

Abstract
A major challenge in combating the human immunodeficiency virus (HIV) epidemic is the development of vaccines capable of inducing potent, persistent cellular immunity and broadly reactive neutralizing antibody responses to HIV type 1 (HIV-1). We report here the results of a preclinical trial using the chimpanzee model to investigate a combination vaccine strategy involving sequential priming immunizations with different serotypes of adenovirus (Ad)/HIV-1MNenv/rev recombinants and boosting with an HIV envelope subunit protein, oligomeric HIVSF162 gp140ΔV2. The immunogenicities of replicating and nonreplicating Ad/HIV-1MNenv/rev recombinants were compared. Replicating Ad/HIV recombinants were better at eliciting HIV-specific cellular immune responses and better at priming humoral immunity against HIV than nonreplicating Ad-HIV recombinants carrying the same gene insert. Enhanced cellular immunity was manifested by a greater frequency of HIV envelope-specific gamma interferon-secreting peripheral blood lymphocytes and better priming of T-cell proliferative responses. Enhanced humoral immunity was seen in higher anti-envelope binding and neutralizing antibody titers and better induction of antibody-dependent cellular cytotoxicity. More animals primed with replicating Ad recombinants mounted neutralizing antibodies against heterologous R5 viruses after one or two booster immunizations with the mismatched oligomeric HIV-1SF162 gp140ΔV2 protein. These results support continued development of the replicating Ad-HIV recombinant vaccine approach and suggest that the use of replicating vectors for other vaccines may prove fruitful.
The overall immune responses elicited naturally by human immunodeficiency virus (HIV) infection are not effective at controlling viral replication or disease progression. HIV establishes persistence by immune evasion strategies (13), inherently resisting neutralizing antibodies, repeatedly selecting mutants that escape antibody and T-cell immune responses, and avoiding cytotoxic T-lymphocyte (CTL) killing and impairing CD4 T-cell function by downregulation of major histocompatibility complex class I and CD4 molecules from the surface of infected cells. Since HIV is exquisitely adapted for pathogenesis, an efficacious HIV vaccine will need to induce broader, more potent cellular and humoral immune responses than those elicited by natural infection (7).
Live viral vectors, such as adenovirus (Ad), as vaccine vehicles present one option for inducing more potent immunity. Ads are advantageous because they target epithelial cells of the upper respiratory tract and gut, inducing mucosal immunity critical for preventing HIV infection at genital and/or rectal sites. Ads infect immature dendritic cells (DC), leading to DC maturation and efficient antigen presentation of inserted viral gene products (49, 50). Ads are highly immunogenic and engage both arms of the immune system, eliciting long-lasting cellular and humoral immunity to inserted gene products.
Replication-competent (replicating) Ad-HIV recombinants exploit the potential of Ad vectors for eliciting persistent immune responses. In replicating Ad recombinants, expression of the encoded HIV antigen is incorporated into the Ad replication cycle, so lower immunization doses can achieve longer and higher expression levels of HIV gene product in vivo than replication-defective Ad recombinants. In vivo replication of Ad recombinants stimulates production of proinflammatory cytokines that can augment immune responses. Apoptotic cells arising from Ad replication can provide DC with exogenous antigens for initiation of T-cell responses through cross-presentation (12). Although vaccine vectors may compete with transgenes for induction of immune responses (see below), strong immune responses to Ad antigens may paradoxically enhance immunity to transgene-encoded HIV antigens via CD8-T-cell-mediated autocrine help (39), whereby CD8+ T cells can provide help for other responding CD8+ T cells if present in sufficient numbers (43).
A combination vaccine regimen involving priming with replicating Ad-HIV or simian immunodeficiency virus (SIV) recombinants and boosting with HIV or SIV envelope proteins has elicited strong cellular, humoral, and mucosal immune responses to inserted HIV and SIV gene products in both chimpanzee and macaque models (22, 23, 32, 51). Chimpanzees immunized with this regimen exhibited long-lasting protection against HIV challenges (22, 34), whereas macaques have shown significant protection against a highly pathogenic SIVmac251 challenge (33, 48). Recently, such priming with multigenic Ad-SIV recombinants and boosting with envelope protein subunits induced potent protection against SIVmac251 intrarectal challenge. A total of 39% of immunized macaques remained aviremic after challenge or cleared or controlled plasma viremia to the threshold of detection (33). Protection during the chronic phase of infection was correlated with vaccine-induced cellular immunity and during the acute phase of infection with anti-envelope binding antibodies. The latter antibodies have been recently shown to mediate antibody-dependent cellular cytotoxicity (ADCC), and the activity was significantly correlated with reduced acute-phase viremia (15).
Replication-defective (nonreplicating) Ad recombinants lacking E1 genes required for replication are also being developed (6, 11, 20, 35). In macaques, a nonreplicating Ad5-SIVgag recombinant combined with SIVgag DNA priming very effectively induced high frequencies of SIV-specific T cells and significantly reduced viral burden after a SHIV89.6P challenge (35).
Although these animal model results are encouraging, some obstacles need to be overcome before using Ad vaccines in humans. Preexisting immunity to Ad vectors can impede induction of effective immunity to encoded immunogens. Anti-Ad5 immunity has suppressed nonreplicating Ad/HIV vaccines in mice (4, 38, 46) and rhesus macaques (6). Since Ad-neutralizing antibodies are serotype specific, anti-Ad immunity may be surmounted to a large extent by use of alternative Ad vectors not prevalent in humans (10, 42) or by sequential immunization with Ad vectors of different serotype (22). Use of high-dose nonreplicating Ad vaccines (>1010 PFU) to overcome prior immunity, however, can result in significant toxicity and immunopathology (21, 28, 41). Lower doses of replication-defective Ad vector may avoid toxicity, but at the expense of an adequate immune response as seen in a phase I trial in which only 40% of the volunteers exhibited a positive response to the vaccine (8). Thus, circumventing anti-Ad immunity while using an immunization dose that avoids immunopathology and yet achieves sufficient antigen expression for induction of potent immune responses is critical.
We hypothesized that sequential immunization with low doses of replicating Ad recombinants based in different serotypes would better induce potent persistent cellular immunity to the encoded antigen and more effectively prime strong antibody responses to an envelope protein boost, while circumventing anti-Ad immunity than a nonreplicating Ad recombinant. Therefore, we compared the priming ability and immune responses elicited by low-dose replicating and higher-dose nonreplicating Ad recombinants in chimpanzees, permissive for replication of human Ad, to allow evaluation of both vectors. To evaluate priming of antibody responses, we included boosts with oligomeric HIVSF162 gp140ΔV2, an HIV envelope subunit now being tested in phase I clinical trials. This optimally modified HIV envelope immunogen possesses a trimeric structure, has critical neutralizing epitopes exposed, and elicits antibodies able to neutralize a spectrum of clade B primary isolates (2, 37). In parallel, we assessed induction of humoral and cellular immune responses to the Ad vectors themselves.
MATERIALS AND METHODS
Animals.
Ten chimpanzees were placed into immunization groups well balanced for prior Ad5- and Ad7-neutralizing antibodies (P = 0.82 and 0.65, respectively) (Table 1). The Animal Care and Use Committees of the University of Louisiana at Lafayette and Southwest Foundation for Biomedical Research approved animal use protocols.
TABLE 1.
Demographic information on chimpanzees
| Chimpanzee | Ad vector | Dose | Centera | Age (yr) | Sex | Titerb
|
|
|---|---|---|---|---|---|---|---|
| Ad5 | Ad7 | ||||||
| 271 | Ad5- or Ad7-ΔE3HIVMNenv/rev | 107 | SW | 18 | M | 16 | 8 |
| 363 | Ad5- or Ad7-ΔE3HIVMNenv/rev | 107 | SW | 11 | M | 8 | 4 |
| A163 | Ad5- or Ad7-ΔE3HIVMNenv/rev | 107 | NI | 39 | F | 128 | 16 |
| 182D | Ad5- or Ad7-ΔE3HIVMNenv/rev | 108 | NI | 20 | F | 8 | <4 |
| 386 | Ad5- or Ad7-ΔE3HIVMNenv/rev | 108 | SW | 10 | M | 32 | 16 |
| 376 | Ad5- or Ad7-ΔE1E3HIVMNenv/rev | 108 | SW | 10 | M | 16 | 4 |
| 360 | Ad5- or Ad7-ΔE1E3HIVMNenv/rev | 108 | SW | 11 | F | 128 | <4 |
| 373 | Ad5- or Ad7-ΔE1E3HIVMNenv/rev | 109 | SW | 10 | F | 8 | 8 |
| A136 | Ad5- or Ad7-ΔE1E3HIVMNenv/rev | 109 | NI | 37 | F | 64 | <4 |
| A003 | Ad5- or Ad7-ΔE1E3HIVMNenv/rev | 109 | NI | 15 | F | 64 | 8 |
Chimpanzees were housed at Southwest Foundation for Biomedical Research (SW) and the New Iberia Research Center (NI) as indicated.
Chimpanzees were initially screened for Ad5 and Ad7 neutralizing antibody titers and placed into immunization groups. At the time of study initiation (week 0), the Ad5 and Ad7 titers remained well balanced between animals immunized with replicating and nonreplicating Ad recombinants (P = 0.82 and 0.65 for Ad5 and Ad7, respectively). Titers are expressed as reciprocal serum dilutions.
Vaccines.
Ad5- and Ad7ΔE3HIVMNenv/rev encoding gp160 were obtained from Wyeth-Lederle Vaccines (Pearl River, N.Y.) under a Cooperative Research and Development Agreement. Ad7ΔE1E3HIVMNenv/rev was previously described (29). Ad5ΔE1E3HIVMNenv/rev was constructed by amplifying HIVenv/rev from plasmid Ad5-E3envrev (Wyeth-Lederle Vaccines) and inserting it into the AdEasy pshuttle (17). The pshuttle/HIVenv/rev was digested with PmeI and cotransformed into Escherichia coli BJ5183 cells with pAdEasy-1. Recombinants pAdEasy/HIVenv/rev were selected and confirmed by restriction enzyme analyses. The linearized pAdEasy/HIVenv/rev was transfected into 293 cells. Plaques containing Ad5ΔE1E3HIVenv/rev recombinant were picked and amplified. The Ad5ΔE1E3HIVMNenv/rev recombinant was purified twice on cesium chloride. HIV envelope expression of the four Ad recombinants was confirmed by Western blotting.
Oligomeric HIV-1 SF162 gp140ΔV2 has been described (37).
ELISPOT assay.
Gamma interferon (IFN-γ)-secreting cells in response to HIV-1 Env, Ad5 fiber, and Ad7 fiber peptides were evaluated by ELISPOT by using human IFN-γ enzyme-linked immunospot (ELISPOT) kits (BD Biosciences, San Diego, Calif.). HIV-1MN Env peptides included 80 20-mer peptides with a 10-amino-acid overlap (AIDS Research and Reference Reagent Program, National Institute of Allergy and Infectious Disease, National Institutes of Health). Ad5 fiber and Ad7 fiber peptides consisted of 142 and 78 15-mers, respectively, overlapping by 11 amino acids (Advanced BioScience Laboratories, Inc. [ABL], Kensington, Md.). Peptides were pooled to yield 2 μg of each peptide/ml in the assay. The assay was conducted as specified by the manufacturer with chimp peripheral blood mononuclear cells (PBMC) at 4 × 105 to 0.5 × 105 in 100 μl of RPMI 1640 medium containing 10% human AB serum and 2 mM l-glutamine (R-10) per well and a 30-h culture period with peptide pools. Concanavalin A (Sigma) at 5 μg/ml, R-10, and R-10 containing 0.3% dimethyl sulfoxide were positive and negative controls. Spots were counted by using a KS ELISPOT reader (Zeiss, Inc.). Negative control spots were subtracted.
T-cell proliferation.
Chimp PBMC (105 cells/well) were cultured for 6 days in triplicate in 200 μl of R-10 with HIVIIIB gp120 (ABL) at 1 μg/ml, pulsed overnight with [3H]thymidine (1 μCi/well), harvested, and counted. Concanavalin A (5 μg/ml) and R-10 served as positive and negative controls. Stimulation indices (SI) were calculated by dividing the mean counts per minute with antigen by the mean counts per minute with R-10.
Antibody assays.
Binding antibodies to HIVIIIB gp160 and HIVSF162 gp140ΔV2 were assessed by enzyme-linked immunosorbent assay. Neutralizing antibodies against HIVMN were determined by using MT-2 cells (27). Neutralization of PBMC-grown HIVSF162 and eight other primary HIV clade B isolates, all grown on PBMC except ADA grown on 293T cells, was determined with M7-Luc cells (26).
ADCC was determined by using a rapid, flow cytometry-based assay (V. R. Gómez-Román et al., submitted for publication). Briefly, heat-inactivated chimpanzee sera were incubated 4 h with 50:1 ratios of human PBMC effector cells and CEM-NKr target cells, double stained with (CSFE) 5-(and 6-)carboxyfluorescein diacetate succinimidyl ester and PKH-26, and coated with HIVIIIB gp120. Cells were washed and fixed in 3.7% paraformaldehyde in phosphate-buffered saline (PBS). The percent killing (i.e., the percentage of CSFE-negative cells within the PKH-26 high population) was determined by flow cytometry.
Virus detection by plaque assay.
Nasal swabs, collected in PBS containing 0.1% bovine serum albumin, 0.01% thimerosal, and 750 KalliKrein inhibitor unit of aprotinin, were fivefold serial diluted in Dulbecco modified Eagle medium, applied to confluent A549 cell monolayers, and cultured under 1% agar overlays. Plaques were counted on day 14.
Statistical analysis.
Differences between immunization groups and between doses within groups were tested by using a repeated-measures analysis of variance model including the two factors, their interaction, and time effects in a backward selection procedure. Proliferative responses were also analyzed for a linear trend using repeated measures regression. SI and antibody titers were logarithmically transformed before analysis, and cellular responses to Ad5 and Ad7 fiber peptides were square-root transformed to reduce skewness in the residual distributions, which were consistent with normality over the multiple analyses performed.
RESULTS
Study design and immunization schedule.
Replicating (ΔE3) and nonreplicating (ΔE1E3) recombinant viruses based in Ad5 and Ad7 vectors were constructed containing the identical HIVMNenv/rev gene insert encoding full-length gp160. The gene expression of each of the four Ad recombinants was analyzed by Western blotting after infection of human cells in vitro (Fig. 1). Human 293 cells which contain Ad5 E1 genes can complement the deleted E1 region necessary for replication of the nonreplicating Ad5 vectors. 293-ORF6 cells additionally contain the Ad5 E4 gene, which allows efficient replication of Ad7ΔE1ΔE3 recombinants. Replication of nonreplicating vectors complemented by cell lines in trans is often somewhat less efficient than replication of a self-replicating vector. Nevertheless, the expression level of gp160 from both replicating- and nonreplicating Ad vectors was comparable in the 293 and 293-ORF6 cells. In contrast, env expression of the nonreplicating Ad recombinants in A549 cells was transient, and the level was lower than the replication-competent Ad recombinants, since A549 cells are not permissive for nonreplicating Ad vectors. The A549 cells lack Ad E1 genes, representative of the in vivo situation. Nonreplicating Ad-HIVMNenv/rev recombinants can enter cells and express the inserted env gene under the control of a cytomegalovirus promoter. However, unlike replicating Ad-HIVMNenv/rev recombinants, env gene expression of nonreplicating Ad vectors is not associated with continued rounds of Ad vector replication. Thus, gp160 expression after infection of A549 cells with nonreplicating Ad5- or Ad7-HIVMNenv/rev was transient, and the level was lower than that seen with the replicating Ad5- and Ad7-HIVMNenv/rev recombinants. For this reason, the nonreplicating Ad-HIVMNenv/rev recombinants were administered at doses up to 100-fold higher than the replicating Ad-HIVMNenv/rev recombinants as described below. Doses higher than 109 PFU were not used due to the potential for toxicity. Studies of gene therapy applications in humans showed that intranasal doses of 109 PFU of replication-defective Ad recombinants were safely tolerated, whereas 10-fold-higher doses resulted in local mucosal inflammation (19, 47).
FIG. 1.

Expression of HIVMN envelope by replicating and nonreplicating Ad recombinants. The env gene expression of replicating and nonreplicating Ad-HIVMNenv/rev recombinants was analyzed in 293 or 293-ORF6 cells and in A549 cells by Western blotting with the HIV-1 gp41 monoclonal antibody 2F5. Equivalent PFU of each recombinant were used to infect the indicated cell lines. The nonreplicating Ad7-ΔE1ΔE3-HIV-1MNenv/rev recombinant did not grow in 293 cells but required additional complementation by ORF6 of the Ad5 E4 gene present in 293-ORF6 cells (5) as described previously (29).
Two groups of five chimpanzees each, well matched for prior Ad5 and Ad7 neutralizing antibody titers (Table 1), were intranasally primed with replicating or nonreplicating Ad5-HIVenv/rev recombinants at week 0 and at week 13 with similar recombinants based in the Ad7 vector (Fig. 2). Three chimpanzees were immunized with 107 PFU of the replication-competent Ad recombinants, and three were administered a 100-fold-higher dose (109 PFU) of the replication-defective Ad recombinants. Two chimpanzees in each immunization group received an intermediate bridging dose of 108 PFU of the replicating or nonreplicating recombinants to facilitate statistical analysis. All animals were boosted intramuscularly with oligomeric HIVSF162 gp140ΔV2 at weeks 37 and 49.
FIG. 2.

Immunization schedule. Ad recombinants were administered in 0.5 ml of PBS per nostril. Oligomeric HIV-1SF162 gp140ΔV2 was administered in MF59 adjuvant (1:1 by volume).
Enhanced cellular immunity elicited by replicating Ad recombinants.
Cellular immune responses throughout the immunization regimen were initially assessed on PBMC by ELISPOT assay using overlapping HIV-1MN envelope peptides. Both replicating and nonreplicating Ad recombinants effectively elicited HIV Env peptide-specific IFN-γ-secreting cells (Fig. 3A to D). After the initial Ad5-recombinant immunization, the highest mean spot-forming cells (SFC) for chimpanzees immunized with 107 and 108 PFU of replicating Ad5-HIVenv/rev were 1,085 and 1,702/106 PBMC, respectively. Chimps immunized with 108 and 109 PFU of nonreplicating Ad5-HIVenv/rev exhibited the highest mean responses of 752 and 1,105 SFC. After the second immunization with replicating Ad7-HIVenv/rev recombinants, the highest mean SFC were 903 and 1,455 with doses of 107 and 108 PFU, respectively, and with nonreplicating Ad7-HIVenv/rev recombinants the highest mean values for SFC were 846 and 688, with doses of 108 and 109 PFU, respectively. Over weeks 1 to 13 after Ad5-recombinant administration, a statistically significant increase in SFC due to both the replicating vector (P = 0.0016) and higher dose (P = 0.0083) was seen. A marginally nonsignificant increase in SFC in the replicating group followed Ad7 recombinant administration (weeks 14 to 25, P = 0.051).
FIG. 3.

HIV envelope-specific cellular immune responses. (A to D) Secretion of IFN-γ in response to overlapping HIVMN Env peptides. The results for individual chimpanzees are grouped by immunization dose. Arrows mark Ad5 and Ad7 recombinant immunizations at weeks 0 and 13 and HIVSF162 gp140ΔV2 boosts at weeks 37 and 49. (E) Proliferative responses to HIVIIIB gp120 prior to and 2 weeks after each HIVSF162 gp140ΔV2 boost. Mean SI ± the standard errors of the mean are shown.
HIV-specific cellular immunity persisted over 61 weeks. After the second HIVSF162 gp140ΔV2 boost, SFC peaked at the highest mean levels, with 1,530 and 2,332 SFC exhibited by animals primed with 107 and 108 PFU of replicating Ad-HIVenv/rev, respectively, and 1,005 and 1,395 SFC in animals primed with 108 and 109 PFU of nonreplicating Ad-HIVenv/rev recombinants, respectively. These later responses were perhaps partly attributable to induction of CD4+ helper T cells by the envelope boosters. Over the three time periods after the Ad7-recombinant administration (weeks 29 to 37) and each protein boost (weeks 39 to 49 and weeks 51 to 61), chimpanzees primed with 108 PFU of replicating Ad recombinants exhibited significantly higher numbers of SFC compared to the other three groups (P = 0.0005, 0.020, and 0.0002, respectively), which were not significantly different from each other.
Priming of T-cell proliferative responses was assessed beginning with the first HIVSF162 gp140ΔV2 boost. Strong responses to HIVIIIB gp120 were seen in both immunization groups (Fig. 3E), although more potent proliferative activity was seen in the replicating group. After the first gp140 immunization, four of five chimpanzees in the replicating group exhibited positive SI ranging from 3.0 to 23.7, whereas three of five chimpanzees in the nonreplicating group exhibited SI of 2.7 to 4.1. After the second gp140 booster immunization, significantly higher proliferation in the replicating group was attributable to the replicating vector (P = 0.022) and higher dose (P = 0.0079). Over the three time points evaluated, there was a statistically significant linear trend for better priming of T-cell proliferative responses by the replicating compared to the nonreplicating Ad recombinants (P = 0.010).
Enhanced humoral immunity primed by replicating Ad vectors.
HIV-envelope-specific serum binding antibodies were evaluated with HIVIIIB gp160, heterologous to both the priming and boosting vaccines (Fig. 4). Chimpanzees in the replicating Ad recombinant group developed binding antibodies earlier than those in the nonreplicating group and displayed an ∼10-fold increase in median antibody titer that was maintained until the second protein boost when Ad recombinant priming no longer had an effect. Statistical analysis revealed that after Ad priming and each protein boost, chimpanzees primed with the replicating Ad recombinants developed significantly higher antibody titers to the heterologous HIVIIIB gp160 (P = 0.018; Table 2). In contrast, antibody titers to HIVSF162 gp140ΔV2 were higher in the higher-dose groups (P = 0.0040) but independent of the vector, indicating a direct effect of the homologous protein boost (Table 2).
FIG. 4.

Median binding antibody titers to HIVIIIB gp160 in chimpanzees primed with replicating (•) and nonreplicating (○) Ad recombinants.
TABLE 2.
Anti-envelope binding antibodies elicited by the prime-boost immunization regimens
| Titer group and Immunogen | Dose | Geometric mean titer postimmunizationa
|
||
|---|---|---|---|---|
| After second Ad prime | After first gp140 boost | After second gp140 boost | ||
| Titer against HIVIIIB gp160 | ||||
| Replicating Ad | 107 | 397* | 3,794* | 28,360* |
| Replicating Ad | 108 | 68* | 4,021* | 213,307* |
| Nonreplicating Ad | 108 | 10 | 51 | 10,471 |
| Nonreplicating Ad | 109 | 22 | 161 | 49,570 |
| Titer against HIVSF162 gp140ΔV2 | ||||
| Replicating Ad | 107 | 475 | 3,237 | 13,462 |
| Replicating Ad | 108 | 4,242 | 43,589 | 65,799** |
| Nonreplicating Ad | 108 | 48 | 2,784 | 21,953 |
| Nonreplicating Ad | 109 | 56 | 2,672 | 43,496** |
Serum titers (reciprocal serum dilutions) were obtained 2 weeks past the indicated immunizations. *, Higher titers to heterologous gp160 in chimpanzees primed with replicating Ad recombinants independent of dose (P = 0.018); **, higher titers to homologous gp140ΔV2 Env boost independent of vector (P = 0.0040).
Sera from chimpanzees primed with replicating Ad recombinants were also better able to neutralize T-cell line-adapted HIV-1MN (P = 0.019; Fig. 5A). Antibodies able to neutralize primary HIVSF162 appeared earlier in the replication-competent group (Fig. 5B), but no statistically significant difference in neutralizing titer was seen between immunization groups, indicating the greater effect of the protein boost on the homologous antibody response.
FIG. 5.

Neutralizing antibodies induced by the vaccine regimen. The 50% endpoint titers are shown. TCLA, T-cell line adapted.
Because the HIVSF162 gp140ΔV2 vaccine has elicited broad neutralizing antibodies against several HIV clade B isolates (37), we analyzed eight additional clade B primary isolates (Table 3). After a single boost with HIVSF162 gp140ΔV2, three of five chimpanzees in the replicating group and two of five in the nonreplicating group developed cross-reactive antibodies that neutralized one or more primary isolates. After the second protein boost, all five chimpanzee sera from the replicating group neutralized one to eight of the primary isolates, while three of five sera from the nonreplicating group neutralized one to six isolates. Although there was a tendency for broader primary isolate neutralization by chimpanzees primed with replicating recombinants, it was not statistically significant.
TABLE 3.
The combination Ad-HIVenv/rev priming/gp140ΔV2 boosting regimen elicits broadly reactive antibodies able to neutralize heterologous R5-tropic primary HIV-1 clade B isolates
| Chimpanzee no./Ad dose | Neutralization (% reduction in RLU after first/after second gp140 boost)a
|
|||||||
|---|---|---|---|---|---|---|---|---|
| Bal | JR-FL | Bx08 | 6101 | 692 | 1168 | 1196 | ADA | |
| Replicating Ad | ||||||||
| 271/107 | 45/26 | 21/0 | 23/78 | 0/0 | 12/45 | 10/0 | 55/37 | 58/59 |
| 363/107 | 92/43 | 93/0 | 99/67 | 96/0 | 50/10 | 92/0 | 87/77 | 0/0 |
| A163/107 | 15/88 | 0/81 | 0/96 | 0/90 | 0/85 | 0/82 | 0/97 | 0/80 |
| 386/108 | 35/30 | 4/0 | 42/24 | 0/0 | 21/15 | 0/0 | 0/62 | 32/36 |
| 182D/108 | 35/60 | 0/0 | 70/86 | 0/0 | 23/42 | 0/0 | 83/95 | 0/50 |
| Nonreplicating Ad | ||||||||
| 376/108 | 8/14 | 0/0 | 0/68 | 0/0 | 0/36 | 0/0 | 30/36 | 5/23 |
| 360/108 | 0/9 | 0/1 | 50/72 | 0/19 | 25/12 | 0/0 | 55/33 | 63/77 |
| 373/109 | 45/77 | 38/58 | 53/83 | 27/24 | 51/54 | 39/36 | 84/96 | 79/82 |
| A003/109 | 0/0 | 0/0 | 030 | 0/0 | 0/5 | 0/0 | 31/31 | 26/44 |
| A136/109 | 0/0 | 0/0 | 9/40 | 0/0 | 0/5 | 0/0 | 0/37 | 21/22 |
Sera obtained 2 weeks after each boost with HIVSF162 gp140ΔV2 were evaluated at 1:15 dilutions. Positive results (>50% reduction in relative luciferase units [RLU]) are in boldface.
We recently described a vaccine-elicited functional antibody response mediating ADCC activity (15). The ADCC response, elicited by Ad-SIV recombinant priming and boosted by SIV envelope, was correlated with reductions in acute-phase viremia after a rectal challenge of immunized rhesus macaques with pathogenic SIVmac251. Therefore, we evaluated sera of the similarly immunized chimpanzees for antibody mediating this functional activity. The results showed that, in addition to neutralizing antibodies, ADCC activity mediated by chimpanzee serum antibodies was also induced by the vaccine regimen (Fig. 6). A significant increase in the percent killing of HIV Env-specific target cells due to the replicating Ad recombinants compared to the nonreplicating Ad recombinants was observed over the four time points examined at weeks 15 to 51 (P = 0.022). The geometric mean antibody titers mediating ADCC in both groups were ≥100,000 (data not shown) by week 51 when the second gp140ΔV2 immunization greatly boosted anti-envelope binding antibodies.
FIG. 6.
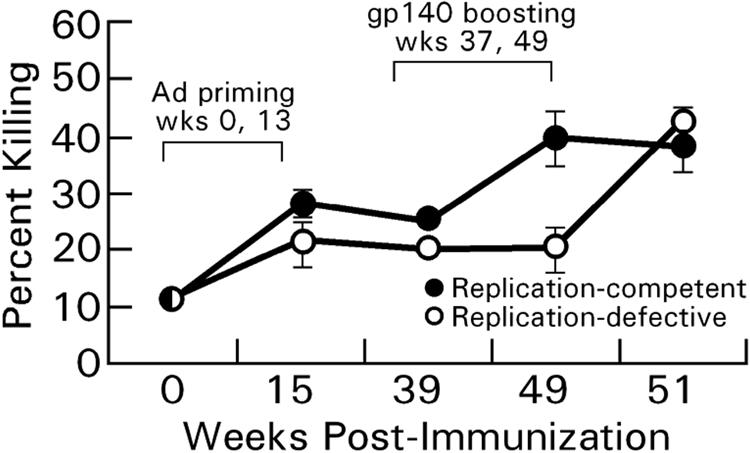
Kinetics of induction of ADCC activity. Mean percent killing of CEM-NKr cells coated with HIVIIIB gp120 by sera (diluted 1:10) of chimpanzees administered replication-competent and replication-defective Ad recombinants is shown. The cutoff for positive killing is 20.4% (negative control plus three standard deviations).
Cellular immune and humoral responses against Ad vectors.
Ad vectors can enhance immunogenicity of transgene-encoded antigens. However, vaccine-induced anti-Ad immunity can also suppress transgene-encoded antigen expression upon readministration of a similar serotype-based Ad vector (44, 45, 47). To circumvent anti-Ad5 immunity elicited by the first Ad5-HIVMNenv/rev immunization, we used a sequential immunization with Ad7-HIVMNenv/rev. We monitored Ad-specific cellular and humoral immune responses elicited by the sequential Ad5 and Ad7 recombinant immunizations to evaluate effects of vector-specific immunity. Potent cellular immune responses to Ad5 fiber peptides were observed in all animals within 2 weeks of immunization with the Ad5 recombinants (Fig. 7A and B). Higher numbers of IFN-γ-secreting cells were associated with higher immunization doses (P = 0.0077). Low to moderate responses to the Ad5 fiber persisted during the 13 weeks following each Ad recombinant immunization and were associated with in vivo replication, since the replicating Ad5 recombinant maintained higher Ad5-specific responses than the nonreplicating Ad5 recombinant (P = 0.0010, weeks 1 to 13; P = 0.046, weeks 14 to 25). In spite of substantial heterogeneity between the Ad5 and Ad7 fiber proteins, modest cross-reactivity to the Ad7 fiber was induced by Ad5 recombinant immunization (weeks 1 to 13; Fig. 7C and D) associated with higher doses (P = 0.0005) but not vector type.
FIG. 7.

Vaccine-induced Ad-specific immunity. Secretion of IFN-γ in response to overlapping Ad5 (A and B) and Ad7 (C and D) fiber peptides. Mean values for chimpanzees receiving the same immunizing dose are shown.
The cellular response to the Ad7 fiber protein after the Ad7 recombinant immunization (weeks 14 to 25; Fig. 7C and D) was lower compared to the anti-Ad5-fiber responses after Ad5 immunization (weeks 1 to 13; Fig. 7A and B) with no difference associated with vector or dose. Higher Ad5 cellular responses may reflect greater immunologic memory to Ad5 resulting from exposure of the chimpanzees to humans, in which Ad5 is more prevalent than Ad7. Persistence of Ad7 immunity over weeks 29 to 37 was associated with higher immunization doses (P = 0.024).
Ad neutralizing antibodies contribute substantially to anti-Ad immunity. To evaluate the effect of serotype-specific neutralizing antibodies on our sequential immunization regimen, we assessed anti-Ad5 and Ad7 neutralizing antibodies over the entire priming period, weeks 0 to 33. In keeping with greater exposure to human Ad5, the chimpanzees had low-level Ad5 neutralizing antibodies at week 0 prior to immunization (Fig. 8A and Table 1). These were boosted by Ad5 recombinant immunization and persisted, perhaps in part due to established immune memory. Ad7 neutralizing antibody titers increased after Ad7 recombinant immunization (Fig. 8B) but subsequently decreased rapidly in the nonreplicating Ad7 recombinant group. Persistent titers in the Ad7 replicating group reflect in vivo replication with resultant prolonged exposure to antigen. Overall, our results reflect the known serotype specificity of Ad neutralizing antibodies.
FIG. 8.
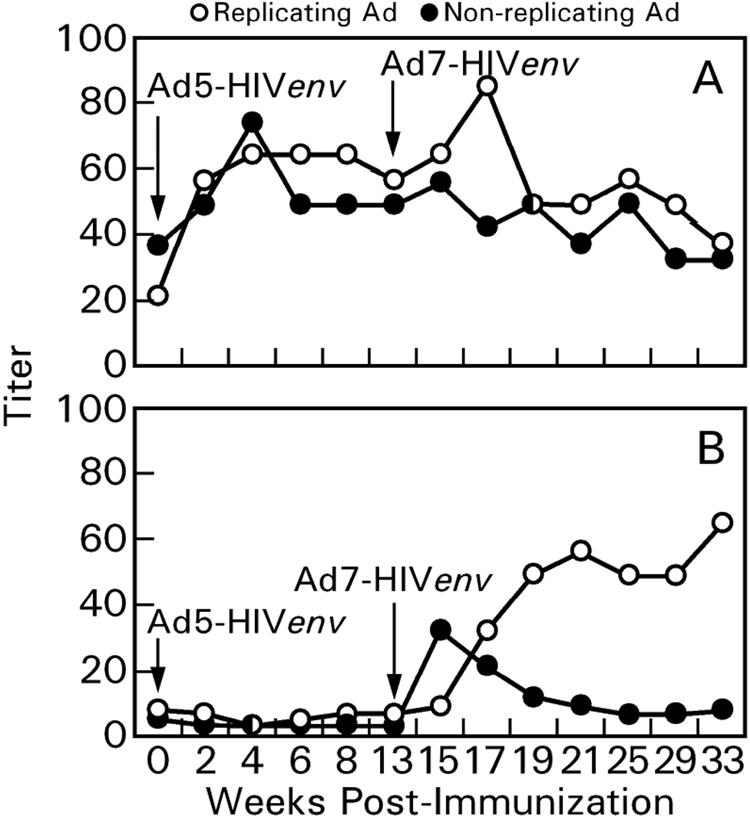
Geometric mean neutralizing antibody titers against Ad5 (A) and Ad7 (B) over the immunization course.
The sequential Ad7 recombinant immunization effectively circumvented anti-Ad5 immunity elicited by the Ad5 recombinants consistent with earlier reports (4, 11, 22). Anti-Ad5 neutralizing antibodies did not neutralize the Ad7-HIVMNenv/rev recombinants. Further, the cross-reactive anti-Ad7 cellular immune response elicited by Ad5 immunization exerted little effect on the replicability of Ad7 recombinants, evaluated by infectious virus shed in nasal secretions. The replicating Ad7 recombinant exhibited similar levels and persistence of replication compared to the Ad5 recombinant (Fig. 9A and B). The results suggest that immunity to a transgene induced by a two-dose sequential immunization regimen with recombinants based in different Ad serotypes is not compromised by the low-level anti-Ad immunity induced. This was verified, as seen in the strong cellular and humoral immune responses to the HIV envelope elicited after Ad7 recombinant immunization (Fig. 3 and 4).
FIG. 9.
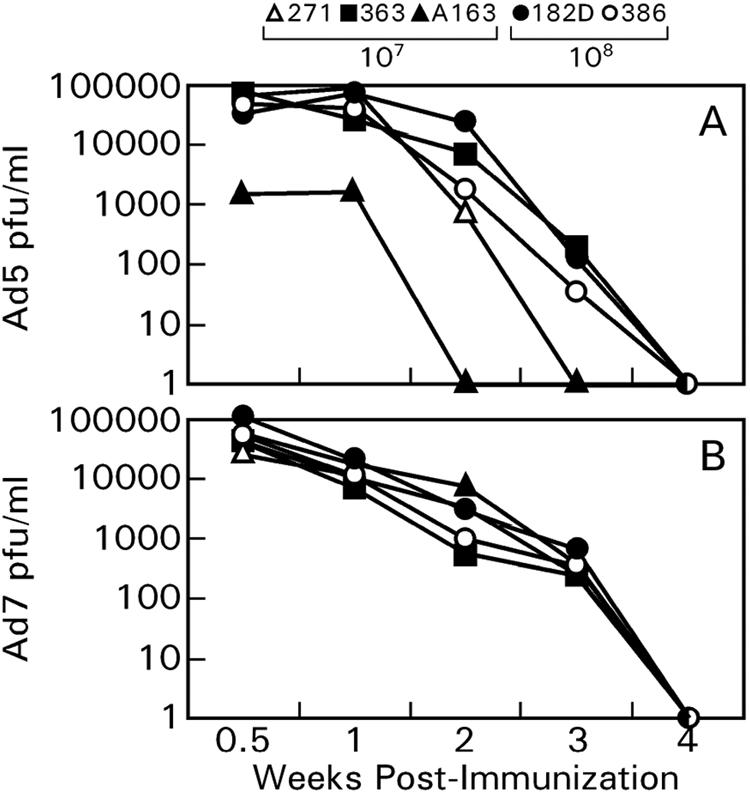
Infectious titers of replication-competent Ad5 (A) and Ad7 (B) recombinants shed into nasal secretions after the Ad5 recombinant immunization at week 0 and the Ad7 recombinant immunization at week 13.
DISCUSSION
Here we have shown that replicating Ad vectors can enhance the immunity of transgene-encoded antigens. At the same or lower doses, replicating Ad recombinants encoding the identical HIVenv/rev elicited significantly stronger HIV-specific cellular and humoral immune responses compared to nonreplicating recombinants. Enhanced cellular immunity was manifested by greater frequency and persistence of HIV Env-specific IFN-γ-secreting cells and by better priming of Env-specific T-cell proliferative responses. Anti-envelope antibodies were more rapidly induced and exhibited higher binding titers against HIVIIIB gp160, an envelope heterologous to the Ad recombinant insert and the protein boost. The replicating recombinants also primed higher neutralizing antibody titers against HIVMN, heterologous to the HIVSF162 gp140ΔV2 boost, and higher ADCC activity.
We speculate that better priming by replicating Ad vectors results from their provision of more HIV antigens for DC presentation through direct infection by Ad recombinants or by Ad-mediated apoptosis or cell lysis. DC acquire antigens from infected dead cells for cross-presentation to CD8 cells. The cross-presentation pathway may be critical for development of effective antiviral immunity (12). Cross-presentation by DC has been reported to be more efficient than direct presentation of HIV antigens (24).
For maximal protection, an HIV vaccine should induce both cellular and humoral immunity. Solely CTL-based AIDS vaccine approaches are limited due to virus escape from immunodominant CTL (3). Antibodies can contribute to vaccine efficacy, as illustrated by passive transfer studies that protected chimpanzees against HIV (9) and macaques against pathogenic SHIV and SIV (1, 16, 25, 30, 31, 40). The HIVSF162 gp140ΔV2 vaccine has elicited broad, potent neutralizing antibodies (2). Here, together with Ad recombinant priming, high-titer neutralizing antibodies were elicited, the highest with the replicating Ad vector. Both Ad vector types coupled with a single HIVSF162 gp140ΔV2 booster immunization primed antibodies that neutralized several primary HIV clade B isolates. Optimal breadth in antibody response may yet be achieved by including additional HIV strains in the recombinant vaccine or booster component.
Functional antibodies mediating ADCC were also significantly elevated in chimpanzees primed with replicating Ad recombinants. ADCC provides a rapid response to invading pathogens, one not dependent on the expansion of memory T cells but only requiring preexisting antibody. Thus, it can potentially enhance vaccine-elicited protection and has been correlated with vaccine protective efficacy in the rhesus macaque model (15).
Consistent with other reports (6, 42), anti-Ad5 cellular immunity was elicited after Ad5 recombinant immunization. However, the modest cross-reactive anti-Ad7 cellular immunity induced was insufficient for suppression of the second immunization with Ad7 recombinant. The sequential immunization strategy successfully induced strong HIV envelope-specific immunity after the week 13 boost with the replicating Ad7 recombinant. Vector-specific immunity may be more of a problem for a nonreplicating vector due to the high doses needed to achieve sufficient expression of inserted genes for induction of an adequate immune response (4, 42). Such high doses of Ad recombinants can result in toxicity and immune pathogenicity that can be compounded in the presence of prior immunity (21, 42).
A lower dose of replicating Ad recombinant more effectively elicits HIV-specific cellular and humoral immunity since in vivo replication of the Ad recombinant elevates the effective dose. Thus, the immunizing dose needs to be carefully established. Although safety is the prime concern with replicating Ad recombinants, over 10 million military recruits were orally administered replication-competent wild-type Ad4 and Ad7 vaccines for 25 years (14). These vaccines were safe and highly effective against outbreaks of acute respiratory disease. Wild-type Ad4 vaccine has also been safely administered intranasally to people at doses of 105 PFU in Ad-seropositive individuals and at doses of 104 PFU in Ad-seronegative people (36). Ads are naturally acquired nasopharyngeally, and they have never been associated with any central nervous system disease. Nevertheless, replication-defective Ad carrying a reporter gene were detected in the olfactory bulb of mice after intranasal administration of 2 × 1010 PFU (20). Therefore, safety studies of Ad-HIV recombinant vaccines for intranasal administration to people will need to proceed cautiously. With 16,000 new HIV infections occurring daily worldwide, there is an urgent need for a vaccine to protect healthy people from HIV infection. A phase I trial of a replicating Ad4-HIV recombinant vaccine is in the planning stages. The safety of oral administration at escalating doses will be evaluated first and then the intranasal route. Overall, the replicating Ad recombinant vaccine approach merits continued development in view of its ability to elicit enhanced immune responses.
Other replicating vectors, alone or in combination, might confer similar advantages to candidate vaccines. Sequential immunization with replicating and nonreplicating vectors from the same or different organisms might combine the initial benefits of a live replicating vaccine with a safer boost, for example, in better inducing mucosal immunity (18), and yet still achieve protective immunity. In addition to vector type, which establishes much of the character and potency of the immunization regimen, a successful AIDS vaccine may require multiple, structurally diverse immunogens to achieve breadth of virus-specific immunity.
Acknowledgments
We thank Irene Kalisz, Sonia Grebogi, Ersell Richardson, and Kris Aldrich for technical assistance and Tong-Chuan He, Howard Hughes Medical Institute, Johns Hopkins Oncology Center, for the AdEasy system. The following were obtained from the AIDS Research and Reference Reagent Program, DAIDS, NIAID: HIV-1MN Env peptides (complete set) and HIV-1 monoclonal antibody 2F5 from Hermann Katinger.
We acknowledge a potential competing interest for three coauthors (E.K., I.S., and S.W.B.), who are employees of Chiron Corp. The company is developing the HIV protein subunit used here as an HIV vaccine.
REFERENCES
- 1.Baba, T. W., V. Liska, R. Hofmann-Lehmann, J. Vlasak, W. Xu, S. Ayehunie, L. A. Cavacini, M. R. Posner, H. Katinger, G. Stiegler, B. J. Bernacky, T. A. Rizvi, R. Schmidt, L. R. Hill, M. E. Keeling, Y. Lu, J. E. Wright, T. C. Chou, and R. M. Ruprecht. 2000. Human neutralizing monoclonal antibodies of the IgG1 subtype protect against mucosal simian-human immunodeficiency virus infection. Nat. Med. 6:200-206. [DOI] [PubMed] [Google Scholar]
- 2.Barnett, S. W., S. Lu, I. Srivastava, S. Cherpelis, A. Gettie, J. Blanchard, S. Wang, I. Mboudjeka, L. Leung, Y. Lian, A. Fong, C. Buckner, A. Ly, S. Hilt, J. Ulmer, C. T. Wild, J. R. Mascola, and L. Stamatatos. 2001. The ability of an oligomeric human immunodeficiency virus type 1 (HIV-1) envelope antigen to elicit neutralizing antibodies against primary HIV-1 isolates is improved following partial deletion of the second hypervariable region. J. Virol. 75:5526-5540. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Barouch, D. H., and N. L. Letvin. 2002. Viral evolution and challenges in the development of HIV vaccines. Vaccine 20(Suppl. 4):A66-A68. [DOI] [PubMed] [Google Scholar]
- 4.Barouch, D. H., M. G. Pau, J. H. Custers, W. Koudstaal, S. Kostense, M. J. Havenga, D. M. Truitt, S. M. Sumida, M. G. Kishko, J. C. Arthur, B. Korioth-Schmitz, M. H. Newberg, D. A. Gorgone, M. A. Lifton, D. L. Panicali, G. J. Nabel, N. L. Letvin, and J. Goudsmit. 2004. Immunogenicity of recombinant adenovirus serotype 35 vaccine in the presence of preexisting anti-Ad5 immunity. J. Immunol. 172:6290-6297. [DOI] [PubMed] [Google Scholar]
- 5.Brough, D. E., A. Lizonova, C. Hsu, V. A. Kulesa, and I. Kovesdi. 1996. A gene transfer vector-cell line system for complete functional complementation of adenovirus early regions E1 and E4. J. Virol. 70:6497-6501. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Casimiro, D. R., L. Chen, T. M. Fu, R. K. Evans, M. J. Caulfield, M. E. Davies, A. Tang, M. Chen, L. Huang, V. Harris, D. C. Freed, K. A. Wilson, S. Dubey, D. M. Zhu, D. Nawrocki, H. Mach, R. Troutman, L. Isopi, D. Williams, W. Hurni, Z. Xu, J. G. Smith, S. Wang, X. Liu, L. Guan, R. Long, W. Trigona, G. J. Heidecker, H. C. Perry, N. Persaud, T. J. Toner, Q. Su, X. Liang, R. Youil, M. Chastain, A. J. Bett, D. B. Volkin, E. A. Emini, and J. W. Shiver. 2003. Comparative immunogenicity in rhesus monkeys of DNA plasmid, recombinant vaccinia virus, and replication-defective adenovirus vectors expressing a human immunodeficiency virus type 1 gag gene. J. Virol. 77:6305-6313. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Desrosiers, R. C. 2004. Prospects for an AIDS vaccine. Nat. Med. 10:221-223. [DOI] [PubMed] [Google Scholar]
- 8.Emini, E. A. 2003. Ongoing development and evaluation of a potential HIV-1 vaccine using a replication-defective adenoviral vector. Presented at the Keystone Symposium on HIV-1 Vaccine Development, Banff, Canada.
- 9.Emini, E. A., W. A. Schleif, J. H. Nunberg, A. J. Conley, Y. Eda, S. Tokiyoshi, S. D. Putney, S. Matsushita, K. E. Cobb, C. M. Jett, J. W. Eichberg, and K. K. Murthy. 1992. Prevention of HIV-1 infection in chimpanzees by gp120 V3 domain-specific monoclonal antibody. Nature 355:728-730. [DOI] [PubMed] [Google Scholar]
- 10.Farina, S. F., G. P. Gao, Z. Q. Xiang, J. J. Rux, R. M. Burnett, M. R. Alvira, J. Marsh, H. C. Ertl, and J. M. Wilson. 2001. Replication-defective vector based on a chimpanzee adenovirus. J. Virol. 75:11603-11613. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Fitzgerald, J. C., G. Gao, A. Reyes-Sandoval, G. N. Pavlakis, Z. Q. Xiang, A. P. Wlazlo, W. Giles-Davis, J. M. Wilson, and H. C. J. Ertl. 2003. A simian replication-defective adenoviral recombinant vaccine to HIV-1 gag. Immunology 170. [DOI] [PubMed]
- 12.Fronteneau, J.-F., M. Larsson, and N. Bhardwaj. 2002. Interactions between dead cells and dendritic cells in the induction of antiviral CTL responses. Curr. Opin. Immunol. 14:471-477. [DOI] [PubMed] [Google Scholar]
- 13.Gandhi, R. T., and B. D. Walker. 2002. Immunologic control of HIV-1. Annu. Rev. Med. 53:149-172. [DOI] [PubMed] [Google Scholar]
- 14.Gaydos, C. A., and J. C. Gaydos. 1995. Adenovirus vaccine in the U.S. military. Mil. Med. 160:300-304. [PubMed] [Google Scholar]
- 15.Gomez-Roman, V. R., L. J. Patterson, D. Venzon, D. Liewehr, K. Aldrich, R. Florese, and M. Robert-Guroff. 2005. Vaccine-elicited antibodies mediate antibody-dependent cellular cytotoxicity correlated with significantly reduced acute viremia in rhesus macaques challenged with SIVmac251. J. Immunol. 174:2185-2189. [DOI] [PubMed] [Google Scholar]
- 16.Haigwood, N. L., A. Watson, W. F. Sutton, J. McClure, A. Lewis, J. Ranchalis, B. Travis, G. Voss, N. L. Letvin, S. L. Hu, V. M. Hirsch, and P. R. Johnson. 1996. Passive immune globulin therapy in the SIV/macaque model: early intervention can alter disease profile. Immunol. Lett. 51:107-114. [DOI] [PubMed] [Google Scholar]
- 17.He, T. C., S. Zhou, L. T. da Costa, J. Yu, K. W. Kinzler, and B. Vogelstein. 1998. A simplified system for generating recombinant adenoviruses. Proc. Natl. Acad. Sci. USA 95:2509-2514. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Herremans, T. M. P. T., J. H. J. Reimerink, A. M. Buisman, T. G. Kimman, and M. P. G. Koopmans. 1999. Induction of mucosal immunity by inactivated poliovirus vaccine is dependent on previous mucosal contact with live virus. J. Immunol. 162:5011-5018. [PubMed] [Google Scholar]
- 19.Knowles, M. R., K. W. Hohneker, Z. Zhou, J. C. Olsen, T. L. Noah, P. C. Hu, M. W. Leigh, J. F. Engelhardt, L. J. Edwards, K. R. Jones, et al. 1995. A controlled study of adenoviral-vector-mediated gene transfer in the nasal epithelium of patients with cystic fibrosis. N. Engl. J. Med. 333:823-831. [DOI] [PubMed] [Google Scholar]
- 20.Lemiale, F., W. Kong, L. M. Akyurek, X. Ling, Y. Huang, B. K. Chakrabarti, M. Eckhaus, and G. J. Nabel. 2003. Enhanced mucosal immunoglobulin A response of intranasal adenoviral vector human immunodeficiency virus vaccine and localization in the central nervous system. J. Virol. 77:10078-10087. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Lozier, J. N., M. E. Metzger, R. E. Donahue, and R. A. Morgan. 1999. Adenovirus-mediated expression of human coagulation factor IX in the rhesus macaque is associated with dose-limiting toxicity. Blood 94:3968-3975. [PubMed] [Google Scholar]
- 22.Lubeck, M. D., R. J. Natuk, M. Myagkikh, N. Kalyan, K. Aldrich, F. Sinangil, S. Alipanah, S. C. S. Murthy, P. K. Chanda, S. Nigida, P. D. Markham, S. Zolla-Pazner, K. Steimer, M. Wade, M. S. Reitz, Jr., L. O. Arthur, S. Mizutani, A. Davis, P. Hung, R. C. Gallo, J. Eichberg, and M. Robert-Guroff. 1997. Long-term protection of chimpanzees against high-dose HIV-1 challenge induced by immunization. Nat. Med. 3:651-658. [DOI] [PubMed] [Google Scholar]
- 23.Malkevitch, N., L. J. Patterson, K. Aldrich, E. Richardson, W. G. Alvord, and M. Robert-Guroff. 2003. A replication competent adenovirus 5 host range mutant-simian immunodeficiency virus (SIV) recombinant priming/subunit protein boosting vaccine regimen induces broad, persistent SIV-specific cellular immunity to dominant and subdominant epitopes in Mamu-A*01 rhesus macaques. J. Immunol. 170:4281-4289. [DOI] [PubMed] [Google Scholar]
- 24.Maranon, C., J. F. Desoutter, G. Hoeffel, W. Cohen, D. Hanau, and A. Hosmalin. 2004. Dendritic cells cross-present HIV antigens from live as well as apoptotic infected CD4+ T lymphocytes. Proc. Natl. Acad. Sci. USA 101:6092-6097. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Mascola, J. R., G. Stiegler, T. C. VanCott, H. Katinger, C. B. Carpenter, C. E. Hanson, H. Beary, D. Hayes, S. S. Frankel, D. L. Birx, and M. G. Lewis. 2000. Protection of macaques against vaginal transmission of a pathogenic HIV-1/SIV chimeric virus by passive infusion of neutralizing antibodies. Nat. Med. 6:207-210. [DOI] [PubMed] [Google Scholar]
- 26.Montefiori, D. C. 2004. Evaluating neutralizing antibodies against HIV, SIV, and SHIV in luciferase reporter gene assays, 12.11.1-12.11.15. In J. E. Coligan, A. M. Kruisbeek, D. H. Margulies, E. M. Shevach, and W. S. Strober (ed.), Current protocols in immunology, in press. John Wiley & Sons, Inc., New York, N.Y. [DOI] [PubMed]
- 27.Montefiori, D. C., W. E. Robinson, Jr., S. S. Schuffman, and W. M. Mitchell. 1988. Evaluation of antiviral drugs and neutralizing antibodies to human immunodeficiency virus by a rapid and sensitive microtiter infection assay. J. Clin. Microbiol. 26:231-235. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Morral, N., W. K. O'Neal, K. Rice, M. M. Leland, P. A. Piedra, E. Aguilar-Cordova, K. D. Carey, A. L. Beaudet, and C. Langston. 2002. Lethal toxicity, severe endothelial injury, and a threshold effect with high doses of an adenoviral vector in baboons. Hum. Gene Ther. 13:143-154. [DOI] [PubMed] [Google Scholar]
- 29.Nan, X., B. Peng, T. W. Hahn, E. Richardson, A. Lizonova, I. Kovesdi, and M. Robert-Guroff. 2003. Development of an Ad7 cosmid system and generation of an Ad7ΔE1ΔE3HIV(MN) env/rev recombinant virus. Gene Ther. 10:326-336. [DOI] [PubMed] [Google Scholar]
- 30.Nishimura, Y., T. Igarashi, N. L. Haigwood, R. Sadjadpour, O. K. Donau, C. Buckler, R. J. Plishka, A. Buckler-White, and M. A. Martin. 2003. Transfer of neutralizing IgG to macaques 6 h but not 24 h after SHIV infection confers sterilizing protection: implications for HIV-1 vaccine development. Proc. Natl. Acad. Sci. USA 100:15131-15136. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Parren, P. W., P. A. Marx, A. J. Hessell, A. Luckay, J. Harouse, C. Cheng-Mayer, J. P. Moore, and D. R. Burton. 2001. Antibody protects macaques against vaginal challenge with a pathogenic R5 simian/human immunodeficiency virus at serum levels giving complete neutralization in vitro. J. Virol. 75:8340-8347. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Patterson, L. J., N. Malkevitch, J. Pinczewski, D. Venzon, Y. Lou, B. Peng, C. Munch, M. Leonard, E. Richardson, K. Aldrich, V. S. Kalyanaraman, G. N. Pavlakis, and M. Robert-Guroff. 2003. Potent, persistent induction, and modulation of cellular immune responses in rhesus macaques primed with Ad5hr-simian immunodeficiency virus (SIV) env/rev, gag, and/or nef vaccines and boosted with SIV gp120. J. Virol. 77:8607-8620. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Patterson, L. J., N. Malkevitch, D. Venzon, J. Pinczewski, V. R. Gomez-Roman, L. Wang, V. S. Kalyanaraman, P. D. Markham, F. A. Robey, and M. Robert-Guroff. 2004. Protection against mucosal simian immunodeficiency virus SIV(mac251) challenge by using replicating adenovirus-SIV multigene vaccine priming and subunit boosting. J. Virol. 78:2212-2221. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Robert-Guroff, M., H. Kaur, L. J. Patterson, M. Leno, A. J. Conley, P. M. McKenna, P. D. Markham, E. Richardson, K. Aldrich, K. Arora, L. Murty, L. Carter, S. Zolla-Pazner, and F. Sinangil. 1998. Vaccine protection against a heterologous, non-syncytium-inducing, primary human immunodeficiency virus. J. Virol. 72:10275-10280. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Shiver, J. W., T. M. Fu, L. Chen, D. R. Casimiro, M. E. Davies, R. K. Evans, Z. Q. Zhang, A. J. Simon, W. L. Trigona, S. A. Dubey, L. Huang, V. A. Harris, R. S. Long, X. Liang, L. Handt, W. A. Schleif, L. Zhu, D. C. Freed, N. V. Persaud, L. Guan, K. S. Punt, A. Tang, M. Chen, K. A. Wilson, K. B. Collins, G. J. Heidecker, V. R. Fernandez, H. C. Perry, J. G. Joyce, K. M. Grimm, J. C. Cook, P. M. Keller, D. S. Kresock, H. Mach, R. D. Troutman, L. A. Isopi, D. M. Williams, Z. Xu, K. E. Bohannon, D. B. Volkin, D. C. Montefiori, A. Miura, G. R. Krivulka, M. A. Lifton, M. J. Kuroda, J. E. Schmitz, N. L. Letvin, M. J. Caulfield, A. J. Bett, R. Youil, D. C. Kaslow, and E. A. Emini. 2002. Replication-incompetent adenoviral vaccine vector elicits effective anti-immunodeficiency-virus immunity. Nature 415:331-335. [DOI] [PubMed] [Google Scholar]
- 36.Smith, T. J., E. L. Buescher, F. H. Top, Jr., W. A. Altemeier, and J. M. McCown. 1970. Experimental respiratory infection with type 4 adenovirus vaccine in volunteers: clinical and immunological responses. J. Infect. Dis. 122:239-248. [DOI] [PubMed] [Google Scholar]
- 37.Srivastava, I. K., L. Stamatatos, E. Kan, M. Vajdy, Y. Lian, S. Hilt, L. Martin, C. Vita, P. Zhu, K. H. Roux, L. Vojtech, C. M. D., J. Donnelly, J. B. Ulmer, and S. W. Barnett. 2003. Purification, characterization, and immunogenicity of a soluble trimeric envelope protein containing a partial deletion of the V2 loop derived from SF162, an R5-tropic human immunodeficiency virus type 1 isolate. J. Virol. 77:11244-11259. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Sumida, S. M., D. M. Truitt, M. G. Kishko, J. C. Arthur, S. S. Jackson, D. A. Gorgone, M. A. Lifton, W. Koudstaal, M. G. Pau, S. Kostense, M. J. Havenga, J. Goudsmit, N. L. Letvin, and D. H. Barouch. 2004. Neutralizing antibodies and CD8+ T lymphocytes both contribute to immunity to adenovirus serotype 5 vaccine vectors. J. Virol. 78:2666-2673. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Truckenmiller, M. E., and C. C. Norbury. 2004. Viral vectors for inducing CD8 T cell responses. Exp. Opin. Biol. Ther. 4:861-868. [DOI] [PubMed] [Google Scholar]
- 40.Van Rompay, K. K., C. J. Berardi, S. Dillard-Telm, R. P. Tarara, D. R. Canfield, C. R. Valverde, D. C. Montefiori, K. S. Cole, R. C. Montelaro, C. J. Miller, and M. L. Marthas. 1998. Passive immunization of newborn rhesus macaques prevents oral simian immunodeficiency virus infection. J. Infect. Dis. 177:1247-1259. [DOI] [PubMed] [Google Scholar]
- 41.Varnavski, A. N., Y. Zhang, M. Schnell, J. Tazelaar, J. P. Louboutin, Q. C. Yu, A. Bagg, G. P. Gao, and J. M. Wilson. 2002. Preexisting immunity to adenovirus in rhesus monkeys fails to prevent vector-induced toxicity. J. Virol. 76:5711-5719. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Vogels, R., D. Zuijdgeest, R. van Rijnsoever, E. Hartkoorn, I. Damen, M. P. de Bethune, S. Kostense, G. Penders, N. Helmus, W. Koudstaal, M. Cecchini, A. Wetterwald, M. Sprangers, A. Lemckert, O. Ophorst, B. Koel, M. van Meerendonk, P. Quax, L. Panitti, J. Grimbergen, A. Bout, J. Goudsmit, and M. Havenga. 2003. Replication-deficient human adenovirus type 35 vectors for gene transfer and vaccination: efficient human cell infection and bypass of preexisting adenovirus immunity. J. Virol. 77:8263-8271. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Wang, B., C. C. Norbury, R. Greenwood, J. R. Bennink, J. W. Yewdell, and J. A. Frelinger. 2001. Multiple paths for activation of naive CD8+ T cells: CD4-independent help. J. Immunol. 167:1283-1289. [DOI] [PubMed] [Google Scholar]
- 44.Yang, Y., Q. Li, H. C. Ertl, and J. M. Wilson. 1995. Cellular and humoral immune responses to viral antigens create barriers to lung-directed gene therapy with recombinant adenoviruses. J. Virol. 69:2004-2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Yang, Y., G. Trinchieri, and J. M. Wilson. 1995. Recombinant IL-12 prevents formation of blocking IgA antibodies to recombinant adenovirus and allows repeated gene therapy to mouse lung. Nat. Med. 1:890-893. [DOI] [PubMed] [Google Scholar]
- 46.Yang, Z. Y., L. S. Wyatt, W. P. Kong, Z. Moodie, B. Moss, and G. J. Nabel. 2003. Overcoming immunity to a viral vaccine by DNA priming before vector boosting. J. Virol. 77:799-803. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Zabner, J., B. W. Ramsey, D. P. Meeker, M. L. Aitken, R. P. Balfour, R. L. Gibson, J. Launspach, R. A. Moscicki, S. M. Richards, T. A. Standaert, S. C. Williams-Warren, A. E. Wadsworth, A. E. Smith, and M. J. Welsh. 1996. Repeat administration of an adenovirus vector encoding cystic fibrosis transmembrane conductance regulator to the nasal epithelium of patients with cystic fibrosis. J. Clin. Investig. 97:1504-1511. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Zhao, J., J. Pinczewski, V. R. Gomez-Roman, D. Venzon, V. S. Kalyanaraman, P. D. Markham, K. Aldrich, M. Moake, D. C. Montefiori, Y. Lou, G. N. Pavlakis, and M. Robert-Guroff. 2003. Improved protection of rhesus macaques against intrarectal simian immunodeficiency virus SIV(mac251) challenge by a replication-competent Ad5hr-SIVenv/rev and Ad5hr-SIVgag recombinant priming/gp120 boosting regimen. J. Virol. 77:8354-8365. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Zhong, L., A. Granelli-Piperno, Y. B. Choi, and R. M. Steinman. 1999. Recombinant adenovirus is an efficient and non-perturbing genetic vector for human dendritic cells. Eur. J. Immunol. 29:964-972. [DOI] [PubMed] [Google Scholar]
- 50.Zhong, L., A. Granelli-Piperno, M. Pope, R. Ignatius, M. G. Lewis, S. S. Frankel, and R. M. Steinman. 2000. Presentation of SIV gag to monkey T cells using dendritic cells transfected with a recombinant adenovirus. Eur. J. Immunol. 30:3281-3290. [DOI] [PubMed] [Google Scholar]
- 51.Zolla-Pazner, S., M. Lubeck, S. Xu, S. Burda, R. J. Natuk, F. Sinangil, K. Steimer, R. C. Gallo, J. W. Eichberg, T. Matthews, and M. Robert-Guroff. 1998. Induction of neutralizing antibodies to T-cell line-adapted and primary human immunodeficiency virus isolates with a prime-boost vaccine regimen in chimpanzees. J. Virol. 72:1052-1059. [DOI] [PMC free article] [PubMed] [Google Scholar]