

Abstract
Nitric oxide (NO) is a signalling and defence molecule of major importance in biology. The flavohaemoglobin Hmp of Escherichia coli is involved in protective responses to NO. Because hmp gene transcription is repressed by the O2-responsive regulator FNR, we investigated whether FNR also senses NO. The [4Fe–4S]2+ cluster of FNR is oxygen labile and controls protein dimerization and site-specific DNA binding. NO reacts anaerobically with the Fe–S cluster of purified FNR, generating spectral changes consistent with formation of a dinitrosyl-iron–cysteine complex. NO-inactivated FNR can be reconstituted, suggesting physiological relevance. FNR binds at an FNR box within the hmp promoter (Phmp). FNR samples inactivated by either O2 or NO bind specifically to Phmp, but with lower affinity. Dose-dependent up-regulation of Phmp in vivo by NO concentrations of pathophysiological relevance is abolished by fnr mutation, and NO also modulates expression from model FNR-regulated promoters. Thus, FNR can respond to not only O2, but also NO, with major implications for global gene regulation in bacteria. We propose an NO-mediated mechanism of hmp regulation by which E.coli responds to NO challenge.
Keywords: flavohaemoglobin/FNR/gene regulation/iron–sulfur clusters/nitric oxide sensing
Introduction
Nitric oxide (NO) is a signalling and defence molecule of major importance, but the biochemical basis of resistance to NO and reactive nitrogen species (RNS) is poorly understood. NO and RNS pose distinct biochemical challenges by reacting under appropriate conditions with several key biomolecules including metalloproteins, thiol groups of proteins and low molecular weight thiols such as glutathione and homocysteine (Hcy) (Poole and Hughes, 2000). These cellular targets may themselves serve as signal transducers, sensing the presence of NO and RNS, resulting in changes in gene expression and consequent synthesis of protective enzymes. Regulation of transcriptional activation in some cases integrates the responses to free radicals of both oxidative and nitrosative stresses. Thus, in Escherichia coli, the soxRS and oxyR regulons, known to respond to superoxide anion and hydrogen peroxide, respectively, are also induced by NO (Nunoshiba et al., 1993; Hausladen et al., 1996; Ding and Demple, 2000). These global antioxidant regulators afford protection against NO or nitrosothiols (Nunoshiba et al., 1993; Hausladen et al., 1996), but the resistance mechanisms are not entirely known, despite their importance in allowing bacteria to combat, for example, macrophage microbicidal activity (De Groote et al., 1997).
The most fully understood mechanism for detoxification of NO involves the bacterial flavohaemoglobin (Hmp). Hmp synthesis is activated primarily by NO or RNS, and the physiological function of Hmp in protecting E.coli from NO is established (Poole et al., 1996; Membrillo-Hernández et al., 1999; Poole and Hughes, 2000). Aerobically, Hmp detoxifies NO by acting as an NO oxygenase (Gardner et al., 1998a; Hausladen et al., 1998) or denitrosylase (Hausladen et al., 2001) and affords inducible protection of aconitase activity (Gardner et al., 1998b) and respiration (Stevanin et al., 2000). Anaerobically, Hmp serves as an NO reductase, generating nitrous oxide (Hausladen et al., 1998; Kim et al., 1999).
Consistent with the role of Hmp in NO detoxification, the flavohaemoglobin-encoding gene of E.coli, hmp, is up-regulated by NO and RNS; this appears not to involve SoxRS (Poole et al., 1996). We have reported (Membrillo-Hernández et al., 1998) a mechanism of hmp gene regulation that involves interaction between S-nitrosothiols and Hcy. Intracellular Hcy is an important co-regulator of several genes involved in methionine biosynthesis, via its effects on MetR, a LysR family DNA-binding protein. One gene activated by MetR with Hcy as cofactor is glyA, which, in E.coli, is adjacent to hmp and divergently transcribed from it. Elevated Hcy levels, achieved either by exogenous Hcy or in certain met mutants, decrease hmp expression. Since Hcy has been shown to be nitrosated by S-nitrosoglutathione (GSNO) (Membrillo-Hernández et al., 1998), such nitrosating agents can deplete Hcy pool sizes, and are postulated to enhance MetR binding at a site proximal to hmp, and up-regulate hmp transcription (Membrillo-Hernández et al., 1998). It is important to recognize that this mechanism does not explain hmp regulation by NO itself. First, NO induces hmp expression anoxically, under which conditions NO will not nitrosate Hcy. Secondly, although the S-nitrosoHcy generated on reaction with GSNO breaks down to release NO, which could itself be the inducer, the reaction of Hcy with sodium nitroprusside (SNP) (which also induces hmp) forms a more stable species from which NO is not released. Thus other mechanisms for hmp regulation by NO, particularly anoxically, must be present.
Anaerobically, the global regulator FNR (Kiley and Beinert, 1999; Green et al., 2001) is also involved in the regulation of hmp since an fnr mutation enhances hmp-lacZ expression (Poole et al., 1996). Inspection of the hmp promoter reveals a DNA-binding site for FNR in a repressing position, but how FNR contributes to NO-mediated regulation has remained obscure. Up-regulation of hmp in Salmonella is achieved by a third mechanism involving the iron uptake regulator Fur (Crawford and Goldberg, 1998).
Here, we report an anaerobic regulatory mechanism of hmp induction directly mediated by NO, involving nitrosylation of the [4Fe–4S]2+ clusters in the FNR protein. NO sensing by FNR has far-reaching implications for a full understanding of its global regulatory functions.
Results
FNR is an NO sensor
The starting point for this work was the observation that the oxygen-responsive global transcription factor FNR represses expression of the NO-detoxifying protein Hmp during anaerobic growth (Poole et al., 1996). FNR senses oxygen via the assembly–disassembly of [4Fe–4S]2+ clusters (Green et al., 1996; Khoroshilova et al., 1997; Kiley and Beinert, 1999). Iron–sulfur clusters of other proteins have been shown to react with NO, and thus the clusters of FNR might also constitute a mechanism to account for NO responsiveness of hmp expression. To address this possibility, the effects of NO on the [4Fe–4S]2+ clusters of FNR were investigated in vitro.
The anoxic, reconstituted FNR protein (holo-FNR) showed the characteristic broad absorbance at ∼420 nm (Khoroshilova et al., 1995; Green et al., 1996; Lazzazera et al., 1996; Jordan et al., 1997) and was devoid of electron parmagnetic resonance (EPR) signals. On addition of the fast NO-releasing compound, proline NONOate, the broad absorbance at 420 nm was replaced progressively by a prominent shoulder at 360 nm and a decrease in absorbance at 420 nm with an isosbestic point near 480 nm (Figure 1A). A plot of ΔA360 versus NO concentration shows that the reaction is complete at a [NO]/[FNR] ratio of <3.0 (Figure 1A inset and B).

Fig. 1. Effect of NO on FNR. (A) Optical spectra of [4Fe–4S]2+ FNR (27 µM, monomer) before and after treatment with aliquots of 4.17 mM proline NONOate to give a final concentration of 140 µM NO (72 µM NONOate). Inset: a plot of ΔA360 versus the ratio of [NO]/[FNR]. When no further change was apparent, a sample was withdrawn for EPR analysis. (B) Optical spectra of the same FNR suspension acquired after sequential additions of proline NONOate. The lowest spectrum is FNR in the absence of NO and the uppermost one is at a [NO]/[FNR] ratio of 5.2. The [NO]/[FNR] ratio increases from 0, 0.2, 0.3, 0.6, 1.2, 2.2, 2.9, 3.7 to 5.2, giving rise to dose-dependent absorbance increases at 360 nm.
Dinitrosyl-iron–cysteine (DNIC) complexes have relatively intense optical absorption bonds in the near UV. Figure 2A displays the absorption spectra of yellow and green DNICs having dimeric and monomeric structures (inset), respectively. The extinction coefficients for the two species are as follows: dimeric clusters ε310 = 9650/M/cm, ε362 = 8529/M/cm and ε750 = 100/M/cm; monomeric form ε362 ∼3580/M/cm, ε603 = 300/M/cm and ε772 = 312/M/cm. The dimeric DNIC is EPR silent, whereas the monomeric form has one unpaired electron, S = 1/2, with an axial g-tensor, g = 2.038, 2.024 and 2.06. The EPR spectrum of a sample taken from the titration of FNR with proline NONOate at a [NO]/[FNR] ratio of ∼5 shows just such an axial EPR spectrum (Figure 3A). This signal is strongly reminiscent of the DNIC of cysteine (Kennedy et al., 1997; Vanin et al., 1998). Integration of the g = 2.038 signal gave only 20% of an electron spin per [4Fe–4S]2+ cluster. A minor species appearing at g = 2.15 may arise from a thiyl radical (data not shown; Kennedy et al., 1997). The EPR spectra measured at low temperature (not shown) revealed a trace amount of a [3Fe–4S]1+ cluster at g = 2.01.

Fig. 2. Optical spectra of DNIC complexes. (A) Model complexes. The monomeric species (445 µM; solid line) displays characteristic absorption maxima at 397 nm (ε ∼3580/M/cm), 603 nm (ε ∼299/M/cm) and 772 nm (ε ∼312/M/cm) (Costanzo et al., 2001). The dimeric species (250 µM, dashed line) displays characteristic absorption maxima at 310 nm (ε ∼9650/M/cm), 362 nm (ε ∼8529/M/cm) and 750 nm (ε ∼100 M/cm). Inset: structures proposed for monomeric and dimeric DNIC species (Costanzo et al., 2001). (B) Comparison between experimental (dashed line) and simulated (black line) spectra of DNICs. The simulated spectrum consists of contributions from 20 and 80% of the monomeric and dimeric DNIC species, respectively, plus apoFNR.

Fig. 3. EPR spectra of [4Fe–4S]2+ FNR. (A) The FNR sample used in Figure 1, after treatment with 140 µM NO (72 µM proline NONOate; 27 µM FNR monomer). (B) Increasing magnitudes of EPR signals elicited by addition of aliquots of NO solution (NO and FNR concentrations were as indicated). The temperature was 77 K. Microwave power and frequency were in (A) 2.007 mW and 9.653 GHz, and in (B) 2.000 mW and 9.669 GHz, respectively. Modulation amplitude, frequency and receiver gain were in (A) 5 gauss (0.5 mT), 100 kHz and 5.02 × 105 and in (B) 10 gauss (1 mT), 100 kHz and 1.00 × 105, respectively.
In a parallel set of experiments, sequential additions of NO solution or air-saturated buffer (∼230 µM O2) to reconstituted FNR also caused absorbance decreases at 420 nm. Some difficulty was experienced with slow protein precipitation causing baseline variations but, during this titration with NO, samples were withdrawn for EPR spectroscopy. The main species detected at 70 K again was typical for a monomeric DNIC (Figure 3B). For example, the EPR spectra of NO-treated FNR are very similar to the EPR spectra of the d7 form of DNIC-aconitase (Kennedy et al., 1997). At 10 K, varying intensities of the signal for the [3Fe–4S]1+ cluster (g = 2.02) were also seen, as previously observed when [4Fe–4S]2+ FNR is briefly exposed to oxygen (Green et al., 1996).
The experimental EPR results (Figure 3) and optical spectra (Figure 1) suggest a mixture of the yellow and green forms of DNIC. We therefore simulated the optical spectrum by combining spectra of the monomeric and dimeric forms in the ratio 20:80. The result is shown in Figure 2B. The match to the experimental spectrum (Figure 2B, dashed line) is good, although the presence of some light-scattering precipitate causes background absorption sloping towards the UV.
An FNR sample exposed to NO solution and lacking the 420 nm peak was flushed with N2 gas for 60 min, then used in a standard procedure previously shown to be effective for reconstitution of the Fe–S cluster (Jordan et al., 1997). Importantly, after overnight incubation, the protein showed the same optical features (420 nm maximum) as an NO-untreated, but reconstituted, sample (data not shown).
These data strongly suggest that the [4Fe–4S]2+ cluster of FNR can react with NO to form a mixture of monomeric and dimeric DNICs, and that the NO-treated protein remains native and can be reconstituted. Thus the conversion of the native [4Fe–4S]2+ cluster to DNICs could provide the molecular basis of a direct NO-responsive switch.
Interaction of FNR with wild-type and mutant Phmp
A putative FNR box, TTGAG----ATCAA, with a strong resemblance to the consensus FNR-binding sequence is centred at position +5.5 in the Phmp region. To determine whether the putative FNR box is actually recognized by FNR, mutations were made in which the central G (underlined above) was mutated to A, and the corresponding C (also underlined), in the second half of the box, was changed to T. The interaction of FNR with its target sequence was then analysed by gel retardation. At holo-FNR concentrations as low as 0.4 and 0.5 µM, 9 and 20% of the wild-type Phmp, respectively, is retarded (Figure 4A). No retardation could be detected when the FNR box was mutated and incubated with holo-FNR (Figure 4D). Interestingly, wild-type Phmp was also retarded (11 and 26%) by dioxygen- and NO-treated FNR lacking the 420 nm peak, but this occurred only with higher protein concentrations (i.e. >>2 µM) (Figure 4B and C). The retardation profile suggests that holo-FNR, as well as O2- and NO-treated FNR, may bind Phmp in a similar, namely dimeric, form, but with different affinities. The protein–DNA dissociation constants (Kd) for the three FNR forms were determined by gel retardation and repressor–operator interaction analyses. Both titration methods showed similar results. O2- and NO-treated FNR had similar affinities for Phmp with apparent dissociation constants of Kd = 2.5–3 µM. The affinity for holo-FNR, Kd = 1 µM, is greater than that of either O2- or NO-treated FNR.

Fig. 4. DNA binding by holo-FNR, O2- and NO-treated FNR proteins. Gel retardation assays were performed by incubating the 32P-labelled Phmp DNA fragment (wild-type) with increasing amounts of (A) holo-FNR, (B) O2-treated FNR and (C) NO-treated FNR protein. The 32P- labelled Phmp DNA fragment (mutated) was incubated with increasing amounts of (D) holo-FNR. Arrows indicate the FNR–target DNA (black) and the FNR–Phmp complex (white). Protein concentrations (monomer) are shown.
DNase I protection by holo-FNR, O2-treated and NO-treated FNR
DNase I footprinting confirmed the different binding affinities of holo-FNR, O2- and NO-treated FNR obtained from titration experiments. A 26 bp region overlapping the FNR box and located between –6 and +20 in Phmp was protected by the addition of 0.2 µM holo-FNR (Figure 5A). Increasing protein concentrations also revealed hypersensitive sites. Once the FNR concentration reached 2–12 µM, an AT-rich hexamer centred at –15.5 appears to be protected from cleavage (Figure 5A and C). The same footprinting pattern was observed with both O2-treated FNR (data not shown) and NO-treated FNR (Figure 5B). Consistent with gel retardation results, 0.2 µM O2- or NO-treated FNR failed to protect the FNR box and protection was seen from 2 µM protein (Figure 5B). Again, the AT-rich hexamer and the hypersensitive sites were revealed by adding further protein. However, in the case of NO-treated FNR (Figure 5B), the hypersensitive +1G and +2A sites were substituted by a –1T site. Also, extensive non-specific DNA protection could be seen upstream of the hypersensitive sites found immediately 5′ to the AT-rich hexamer when higher concentrations of NO-treated FNR were used, indicating that this form of the protein not only has a lower affinity for DNA but also binds with reduced specificity (Figure 5B). Escherichia coli RNA polymerase (RNAP) was used in DNase I footprinting assays to determine whether the σ70-holoenzyme recognizes Phmp. By itself, RNAP bound to a 64 bp DNA region (–44 to +20) containing the –10 and –35 hexamers, as well as the FNR box (Figure 5C).

Fig. 5. DNase I protection by holo-FNR, NO-treated FNR and RNAP bound to Phmp. The 32P-labelled Phmp DNA fragment was digested with DNase I in the presence of various concentrations of (A and C) holo-FNR, (B) NO-treated FNR and (C) RNAP. Template strand patterns shown were obtained with no FNR (lanes -), 0.2 µM (lanes 1), 2 µM (lanes 2), 12 µM (lanes 3) and 20 µM (lanes 4) FNR protein (as monomer), and with 69 nM RNAP (lane 5). Lanes G provide a calibration for the GC base pairs in the Phmp region. Hypersensitive sites are marked with black or white arrowheads. Hollow boxes show the region protected by dimeric FNR binding. Striped boxes show the position of a DNase I-protected AT-rich hexamer. The filled boxes indicate the region bound by RNAP σ70 subunit. The –10 and –35 hexamers, the ribosome-binding site (rbs), the transcription start site (+1) and the FNR box are indicated on the Phmp sequence shown below.
In vivo modulation of FNR activity by NO
Regulation of hmp transcription by NO and RNS appears complex, and at least one regulatory mechanism involving MetR is known (Membrillo-Hernández et al., 1998; Poole and Hughes, 2000). In order to assess the modulation of FNR activity by NO in vivo, we used an E.coli strain carrying a single copy of a Phmp-lacZ fusion (RKP2178). Previously, a bolus addition of a solution of NO gas was used to demonstrate up-regulation of hmp but, in view of the reactivity of NO and the demonstration that E.coli cells consume NO, even anaerobically (Kim et al., 1999), we have used both NO gas solutions (see later, Figure 7) and NO provided by its release from NOC-5 and NOC-7. Figure 6A (traces 1–3) shows the predicted progress of NO release from both NOC-5 and NOC-7 (10 µM of each, final concentration). The release from NOC-5 (half-life of 25 min at pH 7.0, 37°C) is shown as trace 1, and from the donor with a much shorter half life, NOC-7 (5 min at pH 7.0, 37°C) as trace 2. Trace 3 is the sum of the calculated individual NO release profiles. The actual NO concentration in solution (trace 4) was monitored continuously during anaerobic growth with an NO electrode; during the first 10 min of culture, the measured and theoretical NO levels were in good agreement but thereafter NO concentration declined. Nevertheless, significant NO in the range of concentrations previously shown to up-regulate a Phmp-lacZ fusion (Poole et al., 1996) were sustained in the experiment.

Fig. 7. NO modulates gene expression in vivo by reacting with FNR. Anaerobic cultures of E.coli strains carrying (A) the FNR-inducible promoter [FF(–71.5)::lacZ] or (B) the FNR-repressible promoter (FFgalΔ4::lacZ) were treated with different amounts of NO. β-galactosidase activity is the mean of three independent experiments.

Fig. 6. NO derepresses hmp expression in vivo by reacting with FNR. (A) Release of NO by the NOC mixture over 1 h. Traces 1 and 2 are the theoretical NO release patterns at 37°C (in PBS pH 7.4) by NOC-7 (half-life 5 min) and NOC-5 (half-life 25 min), respectively. Trace 3 is the total NO release by the NOC mixture (addition of traces 1 and 2). Trace 4 is the measured NO release by the NOC mixture under the same conditions of NO challenge in anoxic LB-containing cells (OD600 of ∼0.2) at 37°C. Anaerobic cultures of (B) RKP2178 [Φ(hmp-lacZ)1] and (C) RKP2185 (RKP2178 but fnr-271::Tn10) were challenged with different amounts of NOC mixture. The β-galactosidase activity is the mean of three independent repetitions of the experiment; bars show standard deviations.
To determine whether these concentrations of NOC compounds or NO generated were cytotoxic, the anaerobic growth of cultures of strains RKP2178 [Φ(hmp-lacZ)] or RKP2185 [Φ(hmp-lacZ) fnr] (see below) was measured. NOC-5 and NOC-7 were added (5 or 10 µM of each) to exponentially growing cultures that had been pre-grown in sealed tubes to a Klett reading of 20–25. In the first 30 min after NOC addition, growth rates declined slightly but then resumed the rates characteristic of control cultures (data not shown). In an additional test of toxicity, measurements of viable cell numbers were made during a similar anaerobic experiment. Wild-type hmp+ cells were resistant to 500 µM of each NOC compound, whereas hmp mutant cells (strain RKP4600) were sensitive to 50 µM of each NOC compound (results not shown). We have also shown that E.coli can be grown at NOC concentrations as high as 5 mM (M.Binet, J.Laver, H.Cruz-Ramos, R.K.Poole, unpublished data). Thus, NOC compounds in cultures sustain NO in solution over 30 min at concentrations that up-regulate hmp yet are not toxic for growth.
β-galactosidase activities were assayed following the addition of different concentrations of an NOC mixture. Increasing concentrations of NOC compounds elicited an increase in Φ(hmp-lacZ) activity in a wild-type background (Figure 6B) but, in an fnr strain, NOC addition was without effect on the derepressed level of Φ(hmp-lacZ) activity (Figure 6C). These results are consistent with NO reacting with FNR in vivo with consequent derepression of hmp transcription, but additional regulatory mechanisms are suggested by the fact that the derepressed level in the wild-type strain (Figure 6B) is lower than in the fnr mutant (Figure 6C).
In order to assess (i) the modulation of FNR activity by NO in vivo in a simpler system, and (ii) the implications for regulation of other genes in the FNR regulon, we used the pRW50[FF(–71.5)::lacZ] and pRW50(FFgalΔ4::lacZ) fusion plasmids (Williams et al., 1998) providing relatively simple semi-synthetic promoters, at which induction or repression is totally dependent on anaerobic FNR binding. These constructs were used to transform Δlac and Δlac Δfnr strains. The β-galactosidase activities of the anaerobically grown strains were assayed following the addition of different volumes of NO solution. It was anticipated that any disruption by NO of FNR might reduce its inducing activity on FF(–71.5)::lacZ and derepress the FFgalΔ4::lacZ promoter fusion. The titration in vivo of the FNR-inducible promoter [FF(–71.5)] with NO showed a progressive decrease in β-galactosidase activity, reaching only 40% of the NO-untreated level at 25 µM NO (Figure 7A). The titration curve of the FNR-repressible promoter (FFgalΔ4) showed a bell-like shape (Figure 7B). Promoter derepression was observed from 5 to 10 µM NO (1.6- to 2.5-fold increase, respectively) and the highest reporter activity (∼5-fold increase) was reached with 25 µM NO. These data showed that NO is effective as a signal molecule in a relatively narrow concentration range. NO concentrations above 25 µM appear to destabilize lacZ transcripts.
As expected, the FF(–71.5) promoter (strain RKP2761) was inactive in the absence of fnr (93 ± 13 Miller units of β-galactosidase when 10, 20, 30 or 50 µM NO was added; data not shown). Again, as expected, the FFgalΔ4 promoter (strain RKP2762) was active in the fnr background regardless of the presence of NO (210 ± 30 Miller units of β-galactosidase when 10, 20, 30 or 50 µM NO was added). In control experiments, β-galactosidase activities were measured on strains [RKP2672 (Δlac) and RKP2760 (Δlac Δfnr)] carrying the pRW50 plasmid but without the promoter-lacZ fusion; in all cases, the activities were low (<2 Miller units of β-galactosidase) and showed no pattern of variation during anaerobic growth in the presence of NO.
Discussion
Here we present direct evidence of FNR binding to the Phmp in vitro by gel retardation, repressor–operator interaction and footprinting studies. The FNR box centred at position +5.5 was bound specifically by holo-FNR. RNAP occupied a DNA region overlapping the FNR box. Thus, this regulatory element is located appropriately for repression of transcription initiation by promoter occlusion. Point mutations of the central GC pairs within the FNR box were sufficient to abolish protein–DNA interaction, demonstrating the presence of a single FNR-binding site in Phmp. Other FNR-repressed promoters, such as ndh, narX and fnr, contain more than one FNR box (Guest et al., 1996), and complex regulatory mechanisms involving FNR have been described (Meng et al., 1997; Marshall et al., 2001). To our knowledge, Phmp is the first described naturally occurring promoter where repression by the E.coli FNR protein appears relatively simple, i.e. involving a single FNR box in the repressing position.
The DNA-binding activity of FNR was affected by either O2 or NO treatment, resulting in a lower Phmp affinity and specificity. Nevertheless, some residual DNA-binding ability was observed after treatment of the FNR protein with either O2 or NO. Such residual DNA-binding activity has been reported previously for O2-treated FNR (Lazzazera et al., 1996). However, in other work with different DNA targets, treatment of FNR with O2 (Jordan et al., 1997) and either O2 or NO (Wu et al., 2000) abolished DNA binding. Thus, it is likely that the DNA target influences the ability of O2- or NO-treated FNR to bind. The footprinting data for NO-treated FNR indicate similar, but not identical, DNA distortions occurring upon binding of O2-treated FNR. DNase I-hypersensitive sites arise from protein-induced bending that creates kinks in the DNA structure. The DNase I hypersensitivities and the protected AT-rich hexamer found immediately 5′ to the FNR box remained, whereas the +1 FNR half-site appeared less susceptible to the endonuclease when NO-treated FNR was bound. Whether this difference in DNA binding relates to a different FNR structure due to nitrosylation requires further studies since the structure of the FNR protein has not yet been determined.
NO has been demonstrated to react with the Fe–S cluster of several proteins, including NADH dehydrogenase, succinate dehydrogenase, aconitase and SoxR. The present work demonstrates that a [4Fe–4S]2+ cluster can react with NO to form at least two known products, a dimeric and a monomeric DNIC. This is the first time that the dimeric, yellow, EPR-silent, form of a DNIC has been proposed to occur as a protein-bound cluster. The evidence comes from the optical spectrum taken together with the low spin integration of the EPR-active, green monomeric species. The detailed chemistry of the reaction of NO with a susceptible [4Fe–4S]2+ cluster was not clearly established by the study with aconitase (Kennedy et al., 1997). The aconitase studies also failed to obtain more than ∼70% of the EPR-active, S = 1/2 DNIC species. The optical spectra suggest that in this case also a mixture of DNIC forms may be generated.
The results showing up to three NO per Fe–S cluster to complete the overall reaction in FNR suggest a multistep process. The inorganic chemistry of an iron–cysteine–NO mixture shows that the equilibrium
yellow↔green
is driven to the right by increasing the cysteine to iron ratio. In a protein, this ratio will be raised if iron is lost. Hence, iron released from [4Fe–4S]2+ by NO will shift the equilibrium between the two forms to the right. A possible reaction scheme for FNR, therefore, is that NO converts the [4Fe–4S]2+ cluster to the [2Fe–2S]2+ form, as is the case with O2, followed by further reaction to generate a dimeric then a monomeric DNIC. A reaction between a [2Fe–2S] cluster and NO was proposed in the case of the reaction with SoxR (Ding and Demple, 2000). In either scheme, the [4Fe–4S]2+ cluster would undergo a ligational and, hence, substantial conformational change.
We previously demonstrated that NO acts as an effector in FNR regulation of Azotobacter vinelandii gene expression (Cruz-Ramos et al., 2000; Wu et al., 2000). FnrP from Paracoccus denitrificans also appears sensitive to SNP, as judged by effects in vivo (Hutchings et al., 2002). However, NO sensing by E.coli FNR has special physiological significance since FNR regulates expression of Hmp, which protects against NO and nitrosative stress. The biological significance of this protein is emphasized by our recent finding (Stevanin et al., 2002) that Hmp protects Salmonella from NO-related killing by human macrophages, and similar findings have been obtained with E.coli (T.M.Stevanin, R.C.Read and R.K.Poole, unpublished data). That the sensing of micromolar NO concentrations is physiologically relevant is suggested by the results of Krieglstein et al. (2001) who report the production of NO metabolites (NOx) by polymorphonuclear leukocytes to be in the range 12–37 µM. Similarly, total NO metabolites (measured as nitrite) produced by RAW 264.7 macrophages co-cultured with arginase-deficient Helicobacter pylori are in the approximate range 15–22 µM (Gobert et al., 2001). Here, both adding a solution of NO gas and by maintenance of NO in cultures using NOC compounds, we demonstrate that FNR can sense pathophysiological levels of NO (5–10 µM) in vivo. If intracellular NO concentrations are similar, the FNR concentration in cells (estimated to be 1 µM; Unden and Duchêne, 1987) is appropriate for NO sensing. The present work strongly suggests an NO-mediated mechanism of anaerobic up-regulation of the E.coli hmp gene by nitrosylation of the FNR [4Fe–4S]2+ clusters (Figure 8).
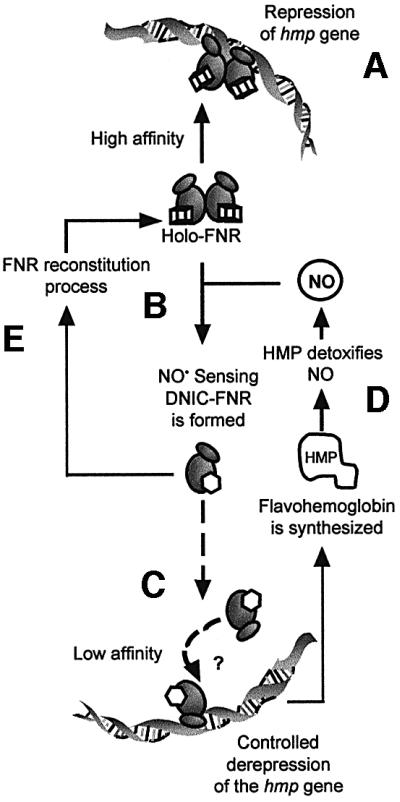
Fig. 8. NO-mediated mechanism of anaerobic up-regulation of the E.coli hmp gene. FNR forms are represented by ovals. Cubes and hexagons represent the [4Fe–4S]2+ and nitrosylated clusters, respectively. DNA (Phmp), NO and Hmp molecules are also shown. (A) Holo-FNR–DNA binding and repression of hmp under anaerobic conditions. (B) FNR senses NO by reaction with [4Fe–4S]2+ clusters to generate DNIC–FNR. This post-transcriptional modification is likely to promote monomerization of FNR. (C) NO-treated FNR (and apo-FNR) can eventually bind Phmp with a lower affinity to avoid sudden and complete derepression of the hmp gene. (D) The NO signal transduction through FNR allows derepression of hmp transcription. Synthesized Hmp protein detoxifies NO to produce N2O or NO3– in the absence or presence of O2, respectively (Poole and Hughes, 2000). (E) When the NO concentration is reduced, putative Fe–S cluster-repairing mechanisms can reconstitute the NO sensor, FNR.
The results suggest that other FNR-regulated genes should be affected by NO, but we are unaware of any cases described thus far in which gene regulation in bacteria is so regulated. The products of other FNR-repressed genes, such as acnA (aconitase), sucAB (α-ketoglutarate dehydrogenase), sucCD (succinyl CoA synthetase), fumA (fumarase A) and icd (isocitrate dehydrogenase) might, for example, represent a response to NO challenge by maintaining flux through the tricarboxylic acid cycle. Other E.coli genes directly repressed by FNR include fpg/mutM (Lee et al., 1998), sodA (Hassan and Sun, 1992) and sodC (Gort et al., 1999). These genes respond to oxidative stress and the products have roles in defence responses. In addition, superoxide dismutases SodA (MnSOD) and SodC (CuZnSOD) in the cytoplasm and periplasm, respectively, might contribute to NO stress tolerance by removing superoxide that can react with NO to form the toxic species, peroxynitrite. The fpg/mutM gene product, oxidized guanine glycosylase, repairs oxidative DNA damage (Chung et al., 1991), removing 8-hydroxyguanine and formamidopyrimidine residues in DNA, although deamination of DNA normally is attributed to NO (Wink et al., 1991).
Evidence that NO signal transduction is achieved by other Fe–S proteins is also presented in eukaryotic systems by the iron regulatory protein IRP1, which has a dual function. As an aconitase, IRP1 possesses a [4Fe–4S] cluster and catalyses the isomerization of citrate and isocitrate. As a cluster-depleted protein, it binds specific RNA structures called iron-responsive elements (IREs) found in several mRNAs, like those encoding the ferritin subunits and transferrin receptor (Hentze and Kühn, 1996; Richardson and Ponka, 1997). The enzymatic/regulatory IRP1 activity is sensitive to endogenous chemical signals provided by nitrosative stress (Pantopoulos and Hentze, 1995; Bouton et al., 1997; Oliveira et al., 1999; Wardrop et al., 2000). Exposure of IRP1 to NO or peroxynitrite flux elicited an increase in IRE-binding activity to the detriment of aconitase activity. SNAP-generated NO increased IRP1 regulatory activity, while SNP-generated NO+ decreased this activity as a function of concentration (Wardrop et al., 2000). At present, the mechanism(s) of these effects on IRP1 remains unclear.
In summary, we have shown that direct modification of the FNR [4Fe–4S]2+ clusters to form protein-bound dinitrosyl-iron–thiol complexes serve as a functional switch in NO-responsive control of gene expression. Specifically, NO inactivation of FNR results in up-regulation of synthesis of Hmp, a protein with key roles in NO resistance. This study reveals FNR to be a major NO-responsive regulator in bacteria, although other NO sensors surely remain to be discovered.
Materials and methods
Bacterial strains and medium
Escherichia coli VJS676 [Δ(argF-lacZ)U169; wild-type] and derivatives RKP2178 [Φ(hmp-lacZ)1], RKP2185 (RKP2178 fnr-271::Tn10) and RKP4600 (same as RKP4545, i.e. hmp::Tn5) have been described previously (Poole et al., 1996; Membrillo-Hernández et al., 1999). The FNR-inducible promoter, FF(–71.5)::lacZ, and FNR-repressible promoter, FFgalΔ4::lacZ, were cloned into pRW50 (Williams et al., 1998). Cells were grown in rich medium (LB), initial pH 7.0. Kanamycin, tetracycline and rifampicin were used at final concentrations of 100, 35 and 50 µg/ml, respectively.
Treatment with NO and nitric oxide releasers
To release NO slowly in cultures, an anoxic equimolar mixture of NOC (Calbiochem) compounds was used: NOC-5 (3-[2-hydroxy-1-(1-methylethyl)-2-nitrosohydrazino]-1-propanamine) and NOC-7 (3-[2-hydroxy-1-methyl-2-nitrosohydrazino]-N-methyl-1-propanamine) freshly prepared within an anaerobic glove box (Mk3, Don Whitley Scientific Ltd) in buffer (25 mM Tris pH 10.0, 100 mM NaCl) and kept at 4°C. The amount of NO released by the NOC mixture in an LB culture (OD600 of ∼0.2) at 37°C was measured at least three times by using simultaneously (Mills et al., 2001) O2 (Rank Bros.) and NO electrodes (World Precision Instruments ISO-NO Mark II) over a 1 h period. The NOC mixture was added by injection into the stirred perspex chamber on depletion of all measurable oxygen by the cells (∼5 min). Calibration of the amount of NO was carried out in the same conditions by injection of various volumes of a freshly prepared anoxic solution of NO (∼1.9 mM in H2O at room temperature; Poole et al., 1996). Theoretical NO release (two NO per NOC) was calculated using half-lives of NOCs in phosphate-buffered saline (PBS; pH 7.4) at 37°C for NOC-7 and NOC–5 of 5 and 25 min, respectively (Calbiochem). Anaerobic cultures of strains RKP2178 and RKP2185 were grown to an OD600 of ∼0.2 as described previously (Poole et al., 1996) in screw-cap glass tubes filled with medium to the brim at 37°C in a water bath. The NOC mixture was added within the glove box to final concentrations in the range of 1–10 µM, and the anaerobic incubation continued for 1 h. Since NOCs are polyamines that can have regulatory effects on gene expression, the control culture (zero NO) was treated with 10 µM of a NOC mixture that was left at room temperature until no signal from the NO electrode was detectable. After treatment with NOC, rifampicin (50 µg/ml) was added to the cultures to stop gene transcription caused by exposure to oxygen. Bacteria were pelleted, washed with PBS and stored at –20°C before β-galactosidase assay.
For studies of strains carrying the pRW50[FF(–71.5)::lacZ] and pRW50(FFgalΔ4::lacZ) fusion plasmids, anaerobic cultures were diluted 20-fold into 10 ml of anoxic LB medium containing tetracycline (35 µg/ml) in 15 ml centrifuge tubes. After 4 h incubation in the anaerobic box (∼25°C), NO was added to final concentrations up to 50 µM, the incubation was continued for 2 h and chloramphenicol (34 µg/ml) was added to the cultures to stop protein synthesis and avoid oxygen interference. Bacteria were pelleted, washed with 5 ml of PBS (137 mM NaCl; 2.7 mM KCl; 10 mM Na2HPO4 and 1.4 mM KH2PO4 pH 7.4) and stored at –20°C before β-galactosidase assay.
Toxicity of NO solutions
To determine the toxicity of NO, cultures of strains VJS676 (wild-type) and RKP4600 (hmp) were grown aerobically at 37°C to an OD600 of ∼0.2. The cultures were divided, one-half being left for 30 min in the glove box at 37°C with slow shaking (50 r.p.m.) in order to obtain anaerobic conditions, and the other half left growing aerobically at 37°C with shaking (100 r.p.m.). Then, cultures were diluted to the same OD600 (∼0.2) with either aerobic or anaerobic LB medium. Cultures were challenged in triplicate with 3, 5, 10, 20, 50, 100 and 500 µM NOC mixture. After 1 h further incubation, the cultures were serially diluted in medium and plated in triplicate on nutrient agar plates containing antibiotic, when appropriate. Plates were incubated overnight at 30°C and colonies were counted. Results were expressed as a percentage of the viable counts in cultures not exposed to the NOC mixture. Anaerobic growth curves were obtained by inoculating 80 µl of an overnight culture into sealed tubes filled to the brim with 8 ml of LB medium. Cultures were grown at 37°C on a slowly shaking platform for 90 min before NOC-5 and NOC-7 were added from stock solutions at high pH. A control tube lacked NOC compounds.
Purification, reconstitution and O2 treatment of FNR
FNR protein was overproduced, purified and reconstituted as described previously (Jordan et al., 1997). FNR samples in TCB (50 mM Tris–HCl, 150 mM NaCl, 2.5 mM CaCl2) were transferred from the anaerobic box in sealed cuvettes to a spectrophotometer (Beckman DU®650). Air-saturated TCB (∼230 µM O2 at 23°C) was added to the sample by injection in the glove box.
Preparation of the NO-treated protein
Two methods were used. In the first, reconstituted FNR protein was treated in the glove box with aliquots of a fresh anoxic solution of NO. Progress of the reaction was monitored as above. In the second, reactions with NO were performed using the fast release agent, proline NONOate (Alexis Chemicals), and the FNR protein was resuspended in nitrate-containing working buffer (25 mM HEPES, 2.5 mM CaCl2, 100 mM NaCl, 100 mM NaNO3 pH 7.5). Proline NONOate was stored at ∼1°C in buffer (same as the NOC mixture). Release of NO occurs with a half-time of ∼13 s when titrated in working buffer at ∼25°C. The amount of NO released per NONOate was measured by capturing the NO with the haem protein Fe(II)-Mb, and estimated to be 1.9 ± 0.1. FNR was held in a sealed cuvette within the glove box and titrated by injection with aliquots of a 4.17 mM stock solution of NONOate. After the addition of each aliquot, the cuvette was placed immediately into a spectrophotometer (Hitachi U3200) scanning at 120 nm/min. At the end point of the titration, an aliquot of NO-treated FNR was withdrawn and put into an EPR tube within the glove box.
Preparation of DNIC complexes
To determine the optical characteristics of DNIC complexes, an anaerobic solution of 500 µM (NH4)2Fe(SO4)2, 1 mM l-cysteine, 25 mM HEPES, 100 mM NaCl pH 7.5 was injected with an aliquot of proline NONOate to give a final concentration of 692 µM (equivalent to 1.38 mM NO). The resulting solution yielded the yellow or dimeric species. Conversion of the dimeric species into the green or monomeric species was achieved by increasing the l-cysteine concentration to 23.5 mM. Optical spectra were obtained using a Hitachi U3200 spectrophotometer and sealed anaerobic cuvettes.
EPR spectroscopy
EPR spectra were recorded on an X-band Bruker EMX spectrometer (9.64 GHz) equipped with an ESR-900 helium-flow cryostat and a TE-102 microwave cavity (Oxford Instruments). Field intensity was monitored using a microwave counter model 2440 (Marconi Instruments). Spin intensities of paramagnetic samples were estimated from integrations of EPR absorption spectra using 1 mM Cu(II)/10 mM EDTA as the standard solution.
Construction of mutant Phmp and labelling
In order to change the FNR box in Phmp, primers RP86 (5′-TAT AAGATGCATTTAAGATACATTAATTAAGATGCAAAAAAAGG-3′, incorporating the underlined mutations, G→A and C→T, and the italicized VspI restriction site) and RP87 (complementary to RP86) were used in an asymmetric temperature cycling reaction with Pwo polymerase and plasmid pRKP1613 as the template. Construct pRKP1613 is the pGEM-7zf(+) cloning vector (Promega) carrying the 628 bp BamHI–SacI fragment from pPL341 (Vasudevan et al., 1991). The parental (methylated) plasmid DNA was then removed by treatment with DpnI, and the remaining DNA was amplified in E.coli. The resulting plasmid pRKP1614 was confirmed by VspI restriction and sequencing. Purified 335 bp SacI–FokI DNA fragments from pRKP1613 and pRKP1614 (Phmp) were labelled with [α-32P]dCTP and Klenow polymerase (Promega). For FNR–DNA interaction studies, purified labelled fragments were left at least overnight in the anaerobic box.
FNR–DNA interactions
Anaerobic gel retardation analyses were as in Wu et al. (2000), except that the 32P-labelled DNA fragment used was Phmp (0.732 pmol in 10 µl). Reconstituted FNR protein was tested for DNA-binding activity after titration with air-saturated TCB or with an NO solution. After drying, gels were applied to a Bio-Rad phosphorimager model GS-363 and analyzed by Molecular Image® System (Bio-Rad) with the local background redefined as zero.
Repressor–operator interaction analyses were performed essentially as in Riggs et al. (1970). Dilutions of FNR protein were mixed with 32P-labelled Phmp in anoxic TCB supplemented with bovine serum albumin (50 µg/ml) and salmon sperm DNA (10 µg/ml) in a final volume of 100 µl. Reactions were incubated for 10 min, before 2 ml of TCB was added to the tubes. After mixing, two reaction samples (1 ml) were filtered through cellulose nitrate filters (25 mm diameter, 0.45 µm pore size; Schleicher and Schuell) previously soaked for 30 min in anoxic TCB. Filters were washed once with 0.5 ml of TCB to eliminate free Phmp then subjected to scintillation counting. The fraction of FNR–Phmp complex was plotted versus the log of FNR concentration, according to the equation Kd = [FNR][DNA]/[FNR·DNA], where [FNR]>>[DNA]. The apparent Kd for the individual assays is the concentration of FNR (as monomer) giving half-maximal binding; the mean of two sets of experiments was taken.
DNase I footprinting reactions of Phmp were carried out anaerobically using anoxic solutions of the Core Footprinting System (Promega) and following the manufacturer’s instructions. Reactions with FNR protein and RNAP (USB) were loaded onto an 8% polyacrylamide sequencing gel. The SacI–FokI fragment from pRKP1613 was also cleaved by dimethyl sulfate/piperidine hydrolysis for use as a DNA sequence ladder.
Acknowledgments
Acknowledgements
We thank Dr R.C.Read (University of Sheffield Medical School) for advice on macrophages. We also thank the BBSRC for supporting this work under the PRES initiative with research grants to R.K.P., M.N.H. and H.C.-R. (PRS12199), and to J.G. and A.J.T. (PRS12148). J.C. held a BBSRC Studentship.
References
- Bouton C., Hirling,H. and Drapier,J.-C. (1997) Modulation of iron regulatory protein functions. Further insights into the role of nitrogen- and oxygen-derived reactive species. J. Biol. Chem., 272, 19969–19975. [DOI] [PubMed] [Google Scholar]
- Chung M.H., Kasai,H., Jones,D.S., Inoue,H., Ishikawa,H., Otsuka,E. and Nishimura,S. (1991) An endonuclease activity of Escherichia coli that specifically removes 8-hydroxyguanine residues from DNA. Mutat. Res., 254, 1–12. [DOI] [PubMed] [Google Scholar]
- Costanzo S., Ménage,S., Purrello,R., Bonomo,R.P. and Fontecave M. (2001) Re-examination of the formation of dinitrosyl-iron complexes during reaction of S-nitrosothiols with Fe(II). Inorg. Chim. Acta, 318, 1–7. [Google Scholar]
- Crawford W.J. and Goldberg,D.E. (1998) Regulation of the Salmonella typhimurium flavohemoglobin gene. A new pathway for bacterial gene expression in response to nitric oxide. J. Biol. Chem., 273, 34028–34032. [DOI] [PubMed] [Google Scholar]
- Cruz-Ramos H., Hughes,M.N. and Poole,R.K. (2000) Nitric oxide is an effector in bacterial gene expression mediated by FNR and FNR-like proteins. In Moncada,S., Gustafsson,L.E., Wiklund,N.P. and Higgs,E.A (eds), The Biology of Nitric Oxide Part 7. Portland Press, London, UK, p. 158.
- De Groote M.A. et al. (1997) Periplasmic superoxide dismutase protects Salmonella from products of phagocyte NADPH-oxidase and nitric oxide synthase. Proc. Natl Acad. Sci. USA, 94, 13997–14001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ding H. and Demple,B. (2000) Direct nitric oxide signal transduction via nitrosylation of iron–sulfur centers in the SoxR transcription activator. Proc. Natl Acad. Sci. USA, 97, 5146–5150. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gardner P.R., Gardner,A.M., Martin,L.A. and Salzman,A.L. (1998a) Nitric oxide dioxygenase: an enzymic function for flavohemoglobin. Proc. Natl Acad. Sci. USA, 95, 10378–10383. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gardner P.R., Costantino,G. and Salzman,A.L. (1998b) Constitutive and adaptive detoxification of nitric oxide in Escherichia coli. Role of nitric-oxide dioxygenase in the protection of aconitase. J. Biol. Chem., 273, 26528–26533. [DOI] [PubMed] [Google Scholar]
- Gobert A.P., McGee,D.J., Akhtar,M., Mendz,G.L., Newton,J.C., Cheng,Y., Mobley,H.L.T. and Wilson,K.T. (2001) Helicobacter pylori arginase inhibits nitric oxide production by eukaryotic cells: a strategy for bacterial survival. Proc. Natl Acad. Sci. USA, 98, 13844–13849. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gort A.S., Ferber,D.M. and Imlay,J.A. (1999) The regulation and role of the periplasmic copper, zinc superoxide dismutase of Escherichia coli. Mol. Microbiol., 32, 179–191. [DOI] [PubMed] [Google Scholar]
- Green J., Bennett,B., Jordan,P., Ralph,E.T., Thomson,A.J. and Guest,J.R. (1996) Reconstitution of the [4Fe–4S] cluster in FNR and demonstration of the aerobic–anaerobic transcription switch in vitro. Biochem. J., 316, 887–892. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Green J., Scott,C. and Guest,J.R. (2001) Functional versatility in the CRP-FNR superfamily of transcription factors: FNR and FLP. In Poole,R.K. (ed.), Advances in Microbial Physiology. Academic Press, London, UK, Vol. 44, pp. 1–34 [DOI] [PubMed] [Google Scholar]
- Guest J.R., Green,J., Irvine,A.S. and Spiro,S. (1996) The FNR modulon and FNR-regulated gene expression. In Lin,E.C.C. and Lynch,A.S. (eds), Regulation of Gene Expression in Escherichia coli. R.G.Landes Company, Georgetown, TX, pp. 317–342. [Google Scholar]
- Hassan H.M. and Sun,H.C.H. (1992) Regulatory roles of Fnr, Fur and Arc in expression of manganese-containing superoxide dismutase in Escherichia coli. Proc. Natl Acad. Sci. USA, 89, 3217–3221. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hausladen A., Privalle,C.T., Keng,T., DeAngello,J. and Stamler,J.S. (1996) Nitrosative stress: activation of the transcription factor OxyR. Cell, 86, 719–729. [DOI] [PubMed] [Google Scholar]
- Hausladen A., Gow,A.J. and Stamler,J.S. (1998) Nitrosative stress: metabolic pathway involving the flavohemoglobin. Proc. Natl Acad. Sci. USA, 95, 14100–14105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hausladen A., Gow,A. and Stamler,J.S. (2001) Flavohemoglobin denitrosylase catalyzes the reaction of a nitroxyl equivalent with molecular oxygen. Proc. Natl Acad. Sci. USA, 98, 10108–10112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hentze W.M. and Kühn,L.C. (1996) Molecular control of vertebrate iron metabolism: mRNA-based regulatory circuits operated by iron, nitric oxide and oxidative stress. Proc. Natl Acad. Sci. USA, 93, 8175–8182. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hutchings M.I., Crack,J.C., Shearer,N., Thompson,B.J., Thomson,A.J. and Spiro,S. (2002) Transcriptional factor FnrP from Paracoccus denitrificans contains an iron–sulfur cluster and is activated by anoxia: identification of essential cysteine residues. J. Bacteriol., 184, 503–508. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jordan P.A., Thomson,A.J., Ralph,E.T., Guest,J.R. and Green,J. (1997) FNR is a direct oxygen sensor having a biphasic response curve. FEBS Lett., 416, 349–352. [DOI] [PubMed] [Google Scholar]
- Kennedy M.C., Antholine,W.E. and Beinert,H. (1997) An EPR investigation of the products of the reaction of cytosolic and mitochondrial aconitases with nitric oxide. J. Biol. Chem., 272, 20340–20347. [DOI] [PubMed] [Google Scholar]
- Khoroshilova N., Beinert,H. and Kiley,P.J. (1995) Association of a polynuclear iron–sulfur center with a mutated FNR protein enhances DNA binding. Proc. Natl Acad. Sci. USA, 92, 2499–2503. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Khoroshilova N., Popescu,C., Münck,E., Beinert,H. and Kiley,P.J. (1997) Iron–sulfur cluster disassembly in the FNR protein of Escherichia coli by O2: [4Fe–4S] to [2Fe-2S] conversion with loss of biological activity. Proc. Natl Acad. Sci. USA, 94, 6087–6092. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kiley P.J. and Beinert,H. (1999) Oxygen sensing by the global regulator, FNR, the role of the iron–sulfur cluster. FEMS Microbiol. Rev., 22, 341–352. [DOI] [PubMed] [Google Scholar]
- Kim S.O., Orii,Y., Lloyd,D., Hughes,M.N. and Poole,R.K. (1999) Anoxic function for the Escherichia coli flavohaemoglobin (Hmp): reversible binding of nitric oxide and reduction to nitrous oxide. FEBS Lett., 445, 389–394. [DOI] [PubMed] [Google Scholar]
- Krieglstein C.F., Cerwinka,W.H., Laroux,F.S., Salter,J.W., Rusell,J.M., Schuermann,G., Grisham,M.B., Ross,C.R. and Granger,D.N. (2001) Regulation of murine intestinal inflammation by reactive metabolites of oxygen and nitrogen: divergent roles of superoxide and nitric oxide. J. Exp. Med., 194, 1207–1218. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lazzazera B.A., Beinert,H., Khoroshilova,N., Kennedy,C. and Kiley,P.J. (1996) DNA binding and dimerization of the Fe–S-containing FNR protein from Escherichia coli are regulated by oxygen. J. Biol. Chem., 271, 2762–2768. [DOI] [PubMed] [Google Scholar]
- Lee H.-S., Lee,Y.S., Kim,H.-S., Choi,J.-Y., Hassan,H.M. and Sun,H.C.H. (1998) Mechanism of regulation of 8-hydroxyguanine endonuclease by oxidative stress: roles of fnr, arcA, and fur. Free Radic. Biol. Med., 24, 1193–1201. [DOI] [PubMed] [Google Scholar]
- Marshall F.A., Messenger,S.L., Wyborn,N.R., Guest,J.R., Wing,H., Busby,S.J.W. and Green,J. (2001) A novel promoter architecture for microaerobic activation by the anaerobic transcription factor FNR. Mol. Microbiol., 39, 747–753. [DOI] [PubMed] [Google Scholar]
- Membrillo-Hernández J., Coopamah,M.D., Channa,A., Hughes,M.N. and Poole,R.K. (1998) A novel mechanism for upregulation of the Escherichia coli K-12 hmp (flavohaemoglobin) gene by the ‘NO releaser’, S-nitrosoglutathione: nitrosation of homocysteine and modulation of MetR binding to the glyA-hmp intergenic region. Mol. Microbiol., 29, 1101–1112. [DOI] [PubMed] [Google Scholar]
- Membrillo-Hernández J., Coopamah,M.D., Anjum,M.F., Stevanin,T.M., Kelly,A., Hughes,M.N. and Poole,R.K. (1999) The flavohemoglobin of Escherichia coli confers resistance to a nitrosating agent, a ‘nitric oxide releaser’ and paraquat and is essential for transcriptional responses to oxidative stress. J. Biol. Chem., 274, 748–754. [DOI] [PubMed] [Google Scholar]
- Meng W., Green,J. and Guest,J.R. (1997) FNR-dependent repression of ndh gene expression requires two upstream FNR-binding sites. Microbiology, 143, 1521–1532. [DOI] [PubMed] [Google Scholar]
- Mills C.E., Sedelnikova,S., Søballe,B., Hughes,M.N. and Poole,R.K. (2001) Escherichia coli flavohaemoglobin (Hmp) with equi stoichiometric FAD and haem contents has a low affinity for dioxygen in the absence or presence of nitric oxide. Biochem. J., 353, 207–213. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nunoshiba T., de Rojas-Walker,T., Wishnok,J.S., Tannenbaum,S.R. and Demple,B. (1993) Activation by nitric oxide of an oxidative-stress response that defends Escherichia coli against activated macrophages. Proc. Natl Acad. Sci. USA, 90, 9993–9997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Oliveira L., Bouton,C. and Drapier,J.-C. (1999) Thioredoxin activation of iron regulatory proteins. Redox regulation of RNA binding after exposure to nitric oxide. J. Biol. Chem., 274, 516–521. [DOI] [PubMed] [Google Scholar]
- Pantopoulos K. and Hentze,M.W. (1995) Nitric oxide signaling to iron-regulatory protein: direct control of ferritin mRNA translation and transferrin receptor mRNA stability in transfected fibroblasts. Proc. Natl Acad. Sci. USA, 92, 1267–1271. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Poole R.K. and Hughes,M.N. (2000) New functions for the ancient globin family: bacterial responses to nitric oxide and nitrosative stress. Mol. Microbiol., 36, 775–783. [DOI] [PubMed] [Google Scholar]
- Poole R.K., Anjum,M.F., Membrillo-Hernández,J., Kim,S.O., Hughes, M.N. and Stewart,V. (1996) Nitric oxide, nitrite and Fnr regulation of hmp (flavohemoglobin) gene expression in Escherichia coli K-12. J. Bacteriol., 178, 5487–5492. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Richardson D.R. and Ponka,P. (1997) The molecular mechanisms of the metabolism and transport of iron in normal and neoplastic cells. Biochim. Biophys. Acta, 1331, 1–40. [DOI] [PubMed] [Google Scholar]
- Riggs A.D., Suzuki,H. and Bourgeois,S. (1970) Lac repressor–operator interaction. I. Equilibrium studies. J. Mol. Biol., 48, 67–83. [DOI] [PubMed] [Google Scholar]
- Stevanin T.M., Ioannidis,N., Mills,C.E., Kim,S.O., Hughes,M.N. and Poole,R.K. (2000) Flavohemoglobin Hmp affords inducible protection for Escherichia coli respiration, catalyzed by cytochromes bo′ or bd, from nitric oxide. J. Biol. Chem., 275, 35868–35875. [DOI] [PubMed] [Google Scholar]
- Stevanin T.M., Poole,R.K., Demoncheux,E.A.G. and Read,R.C. (2002) Flavohemoglobin Hmp protects Salmonella from nitric oxide-related killing by human macrophages. Infect. Immun., in press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Unden G. and Duchêne,A. (1987) On the role of cyclic AMP and the Fnr protein in Escherichia coli growing anaerobically. Arch. Microbiol., 147, 195–200. [DOI] [PubMed] [Google Scholar]
- Vanin A.F., Serezhenkov,A., Mikoyan,V.D. and Genkin,M.V. (1998) The 2.03 signal as an indicator of dinitrosyl-iron complexes with thiol-containing ligands. Nitric Oxide, 2, 224–234. [DOI] [PubMed] [Google Scholar]
- Vasudevan S.G., Armarego,W.L.F., Shaw,D.C., Lilley,P.E., Dixon,N.E. and Poole,R.K. (1991) Isolation and nucleotide sequence of the hmp gene that encodes a haemoglobin-like protein in Escherichia coli K-12. Mol. Gen. Genet., 226, 49–58. [DOI] [PubMed] [Google Scholar]
- Wardrop S.L., Watts,R.N. and Richardson,D.R. (2000) Nitrogen monoxide activates iron regulatory protein 1 RNA-binding activity by two possible mechanisms: effect on the [4Fe–4S] cluster and iron mobilization from cells. Biochemistry, 39, 2748–2758. [DOI] [PubMed] [Google Scholar]
- Williams S.M., Wing,H.J. and Busby,S.J.W. (1998) Repression of transcription initiation by Escherichia coli FNR protein: repression by FNR can be simple. FEMS Microbiol. Lett., 163, 203–208. [DOI] [PubMed] [Google Scholar]
- Wink D.A. et al. (1991) DNA deaminating activity and genotoxicity of nitric oxide and its progenitors. Science, 254, 1001–1003. [DOI] [PubMed] [Google Scholar]
- Wu G., Cruz-Ramos,H., Hill,S., Green,J., Sawers,G. and Poole,R.K. (2000) Regulation of cytochrome bd expression in the obligate aerobe Azotobacter vinelandii by CydR (Fnr). Sensitivity to oxygen, reactive oxygen species and nitric oxide. J. Biol. Chem., 275, 4679–4686. [DOI] [PubMed] [Google Scholar]