

Abstract
The outermost layer of Mycobacterium tuberculosis is composed primarily of two polysaccharides, glucan (GC) and arabinomannan. To analyze the surface polysaccharide composition of M. tuberculosis, we generated a monoclonal antibody (MAb) that binds M. tuberculosis GC and is known as MAb 24c5. Immunofluorescence and whole-mount immunoelectron microscopy indicated that GC is on the outermost portion of the bacteria. M. tuberculosis strains Erdman and CDC 1551 were analyzed for their ability to bind MAb 24c5 after in vitro growth in media with and without the detergent Tween 80. MAb 24c5 bound to Erdman and CDC 1551 at all culture times with only slightly greater apparent affinity after extended culture in the absence of Tween 80, indicating that a stable amount of GC polysaccharide antigen is associated with the cell surface of M. tuberculosis. An enzyme-linked immunosorbent assay indicated that GC is antigenically similar to glycogen, and the amount of GC antigen increased in the media of M. tuberculosis cultures grown either with or without the detergent Tween 80. Other nontuberculosis mycobacteria have antigenically similar GCs on their surfaces after in vitro growth. Inoculation of mice with live bacilli but not inoculation with dead bacilli elicited a strong antibody response to GC consistent with production of this antigen in vivo. Our results provide a more comprehensive picture of the M. tuberculosis cell envelope and the conditions that allow expression of M. tuberculosis GC.
In the early 1990s it was reported that one-third of all Mycobacterium tuberculosis patient isolates in the New York City area were drug resistant, that the number of M. tuberculosis cases in the United States had been increasing since 1985, and that a large percentage of the avoidable adult deaths in the world were caused by tuberculosis (TB) (reviewed in reference 1). With the introduction of directly observed therapy, the problems in the United States have become less severe, but it is estimated that one-third of the human population is infected with M. tuberculosis, and TB remains a leading cause of death worldwide (approximately 2 million deaths per annum) (12). Nearly one-half of the TB cases occur in five Southeast Asian countries (12). There is a desperate need for effective and affordable modalities capable of combating M. tuberculosis, including vaccines. There is considerable evidence that antibody to the M. tuberculosis polysaccharides can modify the course of infection to the benefit of the host (15). Recently, a monoclonal antibody (MAb) to mycobacterial arabinomannan (AM) was shown to prolong survival when it was incubated with M. tuberculosis before infection of mice (33). These findings suggest that M. tuberculosis carbohydrates could include protective antigens.
The outermost portion of the M. tuberculosis surface is composed mainly of polysaccharide, and there is a small amount of protein and almost no detectable lipid (reviewed in reference 7). This layer may correspond to the electron-transparent zone found immediately outside the mycobacterial cell wall by electron microscopy (7). Fibrillar structures present in the electron-transparent zone of intracellular mycobacteria were demonstrated to be glucose rich after in vitro growth (11, 18). Recently, Lemassu and Daffe demonstrated that the two major components of the M. tuberculosis outermost layer are the polysaccharides AM and glucan (GC) (21). These polysaccharides accumulate in the supernatant of M. tuberculosis cultures (9). GC was probably the polysaccharide II fraction identified by Seibert in 1949 as a “large polysaccharide molecule not hitherto described and found in quantity only in culture filtrates of certain strains of tubercle bacilli, viz. H 37, A 33 and BCG…giving the culture filtrates an opalescence” (28). Further characterization by Lemassu and Daffe in the 1990s revealed that GC has a structure similar to that of mycobacterial cytosolic glycogen, although it has a lower molecular mass, approximately 100 kDa (21).
At present, the location of GC on the cell envelope and the amount of GC expressed as a function of M. tuberculosis strain, as a function of growth conditions, or as a function of time are poorly understood. In addition, it is not known whether GC is an immunogenic antigen as a result of infection. In this study we report the generation of monoclonal and polyclonal antibodies to GC and their use to study M. tuberculosis GC expression in vitro and in vivo.
(The data in this paper are from a thesis to be submitted by J. R. Schwebach in partial fulfillment of the requirements for the degree of Doctor of Philosophy from the Sue Golding Graduate Division of Medical Sciences, Albert Einstein College of Medicine, Yeshiva University, Bronx, N.Y.)
MATERIALS AND METHODS
Culture of mycobacteria.
M. tuberculosis strains Erdman and CDC 1551 were obtained from the laboratory of Barry Bloom (Harvard School of Public Health, Boston, Mass.) and grown in Middlebrook 7H9 medium (Difco, Detroit, Mich.) containing 5 ml of glycerol (Sigma, St. Louis, Mo.) per liter. Mycobacterial cultures were grown with (7H9-T medium) or without (7H9 medium) 0.05% Tween 80 (Sigma) for 11, 20, and 25 days in 490-cm2 roller bottles (Corning Inc., Corning, N.Y.) in a 5% CO2 incubator at 37°C at pathogen level 3, as described by Schwebach et al. (26). The number of bacteria used was standardized according to the amount of protein in 100 μl of sedimented bacteria after growth as described previously (26). M. tuberculosis samples were heat killed at 80°C for 2 h, washed twice in TBS (0.1 M Tris base [pH 7.2], 0.15 M NaCl, 0.001 M NaN3), briefly sonicated, and diluted in TBS to a concentration of 9.3 μg of protein per ml as described by Schwebach et al. (26) before use in enzyme-linked immunosorbent assays (ELISA). M. kansasii, M. bovis BCG Pasteur, M. smegmatis mc2155, M. phlei mc219, M. fortuitum mc247, and M. avium mc272 were grown in Sauton's medium as described by Lemassu et al. (22). Cultures of these nontuberculosis mycobacteria were concentrated by sedimentation and heat killed as described previously (26). Aliquots (100 μl) of sedimented bacteria were then washed twice in TBS, brought to a volume of 35 ml in TBS, and sonicated with three 5-s bursts by using a probe tip at 70% output (setting, high; value, 280; Braun Sonic 2000 U Sonifier). An additional 15 ml of TBS was then added to each sample prior to use in ELISA.
M. tuberculosis GC.
M. tuberculosis was isolated from a patient with confirmed TB and pleural effusion at Montefiore Medical Center (Bronx, N.Y.). GC from liquid culture supernatant (Sauton's medium) of this strain was isolated by precipitation with 70% ethanol and was further purified by anion-exchange and gel filtration chromatography, using methods similar to those described by Lemassu and Daffe (21). The GC contained only glucose, as confirmed by gas-liquid chromatography and 13C magnetic resonance spectroscopy.
Generation of a GC-binding MAb.
1-Cyano-4-dimethylaminopyridinium tetratfluoroborate (CDAP) chemistry was used to conjugate GC to recombinant Pseudomonas aeruginosa exoprotein A (rEPA) as described by Konadu et al. (20). Female 6- to 8-week-old BALB/cJ mice were immunized by intraperitoneal injection of 5 μg of GC-rEPA in 200 μl of 50% Freund's incomplete adjuvant (Sigma) in phosphate-buffered saline (PBS). Animals were given a booster dose with 5 μg of GC-rEPA in 200 μl of PBS at 2 and 4 weeks. The mice were bled from the retroorbital plexus 2, 4, and 6 weeks after the initial immunization. Binding of the sera to GC was measured by ELISA, as described below. The mouse with the highest titer was given a booster dose at 8 weeks with 5 μg of GC-rEPA in 200 μl of PBS, and the spleen was harvested 3 days later. Hybridomas were generated as described by de St. Groth and Scheidegger (10). Briefly, spleen cells were fused with NSO cells at a ratio of 4:1, suspended in medium containing hypoxanthine-aminopterin-thymidine (Sigma), plated in 20 96-well culture plates, and incubated at 37°C with 10% CO2. Supernatants from wells containing hybridomas were screened for production of antibody to GC using ELISA. The hybridoma designated 24c5 was recovered and cloned twice on soft agar. The isotype of the antibody was determined by ELISA using isotype-specific secondary antibodies (Southern Biotechnology Associates, Birmingham, Ala.) in the GC ELISA. Supernatant from the 24c5 hybridoma was concentrated by using an Amicon 8400 concentrator and a YM100 membrane (Amicon/Millipore, Bedford, Mass.) under compressed N2. The antibody concentration was determined by ELISA by comparison to a standardized amount of isotype-matched control antibody (ICN Biomedicals, Aurora, Ohio).
Whole-cell mycobacterium ELISA.
An ELISA of M. tuberculosis and other mycobacteria was performed as described previously (26). After washing of bovine serum albumin (BSA)-blocked microtiter plates containing whole-cell mycobacteria, MAb 24c5 was added in TBS, and the plates were incubated for 1 h at 37°C. After the plates were washed, 1 μg of goat anti-mouse alkaline phosphatase-conjugated (GAM-AP) immunoglobulin G1 (IgG1) antibody (Southern Biotechnology Associates) per ml in TBS was added to each plate, and the plates were incubated for 1 h at 37°C and then developed using p-nitrophenylphosphate (Sigma). The M. tuberculosis ELISA plates were processed simultaneously. ELISA measurements were obtained in triplicate for each sample and averaged. Experiments were performed twice (including culture of M. tuberculosis), with similar results.
GC and glycogen ELISA.
Polystyrene microtiter plate wells were coated with 50 μl of GC (10 μg/ml) in carbonate buffer (0.015 M Na2CO3, 0.035 M NaHCO3, 0.003 M NaN3; pH 9.8) by incubating the plates overnight at 4°C. The wells were then blocked by adding 200 μl of 3% BSA in TBS and incubating the plates overnight at 4°C. MAb 24c5 or serum from M. tuberculosis-infected mice was added to the wells and incubated for 1 h at 37°C. For determination of the murine IgG titer to GC, serum was diluted in TBS with 0.075 M β-mercaptoethanol and heated for 1 h at 37°C to dissociate IgM pentamers, and the solution was immediately assayed for IgG reactivity to GC by ELISA. The plates were then washed, and 50 μl of a 1-μg/ml solution of GAM-AP antibody (GAM-AP IgG1 antibody recognizing MAb 24c5 and GAM-AP IgG or IgM antibody for murine serum) was diluted in TBS and added to each well for 1 h at 37°C. The ELISA plates were washed and developed by using p-nitrophenylphosphate substrate, as described by Schwebach et al. (26). Type VIII (slipper limpet) and type IX (bovine liver) glycogens (Sigma) were used in ELISA at concentrations of 1, 10, and 100 μg/ml; the conditions used were identical to those used for GC ELISA.
ELISA of M. tuberculosis extracts and cell wall fractions.
For binding an interphase extract of M. tuberculosis culture supernatant known to contain GC, 300 ml of a day-25 M. tuberculosis culture supernatant was filtered twice with a 0.2-μm-pore-size Millipore filter (Amicon/Millipore) and vacuum concentrated 10-fold at room temperature (RT). The concentrated supernatant was extracted with chloroform-methanol-water (3:4:3, vol/vol/vol). The resulting interphase was collected for ELISA analysis by extraction with 10 ml of distilled H2O. One hundred microliters of the extracted interphase was mixed with 800 μl of TBS and incubated for 1 h at 37°C in 16 wells (50 μl per well) of a 96-well polystyrene microplate (Corning) before blocking with 3% BSA-TBS overnight. An ELISA using MAb 24c5 was then performed with this preparation and all preparations described below by diluting MAb 24c5 threefold from a starting concentration of 100 μg/ml. ELISA of M. tuberculosis and GC (1 μg/ml) were performed as described above; for the M. tuberculosis ELISA M. tuberculosis Erdman cells grown for 20 days in 7H9 medium were used. An AM (10 μg/ml) ELISA was performed as described by Schwebach et al. (26). MAb binding to purified M. tuberculosis Erdman lipoarabinomannan (10 μg/ml), phosphatidylinositol mannoside (10 μg/ml), mycolyl-arabinogalactan-peptidoglycan complex (1 mg/ml), total lipid fraction (1 mg/ml), and fast-growing mycobacterial lipomannan (10 μg/ml) was examined as described by Glatman-Freedman et al. using ELISA (16). These antigens were kindly supplied by P. J. Brennan and J. T. Belisle (Department of Microbiology, Colorado State University, Fort Collins). M. tuberculosis AM was isolated as described previously (26). GAM-AP IgG1 secondary antibody was used in all these ELISA.
ELISA of proteinase K-treated M. tuberculosis.
Treatment of M. tuberculosis with proteinase K during ELISA was done as previously described (16). Briefly, cells of M. tuberculosis Erdman grown in 7H9 medium for 20 days were added to ELISA plates as described above. The M. tuberculosis was then treated with 1 mg of proteinase K (Boehringer Mannheim Biochemicals, Indianapolis, Ind.) per ml in PBS or with PBS alone (as a control) at RT for 20 h before continuation of the ELISA as described above. This enzyme preparation was active in similar experiments used to digest organ homogenate (26).
IF.
For immunofluorescence (IF) studies, MAb 24c5 and the irrelevant isotype-matched control MAb 2D10 (recognizing Cryptococcus neoformans glucuronoxylomannan) were purified from murine ascites fluid using a protein G column (Pierce, Rockford, Ill.). The purified antibodies were then conjugated to the ALEXA-488 fluorophore (Molecular Probes, Eugene, Oreg.) by following the manufacturer's directions. Cells of M. tuberculosis Erdman grown for 25 days in 7H9 medium were centrifuged (2,000 × g, 8 min), washed in PBS, and fixed in 5% PBS-buffered formalin at room temperature for several days (which is a safe method for M. tuberculosis inactivation [27]). The fixed cells were washed three times in PBS. The M. tuberculosis-PBS suspension was then placed on poly-l-lysine slides (Sigma) and air dried overnight. The slides were then blocked with 1% BSA-TBS for 1 h at RT. The blocking solution was then removed, and a 1:100 dilution (10 μg/ml) of conjugated MAb (or conjugated irrelevant control) in 1% BSA-TBS was added to the slides. The slides were then incubated for 1 h at 37°C and washed three times with TBS. Coverslips were placed on the slides before viewing with an AX-70 fluorescence microscope (Olympus, Melville, N.Y.).
Electron microscopy.
For whole-mount transmission electron microscopy, M. tuberculosis Erdman was grown for 25 days in 7H9 medium, collected by centrifugation (2,000 × g, 8 min), washed twice with 0.1 M phosphate buffer (PB) (Fisher), and collected by centrifugation again. A solution containing 1 mg of poly-l-lysine (Sigma) per ml in PB was applied to Formvar/carbon-coated 300-mesh nickel grids (Polysciences, Inc., Warrington, Pa.) for 5 min. Bacteria were then suspended in PB and allowed to settle onto the grids for 15 min. Bacteria were fixed onto the grids using 2.5% glutaraldehyde (Polysciences) in PB. The grids were washed five times (3 min each) in PB and blocked (10 min) in 0.26 M NH4Cl (Fisher) before further blocking for 15 min in 0.5% BSA-1% coldwater fish gelatin PB (BSA-gel PB) (Aurion, Wageningen, The Netherlands). Hybridoma supernatants containing either MAb 24c5 or the irrelevant isotype-matched MAb control (MAb C293, recognizing dipeptidyl peptidase V of Aspergillus fumigatus, kindly provided by M. Feldmesser) were added to BSA-gel PB at a final concentration of 100 μg/ml, and grids were incubated in these solutions for 2 h at RT. The grids were washed five times (3 min each) in BSA-gel PB and transferred to BSA-gel PB containing 0.05% goat anti-mouse IgG conjugated to 5-nm gold (Aurion) for 1 h at 37°C. The grids were washed in BSA-gel PB three times (3 min each) and then in PB three times (3 min each), and then they were fixed overnight by incubation in PB containing 2.5% glutaraldehyde and 2% paraformaldehyde (Electron Microscopy Sciences, Fort Washington, Pa.) at 4°C. The fixed grids were moved to pathogen level 2 for washing in PB (five times, 5 min each) and in distilled H2O (five times, 5 min each) and staining for 90 s with 1% phosphotungstic acid (Fisher). Excess phosphotungstic acid was removed from the grids using Whatman no. 5 filter paper, and the grids were viewed with a 1200EX transmission electron microscope (JOEL, Peabody, Mass.) at an accelerating voltage of 80 kV.
GC capture ELISA.
Capture ELISA were used to detect GC antigen in culture supernatants. These assays were modeled on capture assays developed for detection of C. neoformans glucuronoxylomannan (3), M. tuberculosis lipoarabinomannan (17), and M. tuberculosis AM (26). Serum from a female New Zealand White rabbit immunized intraperitoneally with 50 μg of GC-rEPA in Freund's incomplete adjuvant (Sigma) was used for detection of GC captured by MAb 24c5. ELISA plates were first incubated with 50 μl of TBS containing 2 μg of unlabeled goat anti-mouse IgG1 antibody per ml for 1 h at 37°C. The wells were blocked by adding 200 μl of 1% BSA in TBS and incubating the plates overnight at 4°C. The plates were then washed between steps with TBS containing 0.05% Tween 20 (Sigma) as described above. After the washing, 1 μg of MAb 24c5 per ml in TBS was added to each well of the 96-well plates, and the plates were incubated for 1 h at 37°C. Aliquots of concentrated M. tuberculosis culture supernatant were added to the plates after the washing. Purified GC, added to 7H9 medium, 7H9-T medium, or TBS and serially diluted in TBS, was used as a standard and a positive control. The same media without purified GC were also used as negative controls. Following incubation at 37°C for 1 h, day 7 rabbit serum (stored frozen at −20°C) diluted 1:100 in TBS was added to the 96-well plates. After 1 h of incubation at 37°C, the plates were washed, 1 μg of goat anti-rabbit alkaline phosphatase-conjugated IgM (Southern Biotechnology Associates) per ml in TBS was added, and the assay mixture was developed as described above. An absorbance signal that was ≥1.5 times the background signal was considered positive for detection of GC.
Serum collection from M. tuberculosis-infected mice.
Six- to eight-week-old female BALB/cAnNCrlBR mice (Charles River Laboratories, Wilmington, Mass.) were each infected with 1 × 106 Erdman bacilli in 200 μl of PBS by intravenous inoculation. Just prior to injection, mycobacteria from a frozen vial for which the concentration (CFU per milliliter) was known were thawed and added to 4 ml of PBS containing 0.05% Tween 80 (Sigma) in a 15-ml conical tube. The suspension was then sonicated for 5 s at 60% output (constant duty cycle) with a cup-horn sonicator (Branson, Danbury, Conn.) before inversion and sonication again for 10 s. Three mice were bled from the retroorbital plexus to collect serum at zero time (prior to infection) and 7, 20, and 42 days after infection. The serum was diluted in TBS containing 1 mM azide, filtered twice through a 0.2-μm-pore-size syringe filter, and removed from biohazard safety level 3 for storage at 4°C until analysis by the GC ELISA. An equal number of mice were infused with 1 × 106 heat-killed Erdman bacilli, and serum was collected as described above. The heat-killed bacterium used for these infusions was prepared as described above, except that the thawed vial of M. tuberculosis was heated for 2 h at 80°C prior to injection to ensure that the mycobacterium was killed.
RESULTS AND DISCUSSION
Generation of an MAb to GC.
In this study an M. tuberculosis conjugate vaccine was employed to elicit an antibody response to GC in mice from which a hybridoma producing a GC-binding MAb was generated. Administration of 5 μg of GC-rEPA conjugate in Freund's incomplete adjuvant to BALB/c mice followed by a booster dose of with 5 μg of GC-rEPA in PBS at day 14 elicited IgG to M. tuberculosis GC (Fig. 1). The titer increased after a second booster dose of with 5 μg of GC-rEPA in PBS at day 28. At week 6 after the initial immunization with GC-rEPA, splenocytes from the mouse with the highest serum reactivity (Fig. 1, mouse 4) to GC were fused to NSO myeloma cells, and a single IgG1 MAb (MAb 24c5) was recovered that bound to GC, as determined by ELISA (Fig. 2A). The amount of reactivity was a function of both MAb 24c5 concentration and the amount of GC used to coat the plate (Fig. 2A and B). When a constant amount of MAb 24c5 (1 μg/ml) was tested with variable concentrations of GC, the lowest signal above the background level (1.5 times the background level) was measured in wells coated with 0.63 μg of GC per ml (Fig. 2B), indicating that MAb 24c5 has good ability to bind M. tuberculosis GC.
FIG. 1.

Immunogenicity of GC-rEPA: anti-GC IgG titers of four different mice after immunization and boosting with GC-rEPA. A diagram of the ELISA used to determine titers is on the right. Alk-Phos, alkaline phosphatase.
FIG. 2.

MAb 24c5 ELISA analysis and binding to glycogen. (A) Splenic fusion resulted in MAb 24c5, an IgG1 able to recognize M. tuberculosis GC. Each data point represents the average of three measurements. (B) MAb 24c5 binding to dilute concentrations of GC in the M. tuberculosis GC ELISA. Each data point represents the average of three measurements. (C) MAb 24c5 binding to M. tuberculosis Erdman day-20 7H9 medium (MTB), cell culture supernatant extract (INTPHSE), GC, AM, lipoarabinomannan (LAM), phosphatidylinositol mannoside (PIM), fast-growing mycobacterial lipomannan (LM), mycolyl-arabinogalactan-peptidoglycan complex (mAGP), and total lipid fraction (TLF). Each bar represents the average of two measurements. (D) Effects of proteinase K on binding of MAb 24c5 to M. tuberculosis Erdman day-20 7H9 medium. (E and F) MAb 24c5 assayed for reactivity to type VIII (slipper limpet) (viii) (E) and type IX (ix) (bovine liver) (F) glycogens. Each data point represents the average of three measurements. Error bars show the standard deviations of the means.
MAb 24c5 specifically binds M. tuberculosis GC.
The specificity of MAb 24c5 was explored by testing the reactivity of this MAb with purified M. tuberculosis cell wall fractions (Fig. 2C). Binding of MAb 24c5 was limited to GC and the GC-rich interphase (21) recovered from a chloroform-methanol extract of M. tuberculosis Erdman supernatant (Fig. 2C). MAb 24c5 did not bind M. tuberculosis AM, lipoarabinomannan, phosphatidylinositol mannoside, mycolyl-arabinogalactan-peptidoglycan, total lipid fraction, or mycobacterial lipomannan, which are components of the remainder of the nonprotein M. tuberculosis cell surface (2, 4, 7). In addition, MAb 24c5 did not react with an M. tuberculosis protein antigen(s), as treatment of M. tuberculosis whole cells with proteinase K did not reduce its binding to the bacteria (Fig. 2D). Hence, MAb 24c5 binds M. tuberculosis GC.
M. tuberculosis GC is antigenically similar to glycogen.
MAb 24c5 bound to different sources of glycogen in ELISA, indicating antigenic similarity between M. tuberculosis GC and glycogen (Fig. 2E and F). This is not suprising as GC has structural resemblance to glycogen (21). More binding was observed with higher concentrations of glycogen, and the apparent affinity of binding to type ix glycogen was greater than the apparent affinity of binding to type viii glycogen. Differences in the reactivity of MAb 24c5 with the two forms of glycogen may reflect differences in the branching of these molecules.
MAb 24c5 GC antigen is expressed during in vitro culture.
Previous studies described M. tuberculosis GC on the surface of mycobacteria (22) or M. tuberculosis (21) without determining the quantities of surface GC for different periods of culture growth. To measure the amounts of GC antigen present on the surface of M. tuberculosis after different periods of growth, M. tuberculosis was cultured with (7H9-T medium) and without (7H9 medium) Tween 80 to compare bacteria grown as a pellicle and bacteria grown as a diffuse suspension, respectively, and cells were tested for reactivity with MAb 24c5. For this measurement, the number of bacteria used in the ELISA was standardized on the basis of the amount of protein in sedimented bacterial samples of the same volume as described previously (23, 26). MAb 24c5 bound with similar intensities to M. tuberculosis Erdman and CDC 1551 grown in 7H9 or 7H9-T medium, regardless of the period of growth (Fig. 3). M. tuberculosis CDC 1551 grown in 7H9 medium exhibited only slightly greater binding during later periods of growth than when it was grown for shorter periods of time. Binding for M. tuberculosis Erdman grown in 7H9 medium for 11 days or for CDC 1551 grown in 7H9-T medium for 11 days was only slightly less than the binding observed after growth for longer periods of time. These results indicate that a stable amount of GC antigen is associated with the M. tuberculosis cell surface during in vitro culture. Thus, the relatively constant expression of the GC antigen is different than expression of the AM antigen previously studied (26), as the amount of detectable AM antigen on M. tuberculosis Erdman increased after longer culture periods. In addition, the AM antigen was more prevalent on M. tuberculosis strain Erdman whole cells than on CDC 1551 whole cells, while similar amounts of the GC antigen were detected on the two M. tuberculosis strains (Fig. 3). These differences in AM and GC may have immunological implications, such as implications in the phagocytosis of M. tuberculosis (13, 32).
FIG. 3.

Binding of MAb 24c5 to whole cells of M. tuberculosis Erdman and CDC 1551 at various times during culture. M. tuberculosis Erdman (A and B) and CDC 1551 (C and D) were grown in the absence (A and C) or in the presence (B and D) of Tween 80 for 11, 20, or 25 days before harvest. Each data point represents the average of three measurements. Error bars show the standard deviations of the means. The growth experiments and the ELISA analysis were repeated twice, with similar results. Alk-Phos, alkaline phosphatase.
Localization of GC.
IF microscopy with MAb 24c5 revealed chorded bacterial shapes (Fig. 4A) and apparent clusters (Fig. 4B) of whole M. tuberculosis Erdman cells. Whole-mount electron microscopy of M. tuberculosis showed MAb 24c5 binding within patches of material stained by phosphotungstic acid on the bacterial surface (Fig. 5A, B, and C). Binding of irrelevant isotype-matched control MAbs did not occur in either the IF (Fig. 4C) or electron microscopy (Fig. 5D) analysis, demonstrating that there was specific binding of MAb 24c5. These findings indicate that the GC antigen recognized by MAb 24c5 is found on the surface of mycobacteria. These results are similar to those describing AM on the outermost portion of mycobacteria within a capsule-like material (22, 24, 25).
FIG. 4.
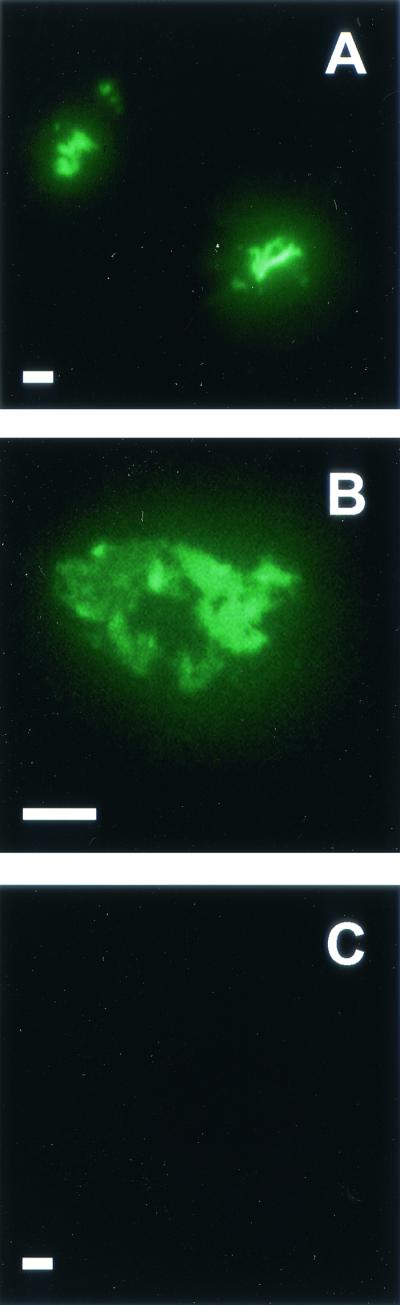
IF microscopy of M. tuberculosis Erdman using MAb 24c5. Formalin-fixed M. tuberculosis Erdman grown in the absence of Tween for 25 days was analyzed by IF microscopy using ALEXA-488-conjugated MAb 24c5 (A and B) or irrelevant control MAb (C). Bars = 5 μm. Slides were viewed at a magnification of ×100 using oil immersion. For photography an exposure time of 27 s was used for all samples. Slides were scanned into Adobe Photoshop, version 5.0, and images were transferred to Microsoft Powerpoint, version 2000, without any modification except enlargement.
FIG. 5.

Immunoelectron microscopy of M. tuberculosis using MAb 24c5. M. tuberculosis Erdman grown in the absence of Tween 80 for 25 days was analyzed for binding of MAb 24c5 (A to C) by using goat anti-mouse IgG sera conjugated to 5-nm gold. Gold appears as black dots, as indicated by arrows (A to C). The images in panels A to C are from three different bacterial fields and two different M. tuberculosis samples. Irrelevant control antibody did not allow binding of gold-conjugated sera (D). Bars = 200 nm.
These results may have important implications for M. tuberculosis pathogenesis. The interaction of pulmonary surfactant with M. tuberculosis and M. tuberculosis lipoglycan has been demonstrated to influence phagocytosis (14, 31), but the lipoglycan may not be the surface exposed element of M. tuberculosis transmitted by aerosol. Instead, the more realistic probability is that M. tuberculosis AM and GC carbohydrates are the surface exposed elements of transmissible M. tuberculosis (13). In accord with this assumption, mycobacterial carbohydrates have been implicated in the binding of these bacteria to the complement receptor CR3 (6), and M. tuberculosis GC is thought to have an important role in host cell invasion (8, 13).
GC antigens are found on the surfaces of other nontuberculosis mycobacteria.
MAb 24c5 was used to determine whether GC is found in nontuberculosis mycobacteria. The MAb 24c5 antigen was detected on the surfaces of M. kansasii, BCG Pasteur, M. smegmatis, M. phlei, M. fortuitum, and M. avium grown in vitro by ELISA (Fig. 6). This is in agreement with the work done by Lemassu et al. (22), which indicated that glucose is present in the extracellular carbohydrates of various mycobacteria. It should be noted that other growth conditions could influence the amount of MAb 24c5 binding to whole-cell mycobacteria. Our results, which are qualitative, could be very different if extended periods of growth were used, for example.
FIG. 6.

Binding of MAb 24c5 to other mycobacteria. Mycobacteria were grown in Sauton's medium without Tween 80 before harvest. Equal volumes of mycobacteria were used in ELISA. Each data point represents the average of three separate measurements. Error bars show the standard deviations of the means.
During growth, the amount of GC antigen increases in M. tuberculosis culture supernatant.
The increased amounts of polysaccharide (34) and polysaccharide antigen (26) in the culture media of M. tuberculosis grown for longer periods imply that M. tuberculosis cells shed polysaccharide antigen during prolonged culture. We therefore developed a capture ELISA to detect the presence of GC in M. tuberculosis culture supernatants (Fig. 7). The plot of absorbance versus GC concentration has a parabolic shape (revealing prozone-like effects for MAb binding to GC) with a limited slope and no conspicuous inflection point after dilution of GC in the assay mixture (Fig. 7A). As the assay tapered to background upon dilution of antigen, we were able to quantify the amounts of 24c5 antigen present in various culture supernatants relative to purified GC in the same media (Table 1). Day 11 M. tuberculosis culture supernatants did not have detectable concentrations of GC antigen (Table 1). Day 20 and day 25 M. tuberculosis culture supernatants were found to contain the GC antigen, irrespective of the presence of Tween 80 in the medium. Both M. tuberculosis Erdman and CDC 1551 accumulated this antigen (Fig. 7) at concentrations of ≥1 mg/ml after 25 days of growth (Table 1). The presence of Tween 80 diminished the amount of GC antigen found at day 25. We were unable to detect the 24c5 GC antigen in M. tuberculosis-infected BALB/c murine organ homogenates using this assay (data not shown). These results are in accord with the accumulation of AM antigen in the media of culture supernatants with or without the detergent Tween 80 (26).
FIG. 7.

GC capture ELISA and analysis of M. tuberculosis culture supernatants. The diagram shows the capture ELISA employing MAb 24c5 (Alk-Phos, alkaline phosphatase). This assay was able to detect the presence of a standard amount of M. tuberculosis (MTB) GC added to culture supernatant with or without the detergent Tween 80 (A). (B to E) M. tuberculosis Erdman (B and C) and CDC 1551 (D and E) were grown in the absence (B and D) or in the presence (C and E) of Tween 80 for 11, 20, or 25 days before the supernatant was harvested and the capture ELISA was performed. The binding to medium alone was subtracted from the values shown in panels B to E. Each data point represents the average of two measurements. Error bars show the standard deviations of the means.
TABLE 1.
GC antigen detection in M. tuberculosis culture supernatanta
| Sample | Antigen concn (μg/ml)
|
|||
|---|---|---|---|---|
| Strain | Growth medium | Day 11 | Day 20 | Day 25 |
| Erdman | 7H9 | <10 | 103 ± 1 | ≥1,000b |
| Erdman | 7H9-T | <10 | 84 ± 8 | 252 ± 4 |
| CDC 1551 | 7H9 | <10 | ≥100 | ≥1,000 |
| CDC 1551 | 7H9-T | <10 | 91 ± 9 | 1,000 ± 100 |
Detection of GC antigen in the GC capture ELISA is indicated by the mean measured concentration of antigen ± standard deviation based on two separate measurements.
In some instances, the ELISA readout exhibited a prozone but did not taper, limiting the precison of the quantitation, and the concentration is expressed as ≥100 or ≥1,000 μg/ml (the minimum concentration of antigen detected).
M. tuberculosis infection elicits an antibody response to M. tuberculosis GC.
We analyzed the murine antibody response for evidence that GC was made in vivo during experimental TB. BALB/c mice were infected with M. tuberculosis Erdman, and sera were collected before infection and on days 7, 20, and 42 after infection. Analysis of the sera indicated that there were increases in both IgG and IgM titers to GC by day 20, with augmented IgG and IgM titers on day 42 (Fig. 8). Sera collected from mice injected with heat-killed M. tuberculosis Erdman at 7, 20, or 42 days postinfection did not demonstrate IgM or IgG titers to GC greater than those found prior to inoculation (data not shown). Hence, M. tuberculosis infection in mice elicits an antibody response to GC.
FIG. 8.

Antibody titer to GC during M. tuberculosis infection. Murine serum was assayed for IgM (A) and IgG (B) antibody titers to GC at zero time and 7, 20, and 42 days after infection of mice with M. tuberculosis Erdman. Each bar represents the results for one separate mouse.
Previous studies have explored the serological activity of GC. Seibert et al. isolated polysaccharide fractions from M. tuberculosis cultures (28, 29, 30), and their polysaccharide fraction II contained an antigen that was most likely GC (7). Serological studies have shown that this fraction is immunologically active after injection into the footpads of guinea pigs (reviewed in reference 9). It was demonstrated that the 10,000-molecular-weight antigen(s), the 30,000- to 44,000-molecular-weight antigen(s), and a heterogeneous high-molecular-weight antigen from the polysaccharide II fraction were reactive with TB patient sera and control sera (5). The heterogeneous high-molecular-weight antigen was reactive with six of eight smear-positive patients and nonreactive with seven purified protein derivative skin test-positive control patients. Unfortunately, this antigen, possibly a crude GC preparation, was also reactive with three of eight purified protein derivative skin test-negative patients. As the polysaccharide II fraction contained multiple smears of unpurified sugars, it is not possible to ascertain from this data whether any specific serological reactivity with GC occurred.
Our evidence indicates that in vivo production of GC occurs during experimental M. tuberculosis infection. The production of IgG indicates that there is a maturing antibody response that usually requires continued or repeated exposure to antigen (19). The IgG antibody titer increased as the time of experimental M. tuberculosis infection increased, and we assume that this finding implies that there was continued in vivo production of GC by growing bacilli that elicited an antibody response. Since infusion of mice with heat-killed M. tuberculosis did not result in an increase in titer in comparison to uninfected mice, it is apparent that M. tuberculosis must have been growing and therefore providing GC for boosting of the antibody response. On the basis of these observations we concluded that GC antigens are made by M. tuberculosis during murine infection.
In conclusion, our results indicate that (i) M. tuberculosis GC is made in vivo, (ii) a constant amount of GC is expressed on the surface of M. tuberculosis during in vitro culture, (iii) M. tuberculosis GC is localized within the outermost surface material of the M. tuberculosis envelope, (iv) M. tuberculosis GC is antigenically similar to glycogen, and (v) similar antigens are found on other mycobacteria. This information should be considered in future studies that seek to better understand the composition of the mycobacterial cell wall or how mycobacteria interact with their host environments, as the surface carbohydrates may be involved in interactions of mycobacteria with host cells (13).
Acknowledgments
A.G.-F. has been an Aaron Diamond Young Investigator Awardee, and this work was supported in part by a grant from the Aaron Diamond Foundation. This work was also supported in part by NIH grant 1K08AI01691 (to A.G.-F.). A.C. is supported by NIH grants AI33142, AI33774, and HL59842. A.C. was also supported by a Burroughs-Wellcome Fund Scholar Award in Experimental Therapeutics. J.R.S. was supported by an NIH Training Grant in HIV, AIDS and Opportunistic Infections (grant 5T32AI07501).
We thank John T. Belisle for supplying lipoarabinomannan and MAb CS-40 as part of NIH contract N01-AI-75320. We also thank the hybridoma facility at the Albert Einstein College of Medicine for technical advice, support, and material supplied by NIH grant CA13330. We are indebted to the Howard Hughes Medical Institute at the Albert Einstein College of Medicine for use of the biohazard safety level 3 facility. For the electron microscope analysis, Frank Macaluso and Carolyn Marks assisted us at the Albert Einstein College of Medicine Analytical Imaging Facility under NIH grant CA 13330.
Editor: R. N. Moore
REFERENCES
- 1.Bloom, B. R., and C. J. Murray. 1992. Tuberculosis: commentary on a reemergent killer. Science 257:1055-1064. [DOI] [PubMed] [Google Scholar]
- 2.Brennan, P. J., and H. Nikaido. 1995. The envelope of mycobacteria. Annu. Rev. Biochem. 64:29-63. [DOI] [PubMed] [Google Scholar]
- 3.Casadevall, A., J. Mukherjee, and M. D. Scharff. 1992. Monoclonal antibody based ELISAs for cryptococcal polysaccharide. J. Immunol. Methods 154:27-35. [DOI] [PubMed] [Google Scholar]
- 4.Chatterjee, D. 1997. The mycobacterial cell wall: structure, biosynthesis and sites of drug action. Curr. Opin. Chem. Biol. 1:579-588. [DOI] [PubMed] [Google Scholar]
- 5.Coates, S. R., D. Hansen, G. Schecter, G. Slutkin, P. Hopewell, L. Affronti, and D. F. Echenberg. 1986. Identification of Mycobacterium tuberculosis antigens in Seibert fractions by immunoblotting. J. Clin. Microbiol. 24:126-130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Cywes, C., H. C. Hoppe, M. Daffe, and M. R. Ehlers. 1997. Nonopsonic binding of Mycobacterium tuberculosis to complement receptor type 3 is mediated by capsular polysaccharides and is strain dependent. Infect. Immun. 65:4258-4266. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Daffe, M., and P. Draper. 1998. The envelope layers of mycobacteria with reference to their pathogenicity. Adv. Microb. Physiol. 39:131-202. [DOI] [PubMed] [Google Scholar]
- 8.Daffe, M., and G. Etienne. 1999. The capsule of Mycobacterium tuberculosis and its implications for pathogenicity. Tuber. Lung Dis. 79:153-169. [DOI] [PubMed] [Google Scholar]
- 9.Daniel, T. M. 1984. Soluble mycobacterial antigens, p. 417-465. In G. P. Kubica and L. G. Wayne (ed.), The mycobacteria, a sourcebook, part A. Marcel Dekker, Inc., New York, N.Y.
- 10.de St. Groth, S. F., and D. Scheidegger. 1980. Production of monoclonal antibodies: strategy and tactics. J. Immunol. Methods 35:1-21. [DOI] [PubMed] [Google Scholar]
- 11.Draper, P., and R. J. Rees. 1970. Electron-transparent zone of mycobacteria may be a defense mechanism. Nature 228:860-861. [DOI] [PubMed] [Google Scholar]
- 12.Dye, C., S. Scheele, P. Dolin, V. Pathania, and R. C. Raviglione. 1999. Consensus statement. Global burden of tuberculosis—estimated incidence, prevalence, and mortality by country. W. H. O. global surveillance. JAMA 282:677-686. [DOI] [PubMed] [Google Scholar]
- 13.Ehlers, M. R., and M. Daffe. 1998. Interactions between Mycobacterium tuberculosis and host cells: are mycobacterial sugars the key? Trends Microbiol. 6:328-335. [DOI] [PubMed] [Google Scholar]
- 14.Ferguson, J. S., D. R. Voelker, F. X. McCormack, and L. S. Schlesinger. 1999. Surfactant protein D binds to Mycobacterium tuberculosis bacilli and lipoarabinomannan via carbohydrate-lectin interactions resulting in reduced phagocytosis of the bacteria by macrophages. J. Immunol. 163:312-321. [PubMed] [Google Scholar]
- 15.Glatman-Freedman, A., and A. Casadevall. 1998. Serum therapy for tuberculosis revisited: reappraisal of the role of antibody-mediated immunity against Mycobacterium tuberculosis. Clin. Microbiol. Rev. 11:514-532. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Glatman-Freedman, A., J. M. Martin, P. F. Riska, B. R. Bloom, and A. Casadevall. 1996. Monoclonal antibodies to surface antigens of Mycobacterium tuberculosis and their use in a modified enzyme-linked immunosorbent spot assay for detection of mycobacteria. J. Clin. Microbiol. 34:2795-2802. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Glatman-Freedman, A., A. J. Mednick, N. Lendvai, and A. Casadevall. 2000. Clearance and organ distribution of Mycobacterium tuberculosis lipoarabinomannan (LAM) in the presence and absence of LAM-binding immunoglobulin M. Infect. Immun. 68:335-341. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Imaeda, T., F. Kanetsuna, and B. Galindo. 1968. Ultrastructure of cell walls of genus Mycobacterium. J. Ultrastruct. Res. 25:46-63. [DOI] [PubMed] [Google Scholar]
- 19.Janeway, C. A., and P. Travers. 1997. The humoral immune response, p. 8:1-8:54. In Immunobiology: the immune system in health and disease, 3rd ed. Garland Publishing Inc., New York, N.Y.
- 20.Konadu, E., J. Shiloach, D. A. Bryla, J. B. Robbins, and S. C. Szu. 1996. Synthesis, characterization, and immunological properties in mice of conjugates composed of detoxified lipopolysaccharide of Salmonella paratyphi A bound to tetanus toxoid with emphasis on the role of O acetyls. Infect. Immun. 64:2709-2715. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Lemassu, A., and M. Daffe. 1994. Structural features of the exocellular polysaccharides of Mycobacterium tuberculosis. Biochem. J. 297:351-357. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Lemassu, A., A. Ortalo-Magne, F. Bardou, G. Silve, M.-A. Lanéelle, and M. Daffe. 1996. Extracellular and surface-exposed polysaccharides of non-tuberculosis mycobacteria. Microbiology 142:1513-1520. [DOI] [PubMed] [Google Scholar]
- 23.Meyers, P. R., W. R. Bourn, L. M. Steyn, P. VanHelden, A. D. Beyers, and G. D. Brown. 1998. Novel method for rapid measurement of growth of mycobacteria in detergent-free media. J. Clin. Microbiol. 36:2752-2754. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Ortalo-Magne, A., A. B. Andersen, and M. Daffe. 1996. The outermost capsular arabinomannans and other mannoconjugates of virulent and avirulent tubercle bacilli. Microbiology 142:927-935. [DOI] [PubMed] [Google Scholar]
- 25.Ortalo-Magne, A., M.-A. Dupont, A. Lemassu, A. B. Andersen, P. Gounon, and M. Daffe. 1995. Molecular composition of the outermost capsular material of the tubercle bacillus. Microbiology 141:1609-1620. [DOI] [PubMed] [Google Scholar]
- 26.Schwebach, J. R., A. Casadevall, R. Schneerson, Z. Dai, X. Wang, J. B. Robbins, and A. Glatman-Freedman. 2001. Expression of a Mycobacterium tuberculosis arabinomannan antigen in vitro and in vivo. Infect. Immun. 69:5671-5678. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Schwebach, J. R., W. R. Jacobs, and A. Casadevall. 2001. Sterilization of Mycobacterium tuberculosis Erdman samples by antimicrobial fixation in a biosafety level 3 laboratory. J. Clin. Microbiol. 39:769-771. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Seibert, F. B. 1949. The isolation of three different proteins and two polysaccharides from tuberculin by alcohol fractionation. Their chemical and biological properties. Am. Rev. Tuberc. 59:86-101. [DOI] [PubMed] [Google Scholar]
- 29.Seibert, F. B., K. O. Pedersen, and A. Tiselius. 1938. Molecular weight, electrochemical and biological properties of tuberculin protein and polysaccharide molecules. Am. Rev. Tuberc. 38:399-405. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Seibert, F. B., and D. W. Watson. 1941. Isolation of the polysaccharides and nucleic acid of tuberculin by electrophoresis. J. Biol. Chem. 140:55-69. [Google Scholar]
- 31.Sidobre, S., J. Nigou, G. Puzo, and M. Riviere. 2000. Lipoglycans are putative ligands for the human pulmonary surfactant protein A attachment to mycobacteria. Critical role of the lipids for lectin-carbohydrate recognition. J. Biol. Chem. 275:2415-2422. [DOI] [PubMed] [Google Scholar]
- 32.Stokes, R. W., and D. Doxsee. 1999. The receptor-mediated uptake, survival, replication, and drug sensitivity of Mycobacterium tuberculosis within the macrophage-like cell line THP-1: a comparison with human monocyte-derived macrophages. Cell. Immunol. 197:1-9. [DOI] [PubMed] [Google Scholar]
- 33.Teitelbaum, R., A. Glatman-Freedman, B. Chen, J. Robbins, E. Unanue, A. Casadevall, and B. R. Bloom. 1998. A monoclonal antibody recognizing a surface antigen of Mycobacterium tuberculosis enhances host survival. Proc. Natl. Acad. Sci. USA 95:15688-15693. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Turcotte, R., and Y. Des Ormeaux. 1972. Influence of the age of mycobacterial cultures on the protein and carbohydrate composition of tuberculins. Can. J. Microbiol. 18:637-645. [DOI] [PubMed] [Google Scholar]