Rubio ML, Paulsen RD, Aguilera A and Cimprich KA (2014) Transcription‐coupled nucleotide excision repair factors promote R‐loop‐induced genome instability. Mol Cell 56, 777–785. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54. Kamileri I, Karakasilioti I, Sideri A, Kosteas T, Tatarakis A, Talianidis I and Garinis GA (2012) Defective transcription initiation causes postnatal growth failure in a mouse model of nucleotide excision repair (NER) progeria. Proc Natl Acad Sci USA 109, 2995–3000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55. Chatzinikolaou G, Apostolou Z, Aid‐Pavlidis T, Ioannidou A, Karakasilioti I, Papadopoulos GL, Aivaliotis M, Tsekrekou M, Strouboulis J, Kosteas T et al. (2017) ERCC1‐XPF cooperates with CTCF and cohesin to facilitate the developmental silencing of imprinted genes. Nat Cell Biol 19, 421–432. [DOI] [PubMed] [Google Scholar]
- 56. Skourti‐Stathaki K, Proudfoot NJ and Gromak N (2011) Human senataxin resolves RNA/DNA hybrids formed at transcriptional pause sites to promote Xrn2‐dependent termination. Mol Cell 42, 794–805. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57. Camino LP, Dutta A, Barroso S, Perez‐Calero C, Katz JN, Garcia‐Rubio M, Sung P, Gomez‐Gonzalez B and Aguilera A (2023) DICER ribonuclease removes harmful R‐loops. Mol Cell 83, 3707–3719.e5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58. Cristini A, Groh M, Kristiansen MS and Gromak N (2018) RNA/DNA hybrid Interactome identifies DXH9 as a molecular player in transcriptional termination and R‐loop‐associated DNA damage. Cell Rep 23, 1891–1905. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59. Polenkowski M, Allister AB, Burbano de Lara S, Pierce A, Geary B, El Bounkari O, Wiehlmann L, Hoffmann A, Whetton AD, Tamura T et al. (2023) THOC5 complexes with DDX5, DDX17, and CDK12 to regulate R loop structures and transcription elongation rate. iScience 26, 105784. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60. Villarreal OD, Mersaoui SY, Yu Z, Masson JY and Richard S (2020) Genome‐wide R‐loop analysis defines unique roles for DDX5, XRN2, and PRMT5 in DNA/RNA hybrid resolution. Life Sci Alliance 3, e202000762. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61. Yang X, Liu QL, Xu W, Zhang YC, Yang Y, Ju LF, Chen J, Chen YS, Li K, Ren J et al. (2019) M(6)a promotes R‐loop formation to facilitate transcription termination. Cell Res 29, 1035–1038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62. Abakir A, Giles TC, Cristini A, Foster JM, Dai N, Starczak M, Rubio‐Roldan A, Li M, Eleftheriou M, Crutchley J et al. (2020) N(6)‐methyladenosine regulates the stability of RNA:DNA hybrids in human cells. Nat Genet 52, 48–55. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63. Kang HJ, Cheon NY, Park H, Jeong GW, Ye BJ, Yoo EJ, Lee JH, Hur JH, Lee EA, Kim H et al. (2021) TonEBP recognizes R‐loops and initiates m6A RNA methylation for R‐loop resolution. Nucleic Acids Res 49, 269–284. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64. Chen L, Chen JY, Huang YJ, Gu Y, Qiu J, Qian H, Shao C, Zhang X, Hu J, Li H et al. (2018) The augmented R‐loop is a unifying mechanism for myelodysplastic syndromes induced by high‐risk splicing factor mutations. Mol Cell 69, 412–425.e6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65. Hamperl S, Bocek MJ, Saldivar JC, Swigut T and Cimprich KA (2017) Transcription‐replication conflict orientation modulates R‐loop levels and activates distinct DNA damage responses. Cell 170, 774–786.e19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66. Matos DA, Zhang JM, Ouyang J, Nguyen HD, Genois MM and Zou L (2020) ATR protects the genome against R loops through a MUS81‐triggered feedback loop. Mol Cell 77, 514–527.e4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67. Sollier J and Cimprich KA (2015) Breaking bad: R‐loops and genome integrity. Trends Cell Biol 25, 514–522. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68. Tresini M, Warmerdam DO, Kolovos P, Snijder L, Vrouwe MG, Demmers JA, van IJckenvan WFJ, Grosveld FG, Medema RH, Hoeijmakers JH et al. (2015) The core spliceosome as target and effector of non‐canonical ATM signalling. Nature 523, 53–58. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69. Cohen S, Puget N, Lin YL, Clouaire T, Aguirrebengoa M, Rocher V, Pasero P, Canitrot Y and Legube G (2018) Senataxin resolves RNA:DNA hybrids forming at DNA double‐strand breaks to prevent translocations. Nat Commun 9, 533. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70. Ohle C, Tesorero R, Schermann G, Dobrev N, Sinning I and Fischer T (2016) Transient RNA‐DNA hybrids are required for efficient Double‐Strand break repair. Cell 167, 1001–1013.e7. [DOI] [PubMed] [Google Scholar]
- 71. Teng Y, Yadav T, Duan M, Tan J, Xiang Y, Gao B, Xu J, Liang Z, Liu Y, Nakajima S et al. (2018) ROS‐induced R loops trigger a transcription‐coupled but BRCA1/2‐independent homologous recombination pathway through CSB. Nat Commun 9, 4115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72. Yasuhara T, Kato R, Hagiwara Y, Shiotani B, Yamauchi M, Nakada S, Shibata A and Miyagawa K (2018) Human Rad52 promotes XPG‐mediated R‐loop processing to initiate transcription‐associated homologous recombination repair. Cell 175, 558–570.e11. [DOI] [PubMed] [Google Scholar]
- 73. Costantino L and Koshland D (2018) Genome‐wide map of R‐loop‐induced damage reveals how a subset of R‐loops contributes to genomic instability. Mol Cell 71, 487–497.e3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74. Lu WT, Hawley BR, Skalka GL, Baldock RA, Smith EM, Bader AS, Malewicz M, Watts FZ, Wilczynska A and Bushell M (2018) Drosha drives the formation of DNA:RNA hybrids around DNA break sites to facilitate DNA repair. Nat Commun 9, 532. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75. Makharashvili N, Arora S, Yin Y, Fu Q, Wen X, Lee JH, Kao CH, Leung JW, Miller KM and Paull TT (2018) Sae2/CtIP prevents R‐loop accumulation in eukaryotic cells. Elife 7, e42733. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76. Yang S, Zhou D, Zhang C, Xiang J and Xi X (2023) Function of m(5)C RNA methyltransferase NOP2 in high‐grade serous ovarian cancer. Cancer Biol Ther 24, 2263921. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77. D'Alessandro G, Whelan DR, Howard SM, Vitelli V, Renaudin X, Adamowicz M, Iannelli F, Jones‐Weinert CW, Lee M, Matti V et al. (2018) BRCA2 controls DNA:RNA hybrid level at DSBs by mediating RNase H2 recruitment. Nat Commun 9, 5376. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78. Michelini F, Pitchiaya S, Vitelli V, Sharma S, Gioia U, Pessina F, Cabrini M, Wang Y, Capozzo I, Iannelli F et al. (2017) Damage‐induced lncRNAs control the DNA damage response through interaction with DDRNAs at individual double‐strand breaks. Nat Cell Biol 19, 1400–1411. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79. San Martin‐Alonso M, Soler‐Oliva ME, Garcia‐Rubio M, Garcia‐Muse T and Aguilera A (2021) Harmful R‐loops are prevented via different cell cycle‐specific mechanisms. Nat Commun 12, 4451. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80. Ortega P, Merida‐Cerro JA, Rondon AG, Gomez‐Gonzalez B and Aguilera A (2021) DNA‐RNA hybrids at DSBs interfere with repair by homologous recombination. Elife 10, e69881. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81. Matsui M, Sakasai R, Abe M, Kimura Y, Kajita S, Torii W, Katsuki Y, Ishiai M, Iwabuchi K, Takata M et al. (2020) USP42 enhances homologous recombination repair by promoting R‐loop resolution with a DNA‐RNA helicase DHX9. Oncogenesis 9, 60. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82. Sessa G, Gomez‐Gonzalez B, Silva S, Perez‐Calero C, Beaurepere R, Barroso S, Martineau S, Martin C, Ehlen A, Martinez JS et al. (2021) BRCA2 promotes DNA‐RNA hybrid resolution by DDX5 helicase at DNA breaks to facilitate their repairdouble dagger. EMBO J 40, e106018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83. Yu Z, Mersaoui SY, Guitton‐Sert L, Coulombe Y, Song J, Masson JY and Richard S (2020) DDX5 resolves R‐loops at DNA double‐strand breaks to promote DNA repair and avoid chromosomal deletions. NAR Cancer 2, zcaa028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84. El Hage A, Webb S, Kerr A and Tollervey D (2014) Genome‐wide distribution of RNA‐DNA hybrids identifies RNase H targets in tRNA genes, retrotransposons and mitochondria. PLoS Genet 10, e1004716. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85. Ferri F, Bouzinba‐Segard H, Velasco G, Hube F and Francastel C (2009) Non‐coding murine centromeric transcripts associate with and potentiate Aurora B kinase. Nucleic Acids Res 37, 5071–5080. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86. Ohkuni K and Kitagawa K (2011) Endogenous transcription at the centromere facilitates centromere activity in budding yeast. Curr Biol 21, 1695–1703. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87. Wong LH, Brettingham‐Moore KH, Chan L, Quach JM, Anderson MA, Northrop EL, Hannan R, Saffery R, Shaw ML, Williams E et al. (2007) Centromere RNA is a key component for the assembly of nucleoproteins at the nucleolus and centromere. Genome Res 17, 1146–1160. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88. Liu Q, Liu Y, Shi Q, Su H, Wang C, Birchler JA and Han F (2021) Emerging roles of centromeric RNAs in centromere formation and function. Genes Genomics 43, 217–226. [DOI] [PubMed] [Google Scholar]
- 89. Mishra PK, Chakraborty A, Yeh E, Feng W, Bloom KS and Basrai MA (2021) R‐loops at centromeric chromatin contribute to defects in kinetochore integrity and chromosomal instability in budding yeast. Mol Biol Cell 32, 74–89. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90. Kabeche L, Nguyen HD, Buisson R and Zou L (2018) A mitosis‐specific and R loop‐driven ATR pathway promotes faithful chromosome segregation. Science 359, 108–114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91. Racca C, Britton S, Hedouin S, Francastel C, Calsou P and Larminat F (2021) BRCA1 prevents R‐loop‐associated centromeric instability. Cell Death Dis 12, 896. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92. Yilmaz D, Furst A, Meaburn K, Lezaja A, Wen Y, Altmeyer M, Reina‐San‐Martin B and Soutoglou E (2021) Activation of homologous recombination in G1 preserves centromeric integrity. Nature 600, 748–753. [DOI] [PubMed] [Google Scholar]
- 93. Altemose N, Logsdon GA, Bzikadze AV, Sidhwani P, Langley SA, Caldas GV, Hoyt SJ, Uralsky L, Ryabov FD, Shew CJ et al. (2022) Complete genomic and epigenetic maps of human centromeres. Science 376, eabl4178. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94. Baker RE and Rogers K (2005) Genetic and genomic analysis of the AT‐rich centromere DNA element II of Saccharomyces cerevisiae . Genetics 171, 1463–1475. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95. Furuyama T and Henikoff S (2009) Centromeric nucleosomes induce positive DNA supercoils. Cell 138, 104–113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96. Guo MS, Kawamura R, Littlehale ML, Marko JF and Laub MT (2021) High‐resolution, genome‐wide mapping of positive supercoiling in chromosomes. Elife 10, e67236. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97. Stolz R, Sulthana S, Hartono SR, Malig M, Benham CJ and Chedin F (2019) Interplay between DNA sequence and negative superhelicity drives R‐loop structures. Proc Natl Acad Sci USA 116, 6260–6269. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98. Fernandes RV, Feretzaki M and Lingner J (2021) The makings of TERRA R‐loops at chromosome ends. Cell Cycle 20, 1745–1759. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99. Glousker G, Fernandes RV, Feretzaki M and Lingner J (2022) Detection of TERRA R‐loops at human telomeres. Methods Mol Biol 2528, 159–171. [DOI] [PubMed] [Google Scholar]
- 100. Siametis A, Stratigi K, Giamaki D, Chatzinikolaou G, Akalestou‐Clocher A, Goulielmaki E, Luke B, Schumacher B and Garinis GA (2024) Transcription stress at telomeres leads to cytosolic DNA release and paracrine senescence. Nat Commun 15, 4061. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101. Cusanelli E, Romero CA and Chartrand P (2013) Telomeric noncoding RNA TERRA is induced by telomere shortening to nucleate telomerase molecules at short telomeres. Mol Cell 51, 780–791. [DOI] [PubMed] [Google Scholar]
- 102. Graf M, Bonetti D, Lockhart A, Serhal K, Kellner V, Maicher A, Jolivet P, Teixeira MT and Luke B (2017) Telomere length determines TERRA and R‐loop regulation through the cell cycle. Cell 170, 72–85.e14. [DOI] [PubMed] [Google Scholar]
- 103. Pires VB, Lohner N, Wagner T, Wagner CB, Wilkens M, Hajikazemi M, Paeschke K, Butter F and Luke B (2023) RNA‐DNA hybrids prevent resection at dysfunctional telomeres. Cell Rep 42, 112077. [DOI] [PubMed] [Google Scholar]
- 104. Balk B, Dees M, Bender K and Luke B (2014) The differential processing of telomeres in response to increased telomeric transcription and RNA‐DNA hybrid accumulation. RNA Biol 11, 95–100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105. Perez‐Martinez L, Ozturk M, Butter F and Luke B (2020) Npl3 stabilizes R‐loops at telomeres to prevent accelerated replicative senescence. EMBO Rep 21, e49087. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106. Silva B, Arora R, Bione S and Azzalin CM (2021) TERRA transcription destabilizes telomere integrity to initiate break‐induced replication in human ALT cells. Nat Commun 12, 3760. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107. Liu S, Hua Y, Wang J, Li L, Yuan J, Zhang B, Wang Z, Ji J and Kong D (2021) RNA polymerase III is required for the repair of DNA double‐strand breaks by homologous recombination. Cell 184, 1314–1329.e10. [DOI] [PubMed] [Google Scholar]
- 108. Pessina F, Giavazzi F, Yin Y, Gioia U, Vitelli V, Galbiati A, Barozzi S, Garre M, Oldani A, Flaus A et al. (2019) Functional transcription promoters at DNA double‐strand breaks mediate RNA‐driven phase separation of damage‐response factors. Nat Cell Biol 21, 1286–1299. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109. Marnef A and Legube G (2021) R‐loops as Janus‐faced modulators of DNA repair. Nat Cell Biol 23, 305–313. [DOI] [PubMed] [Google Scholar]
- 110. Groh M and Gromak N (2014) Out of balance: R‐loops in human disease. PLoS Genet 10, e1004630. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111. Li F, Zafar A, Luo L, Denning AM, Gu J, Bennett A, Yuan F and Zhang Y (2023) R‐loops in genome instability and cancer. Cancers (Basel) 15, 4986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112. Abraham KJ, Khosraviani N, Chan JNY, Gorthi A, Samman A, Zhao DY, Wang M, Bokros M, Vidya E, Ostrowski LA et al. (2020) Nucleolar RNA polymerase II drives ribosome biogenesis. Nature 585, 298–302. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113. Lai F, Damle SS, Ling KK and Rigo F (2020) Directed RNase H cleavage of nascent transcripts causes transcription termination. Mol Cell 77, 1032–1043.e4. [DOI] [PubMed] [Google Scholar]
- 114. Lee JS and Mendell JT (2020) Antisense‐mediated transcript knockdown triggers premature transcription termination. Mol Cell 77, 1044–1054.e3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115. Hamperl S and Cimprich KA (2016) Conflict resolution in the genome: how transcription and replication make it work. Cell 167, 1455–1467. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116. Felipe‐Abrio I, Lafuente‐Barquero J, Garcia‐Rubio ML and Aguilera A (2015) RNA polymerase II contributes to preventing transcription‐mediated replication fork stalls. EMBO J 34, 236–250. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117. Lalonde M, Trauner M, Werner M and Hamperl S (2021) Consequences and resolution of transcription‐replication conflicts. Life (Basel) 11, 637. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118. Gan W, Guan Z, Liu J, Gui T, Shen K, Manley JL and Li X (2011) R‐loop‐mediated genomic instability is caused by impairment of replication fork progression. Genes Dev 25, 2041–2056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119. Gomez‐Gonzalez B, Garcia‐Rubio M, Bermejo R, Gaillard H, Shirahige K, Marin A, Foiani M and Aguilera A (2011) Genome‐wide function of THO/TREX in active genes prevents R‐loop‐dependent replication obstacles. EMBO J 30, 3106–3119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120. Wellinger RE, Prado F and Aguilera A (2006) Replication fork progression is impaired by transcription in hyperrecombinant yeast cells lacking a functional THO complex. Mol Cell Biol 26, 3327–3334. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121. Aiello U, Challal D, Wentzinger G, Lengronne A, Appanah R, Pasero P, Palancade B and Libri D (2022) Sen1 is a key regulator of transcription‐driven conflicts. Mol Cell 82, 2952–2966.e6. [DOI] [PubMed] [Google Scholar]
- 122. Alzu A, Bermejo R, Begnis M, Lucca C, Piccini D, Carotenuto W, Saponaro M, Brambati A, Cocito A, Foiani M et al. (2012) Senataxin associates with replication forks to protect fork integrity across RNA‐polymerase‐II‐transcribed genes. Cell 151, 835–846. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123. Schwab RA, Nieminuszczy J, Shah F, Langton J, Lopez Martinez D, Liang CC, Cohn MA, Gibbons RJ, Deans AJ and Niedzwiedz W (2015) The Fanconi anemia pathway maintains genome stability by coordinating replication and transcription. Mol Cell 60, 351–361. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124. Yuce O and West SC (2013) Senataxin, defective in the neurodegenerative disorder ataxia with oculomotor apraxia 2, lies at the interface of transcription and the DNA damage response. Mol Cell Biol 33, 406–417. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125. Lafuente‐Barquero J, Luke‐Glaser S, Graf M, Silva S, Gomez‐Gonzalez B, Lockhart A, Lisby M, Aguilera A and Luke B (2017) The Smc5/6 complex regulates the yeast Mph1 helicase at RNA‐DNA hybrid‐mediated DNA damage. PLoS Genet 13, e1007136. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126. Barnes CO, Calero M, Malik I, Graham BW, Spahr H, Lin G, Cohen AE, Brown IS, Zhang Q, Pullara F et al. (2015) Crystal structure of a transcribing RNA polymerase II complex reveals a complete transcription bubble. Mol Cell 59, 258–269. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127. Bruning JG and Marians KJ (2020) Replisome bypass of transcription complexes and R‐loops. Nucleic Acids Res 48, 10353–10367. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128. Kumar C, Batra S, Griffith JD and Remus D (2021) The interplay of RNA:DNA hybrid structure and G‐quadruplexes determines the outcome of R‐loop‐replisome collisions. Elife 10, e72286. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129. Saponaro M, Kantidakis T, Mitter R, Kelly GP, Heron M, Williams H, Soding J, Stewart A and Svejstrup JQ (2014) RECQL5 controls transcript elongation and suppresses genome instability associated with transcription stress. Cell 157, 1037–1049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130. Urban V, Dobrovolna J, Huhn D, Fryzelkova J, Bartek J and Janscak P (2016) RECQ5 helicase promotes resolution of conflicts between replication and transcription in human cells. J Cell Biol 214, 401–415. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131. Reginato G and Cejka P (2020) The MRE11 complex: a versatile toolkit for the repair of broken DNA. DNA Repair (Amst) 91–92, 102869. [DOI] [PubMed] [Google Scholar]
- 132. Nimonkar AV, Genschel J, Kinoshita E, Polaczek P, Campbell JL, Wyman C, Modrich P and Kowalczykowski SC (2011) BLM‐DNA2‐RPA‐MRN and EXO1‐BLM‐RPA‐MRN constitute two DNA end resection machineries for human DNA break repair. Genes Dev 25, 350–362. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133. Sturzenegger A, Burdova K, Kanagaraj R, Levikova M, Pinto C, Cejka P and Janscak P (2014) DNA2 cooperates with the WRN and BLM RecQ helicases to mediate long‐range DNA end resection in human cells. J Biol Chem 289, 27314–27326. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134. Daley JM, Tomimatsu N, Hooks G, Wang W, Miller AS, Xue X, Nguyen KA, Kaur H, Williamson E, Mukherjee B et al. (2020) Specificity of end resection pathways for double‐strand break regions containing ribonucleotides and base lesions. Nat Commun 11, 3088. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135. Alfano L, Caporaso A, Altieri A, Dell'Aquila M, Landi C, Bini L, Pentimalli F and Giordano A (2019) Depletion of the RNA binding protein HNRNPD impairs homologous recombination by inhibiting DNA‐end resection and inducing R‐loop accumulation. Nucleic Acids Res 47, 4068–4085. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136. Amon JD and Koshland D (2016) RNase H enables efficient repair of R‐loop induced DNA damage. Elife 5, e20533. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137. Gorthi A, Romero JC, Loranc E, Cao L, Lawrence LA, Goodale E, Iniguez AB, Bernard X, Masamsetti VP, Roston S et al. (2018) EWS‐FLI1 increases transcription to cause R‐loops and block BRCA1 repair in Ewing sarcoma. Nature 555, 387–391. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138. Cristini A, Ricci G, Britton S, Salimbeni S, Huang SN, Marinello J, Calsou P, Pommier Y, Favre G, Capranico G et al. (2019) Dual processing of R‐loops and topoisomerase I induces transcription‐dependent DNA Double‐Strand breaks. Cell Rep 28, 3167–3181.e6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139. Crossley MP, Song C, Bocek MJ, Choi JH, Kousouros JN, Sathirachinda A, Lin C, Brickner JR, Bai G, Lans H et al. (2023) R‐loop‐derived cytoplasmic RNA‐DNA hybrids activate an immune response. Nature 613, 187–194. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140. Tan J, Duan M, Yadav T, Phoon L, Wang X, Zhang JM, Zou L and Lan L (2020) An R‐loop‐initiated CSB‐RAD52‐POLD3 pathway suppresses ROS‐induced telomeric DNA breaks. Nucleic Acids Res 48, 1285–1300. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141. Tan J, Wang X, Phoon L, Yang H and Lan L (2020) Resolution of ROS‐induced G‐quadruplexes and R‐loops at transcriptionally active sites is dependent on BLM helicase. FEBS Lett 594, 1359–1367. [DOI] [PubMed] [Google Scholar]
- 142. Saini N, Roberts SA, Sterling JF, Malc EP, Mieczkowski PA and Gordenin DA (2017) APOBEC3B cytidine deaminase targets the non‐transcribed strand of tRNA genes in yeast. DNA Repair (Amst) 53, 4–14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143. Yu K, Chedin F, Hsieh CL, Wilson TE and Lieber MR (2003) R‐loops at immunoglobulin class switch regions in the chromosomes of stimulated B cells. Nat Immunol 4, 442–451. [DOI] [PubMed] [Google Scholar]
- 144. Zheng Y, Lorenzo C and Beal PA (2017) DNA editing in DNA/RNA hybrids by adenosine deaminases that act on RNA. Nucleic Acids Res 45, 3369–3377. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145. Bradley L and Savage KI (2023) ‘From R‐lupus to cancer’: reviewing the role of R‐loops in innate immune responses. DNA Repair (Amst) 131, 103581. [DOI] [PubMed] [Google Scholar]
- 146. Cuartas J and Gangwani L (2022) R‐loop mediated DNA damage and impaired DNA repair in spinal muscular atrophy. Front Cell Neurosci 16, 826608. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147. Perego MGL, Taiana M, Bresolin N, Comi GP and Corti S (2019) R‐loops in motor neuron diseases. Mol Neurobiol 56, 2579–2589. [DOI] [PubMed] [Google Scholar]
- 148. Richard P and Manley JL (2017) R loops and links to human disease. J Mol Biol 429, 3168–3180. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149. Wells JP, White J and Stirling PC (2019) R loops and their composite cancer connections. Trends Cancer 5, 619–631. [DOI] [PubMed] [Google Scholar]
- 150. Wu Y, Song T and Xu Q (2023) R‐LOOPs on short tandem repeat expansion disorders in neurodegenerative diseases. Mol Neurobiol 60, 7185–7195. [DOI] [PubMed] [Google Scholar]
- 151. Lee HG, Imaichi S, Kraeutler E, Aguilar R, Lee YW, Sheridan SD and Lee JT (2023) Site‐specific R‐loops induce CGG repeat contraction and fragile X gene reactivation. Cell 186, 2593–2609.e18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152. Su XA and Freudenreich CH (2017) Cytosine deamination and base excision repair cause R‐loop‐induced CAG repeat fragility and instability in Saccharomyces cerevisiae. Proc Natl Acad Sci USA 114, E8392–E8401. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153. Suraweera A, Lim Y, Woods R, Birrell GW, Nasim T, Becherel OJ and Lavin MF (2009) Functional role for senataxin, defective in ataxia oculomotor apraxia type 2, in transcriptional regulation. Hum Mol Genet 18, 3384–3396. [DOI] [PubMed] [Google Scholar]
- 154. Cristini A, Tellier M, Constantinescu F, Accalai C, Albulescu LO, Heiringhoff R, Bery N, Sordet O, Murphy S and Gromak N (2022) RNase H2, mutated in Aicardi‐Goutieres syndrome, resolves co‐transcriptional R‐loops to prevent DNA breaks and inflammation. Nat Commun 13, 2961. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 155. Park K, Ryoo J, Jeong H, Kim M, Lee S, Hwang SY, Ahn J, Kim D, Moon HC, Baek D et al. (2021) Aicardi‐Goutieres syndrome‐associated gene SAMHD1 preserves genome integrity by preventing R‐loop formation at transcription‐replication conflict regions. PLoS Genet 17, e1009523. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156. Qu X and Chaires JB (2000) Analysis of drug‐DNA binding data. Methods Enzymol 321, 353–369. [DOI] [PubMed] [Google Scholar]
- 157. Takusagawa F, Takusagawa KT, Carlson RG and Weaver RF (1997) Selectivity of F8‐actinomycin D for RNA:DNA hybrids and its anti‐leukemia activity. Bioorg Med Chem 5, 1197–1207. [DOI] [PubMed] [Google Scholar]
- 158. Francis R, West C and Friedman SH (2001) Targeting telomerase via its key RNA/DNA heteroduplex. Bioorg Chem 29, 107–117. [DOI] [PubMed] [Google Scholar]
- 159. West C, Francis R and Friedman SH (2001) Small molecule/nucleic acid affinity chromatography: application for the identification of telomerase inhibitors which target its key RNA/DNA heteroduplex. Bioorg Med Chem Lett 11, 2727–2730. [DOI] [PubMed] [Google Scholar]
- 160. Laspata N, Kaur P, Mersaoui SY, Muoio D, Liu ZS, Bannister MH, Nguyen HD, Curry C, Pascal JM, Poirier GG et al. (2023) PARP1 associates with R‐loops to promote their resolution and genome stability. Nucleic Acids Res 51, 2215–2237. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 161. Patel PS, Abraham KJ, Guturi KKN, Halaby MJ, Khan Z, Palomero L, Ho B, Duan S, St‐Germain J, Algouneh A et al. (2021) RNF168 regulates R‐loop resolution and genomic stability in BRCA1/2‐deficient tumors. J Clin Invest 131, e140105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 162. Shuck SC and Turchi JJ (2010) Targeted inhibition of replication protein a reveals cytotoxic activity, synergy with chemotherapeutic DNA‐damaging agents, and insight into cellular function. Cancer Res 70, 3189–3198. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 163. Edwards DS, Maganti R, Tanksley JP, Luo J, Park JJH, Balkanska‐Sinclair E, Ling J and Floyd SR (2020) BRD4 prevents R‐loop formation and transcription‐replication conflicts by ensuring efficient transcription elongation. Cell Rep 32, 108166. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 164. Lam FC, Kong YW, Huang Q, Vu Han TL, Maffa AD, Kasper EM and Yaffe MB (2020) BRD4 prevents the accumulation of R‐loops and protects against transcription‐replication collision events and DNA damage. Nat Commun 11, 4083. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 165. Ross NT, Lohmann F, Carbonneau S, Fazal A, Weihofen WA, Gleim S, Salcius M, Sigoillot F, Henault M, Carl SH et al. (2020) CPSF3‐dependent pre‐mRNA processing as a druggable node in AML and Ewing's sarcoma. Nat Chem Biol 16, 50–59. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 166. Tajima N, Fukui K, Uesato N, Maruhashi J, Yoshida T, Watanabe Y and Tojo A (2010) JTE‐607, a multiple cytokine production inhibitor, induces apoptosis accompanied by an increase in p21waf1/cip1 in acute myelogenous leukemia cells. Cancer Sci 101, 774–781. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 167. Amato J, Miglietta G, Morigi R, Iaccarino N, Locatelli A, Leoni A, Novellino E, Pagano B, Capranico G and Randazzo A (2020) Monohydrazone based G‐Quadruplex selective ligands induce DNA damage and genome instability in human cancer cells. J Med Chem 63, 3090–3103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 168. Burger AM, Dai F, Schultes CM, Reszka AP, Moore MJ, Double JA and Neidle S (2005) The G‐quadruplex‐interactive molecule BRACO‐19 inhibits tumor growth, consistent with telomere targeting and interference with telomerase function. Cancer Res 65, 1489–1496. [DOI] [PubMed] [Google Scholar]
- 169. Gowan SM, Harrison JR, Patterson L, Valenti M, Read MA, Neidle S and Kelland LR (2002) A G‐quadruplex‐interactive potent small‐molecule inhibitor of telomerase exhibiting in vitro and in vivo antitumor activity. Mol Pharmacol 61, 1154–1162. [DOI] [PubMed] [Google Scholar]
- 170. Jiang S, Awadasseid A, Narva S, Cao S, Tanaka Y, Wu Y, Fu W, Zhao X, Wei C and Zhang W (2020) Anti‐cancer activity of benzoxazinone derivatives via targeting c‐Myc G‐quadruplex structure. Life Sci 258, 118252. [DOI] [PubMed] [Google Scholar]
- 171. Kim MY, Vankayalapati H, Shin‐Ya K, Wierzba K and Hurley LH (2002) Telomestatin, a potent telomerase inhibitor that interacts quite specifically with the human telomeric intramolecular g‐quadruplex. J Am Chem Soc 124, 2098–2099. [DOI] [PubMed] [Google Scholar]
- 172. Ohnmacht SA, Marchetti C, Gunaratnam M, Besser RJ, Haider SM, Di Vita G, Lowe HL, Mellinas‐Gomez M, Diocou S, Robson M et al. (2015) A G‐quadruplex‐binding compound showing anti‐tumour activity in an in vivo model for pancreatic cancer. Sci Rep 5, 11385. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 173. Rodriguez R, Muller S, Yeoman JA, Trentesaux C, Riou JF and Balasubramanian S (2008) A novel small molecule that alters shelterin integrity and triggers a DNA‐damage response at telomeres. J Am Chem Soc 130, 15758–15759. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 174. Xu H, Di Antonio M, McKinney S, Mathew V, Ho B, O'Neil NJ, Santos ND, Silvester J, Wei V, Garcia J et al. (2017) CX‐5461 is a DNA G‐quadruplex stabilizer with selective lethality in BRCA1/2 deficient tumours. Nat Commun 8, 14432. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 175. Zimmer J, Tacconi EMC, Folio C, Badie S, Porru M, Klare K, Tumiati M, Markkanen E, Halder S, Ryan A et al. (2016) Targeting BRCA1 and BRCA2 deficiencies with G‐Quadruplex‐interacting compounds. Mol Cell 61, 449–460. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 176. Safari M, Litman T, Robey RW, Aguilera A, Chakraborty AR, Reinhold WC, Basseville A, Petrukhin L, Scotto L, O'Connor OA et al. (2021) R‐loop‐mediated ssDNA breaks accumulate following short‐term exposure to the HDAC inhibitor Romidepsin. Mol Cancer Res 19, 1361–1374. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 177. Shaw G and Prowse DM (2008) Inhibition of androgen‐independent prostate cancer cell growth is enhanced by combination therapy targeting hedgehog and ErbB signalling. Cancer Cell Int 8, 3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 178. Tan PK, Liu S, Gunic E and Miner JN (2017) Discovery and characterization of verinurad, a potent and specific inhibitor of URAT1 for the treatment of hyperuricemia and gout. Sci Rep 7, 665. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 179. Yang Y, McBride KM, Hensley S, Lu Y, Chedin F and Bedford MT (2014) Arginine methylation facilitates the recruitment of TOP3B to chromatin to prevent R loop accumulation. Mol Cell 53, 484–497. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 180. Neher TM, Shuck SC, Liu JY, Zhang JT and Turchi JJ (2010) Identification of novel small molecule inhibitors of the XPA protein using in silico based screening. ACS Chem Biol 5, 953–965. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 181. Drew AE, Moradei O, Jacques SL, Rioux N, Boriack‐Sjodin AP, Allain C, Scott MP, Jin L, Raimondi A, Handler JL et al. (2017) Identification of a CARM1 inhibitor with potent in vitro and in vivo activity in preclinical models of multiple myeloma. Sci Rep 7, 17993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 182. Wan Y, Zheng X, Chen H, Guo Y, Jiang H, He X, Zhu X and Zheng Y (2015) Splicing function of mitotic regulators links R‐loop‐mediated DNA damage to tumor cell killing. J Cell Biol 209, 235–246. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 183. Marinello J, Bertoncini S, Aloisi I, Cristini A, Malagoli Tagliazucchi G, Forcato M, Sordet O and Capranico G (2016) Dynamic effects of topoisomerase I inhibition on R‐loops and short transcripts at active promoters. PLoS ONE 11, e0147053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 184. Flanagan JM, Funes JM, Henderson S, Wild L, Carey N and Boshoff C (2009) Genomics screen in transformed stem cells reveals RNASEH2A, PPAP2C, and ADARB1 as putative anticancer drug targets. Mol Cancer Ther 8, 249–260. [DOI] [PubMed] [Google Scholar]
- 185. Ramachandran S, Ma TS, Griffin J, Ng N, Foskolou IP, Hwang MS, Victori P, Cheng WC, Buffa FM, Leszczynska KB et al. (2021) Hypoxia‐induced SETX links replication stress with the unfolded protein response. Nat Commun 12, 3686. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 186. Nguyen HD, Leong WY, Li W, Reddy PNG, Sullivan JD, Walter MJ, Zou L and Graubert TA (2018) Spliceosome mutations induce R loop‐associated sensitivity to ATR inhibition in myelodysplastic syndromes. Cancer Res 78, 5363–5374. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 187. Uehara T, Minoshima Y, Sagane K, Sugi NH, Mitsuhashi KO, Yamamoto N, Kamiyama H, Takahashi K, Kotake Y, Uesugi M et al. (2017) Selective degradation of splicing factor CAPERalpha by anticancer sulfonamides. Nat Chem Biol 13, 675–680. [DOI] [PubMed] [Google Scholar]
- 188. Colak D, Zaninovic N, Cohen MS, Rosenwaks Z, Yang WY, Gerhardt J, Disney MD and Jaffrey SR (2014) Promoter‐bound trinucleotide repeat mRNA drives epigenetic silencing in fragile X syndrome. Science 343, 1002–1005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 189. Volkmann S, Jendis J, Frauendorf A and Moelling K (1995) Inhibition of HIV‐1 reverse transcription by triple‐helix forming oligonucleotides with viral RNA. Nucleic Acids Res 23, 1204–1212. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 190. Xu Z, Pilch DS, Srinivasan AR, Olson WK, Geacintov NE and Breslauer KJ (1997) Modulation of nucleic acid structure by ligand binding: induction of a DNA.RNA.DNA hybrid triplex by DAPI intercalation. Bioorg Med Chem 5, 1137–1147. [DOI] [PubMed] [Google Scholar]
- 191. Angelbello AJ, Chen JL, Childs‐Disney JL, Zhang P, Wang ZF and Disney MD (2018) Using genome sequence to enable the Design of Medicines and Chemical Probes. Chem Rev 118, 1599–1663. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 192. De Magis A, Manzo SG, Russo M, Marinello J, Morigi R, Sordet O and Capranico G (2019) DNA damage and genome instability by G‐quadruplex ligands are mediated by R loops in human cancer cells. Proc Natl Acad Sci USA 116, 816–825. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 193. Ruggiero E and Richter SN (2018) G‐quadruplexes and G‐quadruplex ligands: targets and tools in antiviral therapy. Nucleic Acids Res 46, 3270–3283. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 194. Barroso S, Herrera‐Moyano E, Munoz S, Garcia‐Rubio M, Gomez‐Gonzalez B and Aguilera A (2019) The DNA damage response acts as a safeguard against harmful DNA‐RNA hybrids of different origins. EMBO Rep 20, e47250. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 195. Britton S, Dernoncourt E, Delteil C, Froment C, Schiltz O, Salles B, Frit P and Calsou P (2014) DNA damage triggers SAF‐A and RNA biogenesis factors exclusion from chromatin coupled to R‐loops removal. Nucleic Acids Res 42, 9047–9062. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 196. Chang EY, Tsai S, Aristizabal MJ, Wells JP, Coulombe Y, Busatto FF, Chan YA, Kumar A, Dan Zhu Y, Wang AY et al. (2019) MRE11‐RAD50‐NBS1 promotes Fanconi anemia R‐loop suppression at transcription‐replication conflicts. Nat Commun 10, 4265. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 197. Paulsen RD, Soni DV, Wollman R, Hahn AT, Yee MC, Guan A, Hesley JA, Miller SC, Cromwell EF, Solow‐Cordero DE et al. (2009) A genome‐wide siRNA screen reveals diverse cellular processes and pathways that mediate genome stability. Mol Cell 35, 228–239. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 198. Salas‐Armenteros I, Perez‐Calero C, Bayona‐Feliu A, Tumini E, Luna R and Aguilera A (2017) Human THO‐Sin3A interaction reveals new mechanisms to prevent R‐loops that cause genome instability. EMBO J 36, 3532–3547. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 199. Shen W, Sun H, De Hoyos CL, Bailey JK, Liang XH and Crooke ST (2017) Dynamic nucleoplasmic and nucleolar localization of mammalian RNase H1 in response to RNAP I transcriptional R‐loops. Nucleic Acids Res 45, 10672–10692. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 200. Skourti‐Stathaki K, Torlai Triglia E, Warburton M, Voigt P, Bird A and Pombo A (2019) R‐loops enhance Polycomb repression at a subset of developmental regulator genes. Mol Cell 73, 930–945.e4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 201. Tan‐Wong SM, Dhir S and Proudfoot NJ (2019) R‐loops promote antisense transcription across the mammalian genome. Mol Cell 76, 600–616.e6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 202. Bermejo R, Doksani Y, Capra T, Katou YM, Tanaka H, Shirahige K and Foiani M (2007) Top1‐ and Top2‐mediated topological transitions at replication forks ensure fork progression and stability and prevent DNA damage checkpoint activation. Genes Dev 21, 1921–1936. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 203. Drolet M, Phoenix P, Menzel R, Masse E, Liu LF and Crouch RJ (1995) Overexpression of RNase H partially complements the growth defect of an Escherichia coli delta topA mutant: R‐loop formation is a major problem in the absence of DNA topoisomerase I. Proc Natl Acad Sci USA 92, 3526–3530. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 204. Promonet A, Padioleau I, Liu Y, Sanz L, Biernacka A, Schmitz AL, Skrzypczak M, Sarrazin A, Mettling C, Rowicka M et al. (2020) Topoisomerase 1 prevents replication stress at R‐loop‐enriched transcription termination sites. Nat Commun 11, 3940. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 205. Tuduri S, Crabbe L, Conti C, Tourriere H, Holtgreve‐Grez H, Jauch A, Pantesco V, De Vos J, Thomas A, Theillet C et al. (2009) Topoisomerase I suppresses genomic instability by preventing interference between replication and transcription. Nat Cell Biol 11, 1315–1324. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 206. Saha S and Pommier Y (2023) R‐loops, type I topoisomerases and cancer. NAR Cancer 5, zcad013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 207. Cerritelli SM, Frolova EG, Feng C, Grinberg A, Love PE and Crouch RJ (2003) Failure to produce mitochondrial DNA results in embryonic lethality in Rnaseh1 null mice. Mol Cell 11, 807–815. [DOI] [PubMed] [Google Scholar]
- 208. Lockhart A, Pires VB, Bento F, Kellner V, Luke‐Glaser S, Yakoub G, Ulrich HD and Luke B (2019) RNase H1 and H2 are differentially regulated to process RNA‐DNA hybrids. Cell Rep 29, 2890–2900.e5. [DOI] [PubMed] [Google Scholar]
- 209. Zhao H, Zhu M, Limbo O and Russell P (2018) RNase H eliminates R‐loops that disrupt DNA replication but is nonessential for efficient DSB repair. EMBO Rep 19, e45335. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 210. Crow YJ, Leitch A, Hayward BE, Garner A, Parmar R, Griffith E, Ali M, Semple C, Aicardi J, Babul‐Hirji R et al. (2006) Mutations in genes encoding ribonuclease H2 subunits cause Aicardi‐Goutieres syndrome and mimic congenital viral brain infection. Nat Genet 38, 910–916. [DOI] [PubMed] [Google Scholar]
- 211. Kind B, Muster B, Staroske W, Herce HD, Sachse R, Rapp A, Schmidt F, Koss S, Cardoso MC and Lee‐Kirsch MA (2014) Altered spatio‐temporal dynamics of RNase H2 complex assembly at replication and repair sites in Aicardi‐Goutieres syndrome. Hum Mol Genet 23, 5950–5960. [DOI] [PubMed] [Google Scholar]
- 212. Lim YW, Sanz LA, Xu X, Hartono SR and Chedin F (2015) Genome‐wide DNA hypomethylation and RNA:DNA hybrid accumulation in Aicardi‐Goutieres syndrome. Elife 4, e08007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 213. Gutbrod MJ, Roche B, Steinberg JI, Lakhani AA, Chang K, Schorn AJ and Martienssen RA (2022) Dicer promotes genome stability via the bromodomain transcriptional co‐activator BRD4. Nat Commun 13, 1001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 214. Krishnan R, Lapierre M, Gautreau B, Nixon KCJ, El Ghamrasni S, Patel PS, Hao J, Yerlici VT, Guturi KKN, St‐Germain J et al. (2023) RNF8 ubiquitylation of XRN2 facilitates R‐loop resolution and restrains genomic instability in BRCA1 mutant cells. Nucleic Acids Res 51, 10484–10505. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 215. Morales JC, Richard P, Patidar PL, Motea EA, Dang TT, Manley JL and Boothman DA (2016) XRN2 links transcription termination to DNA damage and replication stress. PLoS Genet 12, e1006107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 216. Laverde EE, Polyzos AA, Tsegay PP, Shaver M, Hutcheson JD, Balakrishnan L, McMurray CT and Liu Y (2022) Flap endonuclease 1 Endonucleolytically processes RNA to resolve R‐loops through DNA Base excision repair. Genes (Basel) 14, 98. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 217. Crow YJ, Chase DS, Lowenstein Schmidt J, Szynkiewicz M, Forte GM, Gornall HL, Oojageer A, Anderson B, Pizzino A, Helman G et al. (2015) Characterization of human disease phenotypes associated with mutations in TREX1, RNASEH2A, RNASEH2B, RNASEH2C, SAMHD1, ADAR, and IFIH1. Am J Med Genet A 167A, 296–312. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 218. Hasanova Z, Klapstova V, Porrua O, Stefl R and Sebesta M (2023) Human senataxin is a bona fide R‐loop resolving enzyme and transcription termination factor. Nucleic Acids Res 51, 2818–2837. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 219. Sakasai R, Isono M, Wakasugi M, Hashimoto M, Sunatani Y, Matsui T, Shibata A, Matsunaga T and Iwabuchi K (2017) Aquarius is required for proper CtIP expression and homologous recombination repair. Sci Rep 7, 13808. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 220. Chakraborty P, Huang JTJ and Hiom K (2018) DHX9 helicase promotes R‐loop formation in cells with impaired RNA splicing. Nat Commun 9, 4346. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 221. Mersaoui SY, Yu Z, Coulombe Y, Karam M, Busatto FF, Masson JY and Richard S (2019) Arginine methylation of the DDX5 helicase RGG/RG motif by PRMT5 regulates resolution of RNA:DNA hybrids. EMBO J 38, e100986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 222. Karyka E, Berrueta Ramirez N, Webster CP, Marchi PM, Graves EJ, Godena VK, Marrone L, Bhargava A, Ray S, Ning K et al. (2022) SMN‐deficient cells exhibit increased ribosomal DNA damage. Life Sci Alliance 5, e202101145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 223. Song C, Hotz‐Wagenblatt A, Voit R and Grummt I (2017) SIRT7 and the DEAD‐box helicase DDX21 cooperate to resolve genomic R loops and safeguard genome stability. Genes Dev 31, 1370–1381. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 224. Boleslavska B, Oravetzova A, Shukla K, Nascakova Z, Ibini ON, Hasanova Z, Andrs M, Kanagaraj R, Dobrovolna J and Janscak P (2022) DDX17 helicase promotes resolution of R‐loop‐mediated transcription‐replication conflicts in human cells. Nucleic Acids Res 50, 12274–12290. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 225. Li L, Germain DR, Poon HY, Hildebrandt MR, Monckton EA, McDonald D, Hendzel MJ and Godbout R (2016) DEAD box 1 facilitates removal of RNA and homologous recombination at DNA Double‐Strand breaks. Mol Cell Biol 36, 2794–2810. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 226. Ribeiro de Almeida C, Dhir S, Dhir A, Moghaddam AE, Sattentau Q, Meinhart A and Proudfoot NJ (2018) RNA helicase DDX1 converts RNA G‐Quadruplex structures into R‐loops to promote IgH class switch recombination. Mol Cell 70, 650–662.e8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 227. Mosler T, Conte F, Longo GMC, Mikicic I, Kreim N, Mockel MM, Petrosino G, Flach J, Barau J, Luke B et al. (2021) R‐loop proximity proteomics identifies a role of DDX41 in transcription‐associated genomic instability. Nat Commun 12, 7314. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 228. Chang EY, Novoa CA, Aristizabal MJ, Coulombe Y, Segovia R, Chaturvedi R, Shen Y, Keong C, Tam AS, Jones SJM et al. (2017) RECQ‐like helicases Sgs1 and BLM regulate R‐loop‐associated genome instability. J Cell Biol 216, 3991–4005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 229. Hamadeh Z and Lansdorp P (2020) RECQL5 at the intersection of replication and transcription. Front Cell Dev Biol 8, 324. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 230. Marabitti V, Lillo G, Malacaria E, Palermo V, Pichierri P and Franchitto A (2020) Checkpoint defects elicit a WRNIP1‐mediated response to counteract R‐loop‐associated genomic instability. Cancers (Basel) 12, 389. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 231. Marabitti V, Lillo G, Malacaria E, Palermo V, Sanchez M, Pichierri P and Franchitto A (2019) ATM pathway activation limits R‐loop‐associated genomic instability in Werner syndrome cells. Nucleic Acids Res 47, 3485–3502. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 232. Silva B, Pentz R, Figueira AM, Arora R, Lee YW, Hodson C, Wischnewski H, Deans AJ and Azzalin CM (2019) FANCM limits ALT activity by restricting telomeric replication stress induced by deregulated BLM and R‐loops. Nat Commun 10, 2253. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 233. Ghisays F, Garzia A, Wang H, Canasto‐Chibuque C, Hohl M, Savage SA, Tuschl T and Petrini JHJ (2021) RTEL1 influences the abundance and localization of TERRA RNA. Nat Commun 12, 3016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 234. Wu W, Bhowmick R, Vogel I, Ozer O, Ghisays F, Thakur RS, Sanchez de Leon E, Richter PH, Ren L, Petrini JH et al. (2020) RTEL1 suppresses G‐quadruplex‐associated R‐loops at difficult‐to‐replicate loci in the human genome. Nat Struct Mol Biol 27, 424–437. [DOI] [PubMed] [Google Scholar]
- 235. Muramatsu M, Kinoshita K, Fagarasan S, Yamada S, Shinkai Y and Honjo T (2000) Class switch recombination and hypermutation require activation‐induced cytidine deaminase (AID), a potential RNA editing enzyme. Cell 102, 553–563. [DOI] [PubMed] [Google Scholar]
- 236. Roy D, Yu K and Lieber MR (2008) Mechanism of R‐loop formation at immunoglobulin class switch sequences. Mol Cell Biol 28, 50–60. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 237. Feretzaki M, Pospisilova M, Valador Fernandes R, Lunardi T, Krejci L and Lingner J (2020) RAD51‐dependent recruitment of TERRA lncRNA to telomeres through R‐loops. Nature 587, 303–308. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 238. Nascakova Z, Boleslavska B, Urban V, Oravetzova A, Vlachova E, Janscak P and Dobrovolna J (2021) RAD51 inhibition induces R‐loop formation in early G1 phase of the cell cycle. Int J Mol Sci 22, 3740. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 239. Carey JL and Guo L (2022) Liquid‐liquid phase separation of TDP‐43 and FUS in physiology and pathology of neurodegenerative diseases. Front Mol Biosci 9, 826719. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 240. Giannini M, Bayona‐Feliu A, Sproviero D, Barroso SI, Cereda C and Aguilera A (2020) TDP‐43 mutations link amyotrophic lateral sclerosis with R‐loop homeostasis and R loop‐mediated DNA damage. PLoS Genet 16, e1009260. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 241. Wood M, Quinet A, Lin YL, Davis AA, Pasero P, Ayala YM and Vindigni A (2020) TDP‐43 dysfunction results in R‐loop accumulation and DNA replication defects. J Cell Sci 133, jcs244129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 242. Han SS, Wen KK, Garcia‐Rubio ML, Wold MS, Aguilera A, Niedzwiedz W and Vyas YM (2022) WASp modulates RPA function on single‐stranded DNA in response to replication stress and DNA damage. Nat Commun 13, 3743. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 243. Sarkar K, Han SS, Wen KK, Ochs HD, Dupre L, Seidman MM and Vyas YM (2018) R‐loops cause genomic instability in T helper lymphocytes from patients with Wiskott‐Aldrich syndrome. J Allergy Clin Immunol 142, 219–234. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 244. Bhatia V, Barroso SI, Garcia‐Rubio ML, Tumini E, Herrera‐Moyano E and Aguilera A (2014) BRCA2 prevents R‐loop accumulation and associates with TREX‐2 mRNA export factor PCID2. Nature 511, 362–365. [DOI] [PubMed] [Google Scholar]
- 245. Hatchi E, Skourti‐Stathaki K, Ventz S, Pinello L, Yen A, Kamieniarz‐Gdula K, Dimitrov S, Pathania S, McKinney KM, Eaton ML et al. (2015) BRCA1 recruitment to transcriptional pause sites is required for R‐loop‐driven DNA damage repair. Mol Cell 57, 636–647. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 246. Liang Z, Liang F, Teng Y, Chen X, Liu J, Longerich S, Rao T, Green AM, Collins NB, Xiong Y et al. (2019) Binding of FANCI‐FANCD2 complex to RNA and R‐loops stimulates robust FANCD2 Monoubiquitination. Cell Rep 26, 564–572.e5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 247. Dominguez‐Sanchez MS, Barroso S, Gomez‐Gonzalez B, Luna R and Aguilera A (2011) Genome instability and transcription elongation impairment in human cells depleted of THO/TREX. PLoS Genet 7, e1002386. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 248. Jackson BR, Noerenberg M and Whitehouse A (2014) A novel mechanism inducing genome instability in Kaposi's sarcoma‐associated herpesvirus infected cells. PLoS Pathog 10, e1004098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 249. He X, Yuan J, Gao Z and Wang Y (2023) Promoter R‐loops recruit U2AF1 to modulate its phase separation and RNA splicing. J Am Chem Soc 145, 21646–21660. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 250. Arif W, Mathur B, Saikali MF, Chembazhi UV, Toohill K, Song YJ, Hao Q, Karimi S, Blue SM, Yee BA et al. (2023) Splicing factor SRSF1 deficiency in the liver triggers NASH‐like pathology and cell death. Nat Commun 14, 551. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 251. Komeno Y, Huang YJ, Qiu J, Lin L, Xu Y, Zhou Y, Chen L, Monterroza DD, Li H, DeKelver RC et al. (2015) SRSF2 is essential for hematopoiesis, and its myelodysplastic syndrome‐related mutations dysregulate alternative pre‐mRNA splicing. Mol Cell Biol 35, 3071–3082. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 252. Shiromoto Y, Sakurai M, Minakuchi M, Ariyoshi K and Nishikura K (2021) ADAR1 RNA editing enzyme regulates R‐loop formation and genome stability at telomeres in cancer cells. Nat Commun 12, 1654. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 253. Gong Y and Liu Y (2023) R‐loops at chromosome ends: from formation, regulation, and cellular consequence. Cancers (Basel) 15, 2178. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 254. Lee YW, Arora R, Wischnewski H and Azzalin CM (2018) TRF1 participates in chromosome end protection by averting TRF2‐dependent telomeric R loops. Nat Struct Mol Biol 25, 147–153. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 255. Chu HP, Cifuentes‐Rojas C, Kesner B, Aeby E, Lee HG, Wei C, Oh HJ, Boukhali M, Haas W and Lee JT (2017) TERRA RNA antagonizes ATRX and protects telomeres. Cell 170, 86–101.e16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 256. Herrera‐Moyano E, Mergui X, Garcia‐Rubio ML, Barroso S and Aguilera A (2014) The yeast and human FACT chromatin‐reorganizing complexes solve R‐loop‐mediated transcription‐replication conflicts. Genes Dev 28, 735–748. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 257. Tsai S, Fournier LA, Chang EY, Wells JP, Minaker SW, Zhu YD, Wang AY, Wang Y, Huntsman DG and Stirling PC (2021) ARID1A regulates R‐loop associated DNA replication stress. PLoS Genet 17, e1009238. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 258. Poli J, Gerhold CB, Tosi A, Hustedt N, Seeber A, Sack R, Herzog F, Pasero P, Shimada K, Hopfner KP et al. (2016) Mec1, INO80, and the PAF1 complex cooperate to limit transcription replication conflicts through RNAPII removal during replication stress. Genes Dev 30, 337–354. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 259. Prendergast L, McClurg UL, Hristova R, Berlinguer‐Palmini R, Greener S, Veitch K, Hernandez I, Pasero P, Rico D, Higgins JMG et al. (2020) Resolution of R‐loops by INO80 promotes DNA replication and maintains cancer cell proliferation and viability. Nat Commun 11, 4534. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 260. Bayona‐Feliu A, Barroso S, Munoz S and Aguilera A (2021) The SWI/SNF chromatin remodeling complex helps resolve R‐loop‐mediated transcription‐replication conflicts. Nat Genet 53, 1050–1063. [DOI] [PubMed] [Google Scholar]
- 261. Yang H, Lachtara EM, Ran X, Hopkins J, Patel PS, Zhu X, Xiao Y, Phoon L, Gao B, Zou L et al. (2023) The RNA m5C modification in R‐loops as an off switch of Alt‐NHEJ. Nat Commun 14, 6114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 262. Shih HT, Chen WY, Wang HY, Chao T, Huang HD, Chou CH and Chang ZF (2022) DNMT3b protects centromere integrity by restricting R‐loop‐mediated DNA damage. Cell Death Dis 13, 546. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 263. Zhao DY, Gish G, Braunschweig U, Li Y, Ni Z, Schmitges FW, Zhong G, Liu K, Li W, Moffat J et al. (2016) SMN and symmetric arginine dimethylation of RNA polymerase II C‐terminal domain control termination. Nature 529, 48–53. [DOI] [PubMed] [Google Scholar]
- 264. Silva S, Camino LP and Aguilera A (2018) Human mitochondrial degradosome prevents harmful mitochondrial R loops and mitochondrial genome instability. Proc Natl Acad Sci USA 115, 11024–11029. [DOI] [PMC free article] [PubMed] [Google Scholar]