

Abstract
Pathogen DNA was isolated from roan antelope (Hippotragus equinus), sable antelope (Hippotragus niger), greater kudu (Tragelaphus strepsiceros), and common gray duiker (Sylvicapra grimmia) in South Africa whose deaths were attributed to either theileriosis or cytauxzoonosis. We developed Theileria species-specific probes used in combination with reverse line blot hybridization assays and identified three different species of Theileria in four African antelope species. The close phylogenetic relationship between members of the genera Theileria and Cytauxzoon, similarities in the morphologies of developmental stages, and confusion in the literature regarding theileriosis or cytauxzoonosis are discussed.
The populations of Hippotragus species, comprising sable (Hippotragus niger) and roan (Hippotragus equinus) antelopes, have shown alarming decreases in numbers in the past decades. In the Kruger National Park, South Africa, for instance, the number of roan antelopes declined from 450 free-ranging animals in 1986 to 30 in 2002. Both sable and roan antelopes are listed on the World Conservation Union Red List as taxa whose continued survival is dependent on active conservation measures (7). Despite good breeding results in South Africa, one of the main reasons for the population decline is an abnormally high rate of calf mortality, which has been attributed to theileriosis, alternatively reported as cytauxzoonosis (23, 26, 27).
Other tick-borne parasitoses of African antelope associated with disease that have been reported in the past include theileriosis (cytauxzoonosis) in greater kudu (Tragelaphus strepsiceros) (17), gray duiker (Sylvicapra grimmia) (18), tsessebe (Damaliscus lunatus) (8), and eland (Taurotragus oryx) (5, 6). Moreover, babesiosis has been reported in roan antelopes (27) and sable antelopes (13, 15, 24), and anaplasmosis has been reported in sable antelopes (4, 24). We report here on fatal cases of theileriosis in gray duiker, translocated roan and sable antelopes, and greater kudu caused by three different Theileria spp. A reverse line blot hybridization (RLB) assay for molecular detection of these parasites is described. The consequences of translocation for each of these species with respect to contracting theileriosis are briefly discussed. Finally, the taxonomic status of Theileria species in wildlife is discussed.
CASE REPORTS
Roan antelope (Hippotragus equinus) originating from Togo and Benin in West Africa contracted theileriosis after release following translocation on a private game farm in the Lowveld of South Africa near Malelane, in southern Mpumalanga. The clinical signs observed were depression, characterized by drooping of the ears, prolonged intermittent recumbency, anorexia, and lagging behind the herd. On inspection after immobilization, the animals were found to be anemic, icteric, and pyrexic, with generalized lymph node enlargement and prolonged bleeding times. Blood smears revealed lymphoblasts parasitized by typical theilerial schizonts, as shown in Fig. 1, and numerous piroplasms infecting erythrocytes. All treatment attempts failed, and the animals were found dead within 2 days after the onset of clinical signs. Postmortem findings were icterus and widespread petechiae (especially in the renal cortex), splenomegaly and red pulp hyperplasia, pulmonary edema, and a generalized lymphoid hyperplasia. Histopathological lesions were consistent with theileriosis as the primary cause of death. A further description of the pathology will be reported elsewhere. Blood samples from two fatal cases were available for molecular characterization of the parasites.
FIG. 1.
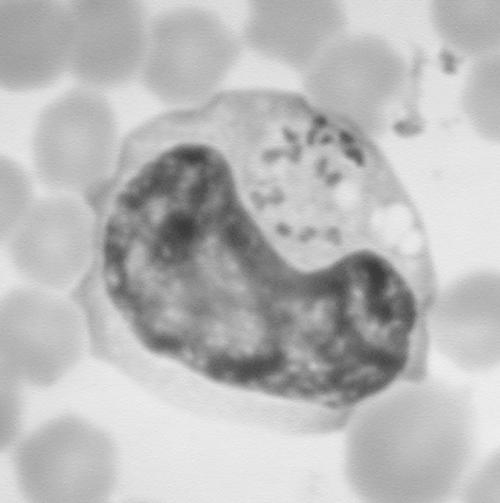
Blood smear photograph showing Theileria sp. (sable) schizonts in lymphoblastoid cells. Magnification, ×250.
A sable antelope (Hippotragus niger) originating from a game ranch in the Pretoria area, Gauteng Province, South Africa, showed clinical signs and postmortem findings similar to those observed in the roan antelope. Clinical signs were characterized by pronounced anemia and icterus. Postmortem findings included petechiae and ecchymoses in the subcutis, lymph nodes, peritoneum, pleura, gallbladder, endo- and epicardia, lungs, and kidneys, as well as cerebral and pulmonary edema. Red pulp hyperplasia of the spleen and hepatosis with cholestasis were marked. A blood sample from this case was available for further characterization.
Soon after arrival, greater kudu (Tragelaphus strepsiceros) translocated from the Eastern Cape Province of South Africa to a game ranch near Mossel Bay, Western Cape Province, developed clinical signs which resembled those described previously for theileriosis in this species (17). Several animals died of the disease, but no postmortem examination was performed. EDTA-anticoagulated blood from one of the animals that died was sent to us for molecular characterization.
A gray duiker (Sylvicapra grimmia) died from theileriosis on a private game farm in Gauteng Province, South Africa. The animal was fully mature and was found dead before any clinical signs could be detected. The macroscopic postmortem lesions found were mild anemia, reasonable to good body condition, peripheral lymph node enlargement (hyperplasia), scattered petechiae in the renal cortex and visceral serosal surfaces, marked splenomegaly resulting from red pulp hyperplasia, and edematous lungs. Microscopic findings on blood smears revealed numerous piroplasms in erythrocytes and schizont-infected lymphocytes and monocytes. Histopathology showed Theileria schizonts in lympho- and monoblast-infiltrated parenchymatous organs. A spleen sample was available for further characterization.
MATERIALS AND METHODS
DNA extraction.
DNA was extracted from EDTA-anticoagulated blood or tissue stored at −20°C with a DNA extraction kit (QIAgen, Southern Cross Biotechnology, Cape Town, South Africa), following the manufacturer's protocols.
PCR amplification, cloning, and sequencing.
PCR products used for the detection of Theileria and Babesia species by RLB were obtained as described previously with primers RLB-F2 (5′-GAC ACA GGG AGG TAG TGA CAA G-3′) and RLB-R2 (biotin-5′-CTA AGA ATT TCA CCT CTG ACA GT-3′) (19). PCR products for the detection of Anaplasma and Ehrlichia species by RLB were amplified as described by Bekker et al. (2), with modifications in the PCR conditions similar to those described for the Theileria and Babesia PCR.
DNA amplicons were cloned into the pGem-T Easy vector (Promega, Leiden, The Netherlands), according to the manufacturer's instructions. Positive clones were selected and PCR amplified with biotinylated M13 primers for subsequent sequencing and screening by RLB to detect the presence of possible paralogue sequences.
For sequencing purposes, the 18S rRNA genes of the parasites identified in this study as Theileria sp. (sable) from sable antelope, Theileria sp. (greater kudu), and Theileria sp. (gray duiker) were PCR amplified. Sequencing was carried out in two parts with primers 18SFTB (5′-GGA TAA CCG TGC TAA TTG TAG G-3′) and RLB-R2 for the first approximately 720 bp and primers RLB-F2 and 18SRTB (5′-GAA TAA TTC ACC GGA TCA CTC G-3′) for the second part, which spanned approximately 1,180 bp and which overlapped the first part by approximately 400 bp. The PCR mixture contained the following in a 50-μl volume: 25 μl of Platinum Quantitative PCR Supermix-UDG (Invitrogen, Laboratory Specialist Services, Clareinch, South Africa), 50 pmol of each primer (Isogen Life Science, Maarssen, The Netherlands), and 5 μl of template DNA.
Amplification was performed in an automated thermocycler (Perkin-Elmer, Foster City, CA). Samples were first heated at 42°C for 3 min to activate the uracil DNA glycosylase (UDG), thereby degrading previous PCR amplicon contamination possibly present, followed by heating for 10 min at 94°C to inactivate the UDG. The second phase consisted of 35 cycles of 45 s at 92°C, 45 s at 57°C, and 45 s at 72°C. Eventually, the samples were heated at 72°C for 7 min and thereafter stored at 4°C until they were sent for sequencing. The PCR product from which the partial sequences of the parasite identified in this study as a Theileria sp. (sable) isolated from roan antelope was prepared by using the PCR procedure used for RLB; and all samples were sent to Inqaba Biotec, Pretoria, South Africa, for sequencing.
Reverse line blot hybridization.
Although RLB has previously been used to identify blood parasites in African wildlife, a further description of the methods is required here due to many changes in comparison with the methods used in the study described in our previous report (19).
Oligonucleotide probes containing an N-terminal N-(trifluoracetamidohexyl-cyanoethyl,N,N-diisopropyl phosphoramidite)-C6 amino linker (Isogen Life Science, Maarssen, The Netherlands) were covalently linked to the RLB membrane (Biodyne C blotting membrane; Pall Biosupport, Ann Arbor, MI) by using the following procedure. The membrane was activated by a 10-min incubation at room temperature in a freshly prepared 10-ml solution of 16% 1-ethyl 3-(3-dimethylaminopropyl)carbodiimide HCl (Sigma, St. Louis, MO). After the membrane was rinsed in distilled water, it was placed in an MN45 miniblotter apparatus (Immunetics, Cambridge, MA). The oligonucleotide probes were diluted to a concentration of 100 to 800 pmol/150 μl in 500 mM NaHCO3 solution (pH 8.4) and linked to the membrane by loading them onto the lanes of the miniblotter, followed by a 1-min incubation. After aspiration of the oligonucleotide probe solutions, the membrane was inactivated by 8 min incubation in a 100 ml 100 mM NaOH solution at room temperature. A washing step for 5 min at 60°C in a 2× SSPE (2× SSPE is 0.36 M NaCl, 20 mM NaH2PO4, and 2 mM EDTA [pH 7.7])-0.1% sodium dodecyl sulfate (SDS) solution took place before the membrane was stored before use in a 20 mM EDTA solution after a 15-min rinsing period in 20 mM EDTA at room temperature.
Hybridization of the RLB membrane with the PCR products was as follows: 10 μl of PCR product was diluted in 140 μl 2× SSPE-0.1% SDS, denatured for 10 min at 100°C, and subsequently chilled on ice to prevent annealing of the denatured strands. In the meantime, the RLB membrane was rinsed for 5 min at room temperature in a 2× SSPE-0.1% SDS solution. After a short centrifugation of the samples, they were loaded onto the RLB membrane by using the MN45 miniblotter apparatus and incubated at 42°C for 1 h. Thereafter, the PCR products were aspirated and the membrane was washed twice at 50°C in a 2× SSPE-0.5% SDS solution, followed by incubation with 10 ml 1:4,000-diluted peroxidase-labeled streptavidin (Boehringer Mannheim, Mannheim, Germany) in 2× SSPE-0.5% SDS at 42°C for 30 min. The membrane was washed twice again with 2× SSPE-0.5% SDS, but this time at 42°C for 10 min, and afterwards was washed twice for 5 min each time in a 2× SSPE solution at room temperature. The membrane was incubated in 10 ml of enhanced chemiluminescence (ECL) detection fluid (Amersham Biosciences, United Kingdom) before exposure to an ECL hyperfilm (Amersham Biosciences) and development. The PCR products were stripped from the membrane by two washes for 30 min each time at 90°C in a 1% SDS solution. Rinsing of the membrane in 20 mM EDTA solution (pH 8.4) for 15 min followed, before storage of the membrane in fresh EDTA at 4°C.
Phylogenetic analyses.
For phylogenetic analysis, the ClustalX program (version 1.81 for Windows) was used to align the 18S rRNA sequences of selected pathogens. This selection was based on their relative importance in veterinary medicine; their occurrence in related African wildlife; or a high level of 18S rRNA sequence similarity with the three described Theileria spp., as found by a BLAST search of the National Center for Biotechnology Information database.
The alignment was manually edited in BioEdit (version 5.0.9) and truncated to the size of the smallest sequence (Theileria. sp. [sable]; 1,400 bp). Several phylogenetic trees were created by using the Treecon package (version 1.3b) and the Phylip package (Felsenstein, version 3.63). Different approaches were evaluated in order to identify clades supported by the highest possible bootstrap values.
Nucleotide sequence accession numbers.
The 18S rRNA gene sequences of the species designated Theileria sp. (sable), Theileria sp. (greater kudu), and Theileria sp. (gray duiker) have been submitted to GenBank and can be retrieved under accession numbers AY748462, AY748465, and AY748466, respectively. The 18S rRNA gene sequences of the Theileria spp. (sable) isolated from the two fatal roan cases have been given accession numbers AY748463 and AY748464.
RESULTS
When the PCR products amplified from sable antelope, greater kudu, and common gray duiker DNA were subjected to reverse line blotting, a Theileria or Babesia catch-all signal without a species-specific signal was detected, indicating the presence of a novel species or a variant of a species. A BLAST search performed with the 18S rRNA gene sequences from these samples revealed a 99.6% identity between the parasite isolated from sable antelope and Theileria sp. (sable) (GenBank accession number L19081) (Fig. 2). The 18S rRNA sequence of the parasite isolated from the common gray duiker showed the highest similarity with Theileria separata (GenBank accession number AY260175) (Fig. 2). The parasite derived from greater kudu showed the most similarities with Theileria sp. (Thung Song) (GenBank accession number AB000270), a parasite isolated from Bos indicus in Thung Song District, Thailand (3). Phylogenetic analysis confirmed these similarities and clustering of these species within the Theileria branch by all methods used. The resulting tree obtained by the neighbor-joining method with the Kimura two-parameter distance calculation is shown in Fig. 2.
FIG. 2.

Neighbor-joining tree showing the phylogenetic relationships of Theileria sp. (sable), Theileria sp. (gray duiker), and Theileria sp. (greater kudu) with other Babesia and Theileria spp. based on full-length 18S rRNA sequences. The numbers represent the percentage of 1,000 replicants (bootstrap support) for which the same branching patterns were obtained. Toxoplasma gondii was used as an outgroup.
Species-specific RLB oligonucleotide probes were deduced from the hypervariable V4 region of the 18S rRNA gene sequence and synthesized by Isogen Life Science, Maarssen, The Netherlands (Table 1). These probes were shown to bind only to their respective target sequences (results not shown). The two samples from the roan antelope reached us at a later stage during our investigation. Initially, both samples gave a positive signal on RLB only with the Theileria sp. (sable)-specific probe, indicating the strong similarity with this parasite. This was later confirmed by aligning the (partial) 18S rRNA sequences from the roan antelope parasites to Theileria sp. (sable), as they were shown to be identical. Positive clones derived from each sample (eight from the sable antelope, seven from the gray duiker, and two from the greater kudu) were shown to bind only to the Theileria sp. (sable), Theileria sp. (duiker), and Theileria sp. (kudu) species-specific RLB probes, respectively.
TABLE 1.
New species-specific RLB probes used in this study
| Oligonucleotide probe specificity | 18S rRNA sequence (5′→3′) | GenBank accession no(s). | Tm (°C)a |
|---|---|---|---|
| Theileria sp. (greater kudu) | CTG CAT TGT TTC TTT CCT TTG | AY748465 | 54.0 |
| Theileria sp. (grey duiker) | CAT TTT GGT TAT TGC ATT GTG G | AY748466 | 56.3 |
| Theileria spp. (sable) | GCT GCA TTG CCT TTT CTC C | L19081, AY748462 to AY748464 | 56.7 |
The melting point (Tm) was determined empirically.
Theileria sp. (sable) was also detected by RLB in a number of samples from other healthy animals of the family Bovidae, such as the African buffalo (Syncerus caffer), klipspringer (Oreotragus oreotragus), reedbuck (Redunca arundinum), blue wildebeest (Connonhaetes taurinus), blesbok (Damaliscus pygargus), and cattle (Bos indicus) from various regions in southern Africa and Tanzania, indicating a wider distribution of this parasite (Table 2). This finding is in accordance with recent findings reported by Spitalska et al. (22), whereby a Theileria sp. with an 18S rRNA sequence similar to that of Theileria sp. (sable) was isolated from red hartebeest (Alcelaphus buselaphus caama) in Namibia.
TABLE 2.
Clinically healthy animals other than roan or sable antelopes whose blood samples were examined by RLB and found to be positive for Theileria sp. (sable) and their origins
| Animal species | Origin |
|---|---|
| African buffalo (Syncerus caffer) | Carthage, South Africa |
| African short-horn cattle (Bos indicus) | Arusha, Tanzania |
| Blesbok (Damaliscus pygargus) | Swaziland |
| Blue wildebeest (Connochaetes taurinus) | Malelane, South Africa |
| Klipspringer (Oreotragus oreotragus) | Rust de Winter, South Africa |
| Reedbuck (Redunca arundinum) | Mankdare, South Africa |
Finally, samples from the animals described in this case report were negative for Anaplasma or Ehrlichia species, based on RLB.
DISCUSSION
Translocation of wildlife is usually performed for conservation of an endangered species or to restock areas for hunting and other commercial purposes. However, such animals may introduce diseases into their new environment; or the introduction of naïve animals into an area of endemicity may have severe consequences, as demonstrated in this study by the fatal cases of disease in roan antelope, sable antelope, and kudu. Furthermore, disease due to a latent theileriosis infection may also be triggered by stress factors resulting from translocation. Health risk assessment of wildlife translocations is greatly facilitated by identification of the possible health hazards (10). In this respect, the application of molecular diagnostics to wildlife diseases (in particular, reverse line blotting) has been shown to be very useful, for instance, in identifying blood parasites associated with mortality in the black rhinoceros (19).
Despite good breeding results for the roan and sable antelopes distributed over several South African game parks, high rates of calf mortality have been observed and have been attributed to theileriosis (26, 27). A possible factor that may contribute to this situation could be related to the confinement of these animals in relatively small wildlife reserves, which leads to high stocking rates that will support the buildup of large tick populations (20). This would lead to a heavy challenge of naïve calves by Theileria-infected ticks. The finding of Theileria sp. (sable) in a wide range of Bovidae other than roan and sable antelope (Table 2) suggests that it is likely that other species form a natural reservoir of this parasite, possibly limiting successful coexistence with susceptible roan and sable antelopes in areas where the vector is present.
The first description of Theileria-like piroplasms in antelopes of the genus Hippotragus was published in 1912. The parasite was observed in erythrocytes of a roan antelope shot in the Gambia, West Africa, and was given the name Theileria hippotragi (25). In South Africa, Theileria parasites were observed in blood smears from sable antelope by Neitz (17). However, in 1948, Neitz and Thomas (18) proposed the name Cytauxzoon (Greek for an increase in the number of cells in an animal) for a new genus to accommodate those Theileria-like parasites with preerythrocytic schizogony in histiocytes, as opposed to schizogony in lymphocytes, with the latter being characteristic for Theileria. Furthermore, in 1974, Wilson and colleagues described a Theileria-like species associated with mortality in juvenile sable and roan antelopes in game reserves in Limpopo Province, South Africa, as Cytauxzoon (26, 27).
Several Cytauxzoon species have been described in African antelope based on these criteria: Cytauxzoon sylvicaprae, which originated from the common gray duiker (18), and Cytauxzoon strepsicerosi in the greater kudu (17). Moreover, mortality due to cytauxzoonosis in South Africa has been reported in tsessebe (8) and giraffe (Giraffa camelopardalis) (14), but specific names were not given. Importantly, a blood parasite infecting eland antelope initially described as Cytauxzoon taurotragi (12) was later shown to be infective to both lymphoid and macrophage-like cell lines and thus behaved as a typical Theileria species. In 1980, Grootenhuis et al. (5) renamed the parasite as Theileria taurotragi and suggested that more work would be required to show the validity of the genus Cytauxzoon. Subsequently, Thomas et al. (1982) suggested synonymization of the genus Cytauxzoon with Theileria based on observations of blood parasites in sable antelope (24). Interestingly, Stoltsz and Dunsterville managed to establish and cultivate a Theileria species in lymphoblastoid cells from a sable antelope that died of theileriosis (23). The 18S rRNA of this species was sequenced (GenBank accession number L19081) and was provisionally named Theileria sp. (sable) based on phylogenetic analysis (1). This sequence is identical to the sequences that we obtained from the fatal roan and sable antelope cases reported in this paper.
It is tempting to suggest that the fatal cases of cytauxzoonosis described in roan and sable antelopes in the past were in fact caused by this Theileria sp. (sable), also because of the strong similarities in clinical signs and pathology. Unfortunately, a search for the original 1974 samples at the Pathology Section of the Faculty of Veterinary Science, University of Pretoria, was fruitless, as none of the samples were found, making molecular characterization and a subsequent comparison thereof with Theileria sp. (sable) impossible.
The same appears to be the case for the parasites from greater kudu and gray duiker, since phylogenetic analysis of the sequenced 18S rRNA shows that they cluster within the Theileria group, as shown by parsimony and neighbor-joining methods (Fig. 2). Since we were unsuccessful in our extensive search for the type specimens of Cytauxzoon sylvicaprae and Cytauxzoon strepsicerosi for an 18S rRNA sequence comparison as well, it remains unclear whether we have in our hands the same species as the described Cytauxzoon species.
If the three Theileria spp. described here were the same parasites that have been referred to as Cytauxzoon spp. in the past, it would leave the genus Cytauxzoon with only a limited number of species of wild and domestic members of the family Felidae, such as Cytauxzoon felis (9). In our phylogenetic analysis, Cytauxzoon felis forms a separate cluster with Theileria equi (Fig. 2), which was previously referred to as Babesia equi and which was reclassified based on the presence of typical characteristics of Theileria, such as intralymphocytic schizonts and phylogenetic evidence (1, 16). Moreover, Kocan et al. redescribed the schizogony in Cytauxzoon felis and found it to be analogous to merozoite formation of Theileria (9). Interestingly, according to those authors, the only difference with Theileria was the observation of schizogony in macrophages instead of lymphocytes. However, development in macrophages is typical for Theileria and has been well documented, for instance, for Theileria annulata (21). This indicates that Cytauxzoon felis may be regarded as a Theileria species. On the other hand, C. felis and Cytauxzoon species from other wild Felidae form a separate cluster from Theileria spp. (Fig. 2).
In conclusion, based on the close phylogenetic relationship between members of the genera Theileria and Cytauxzoon and very similar developmental cycles, we suggest that the taxonomic status of etiological agents of theileriosis in African antelope species be clarified by linking Theileria hippotragi to theileriosis in roan and sable antelopes. Finally, although we would like to follow the suggestion of Levine made in 1971 that Theileria sylvicaprae and Theileria strepsicerosi should replace Cytauxzoon sylvicaprae and Cytauxzoon strepsicerosi, respectively (11), and be used to refer to theileriosis in common gray duiker and greater kudu, respectively, we have not been able to provide sufficient evidence for this because of a lack of type specimens.
Acknowledgments
We thank Gerhard Olivier for information on the fatal cases of theileriosis in greater kudu.
The research reported here was conducted within the framework of the Memorandum of Understanding between the Faculty of Veterinary Science of the University of Pretoria and the Faculty of Veterinary Medicine of Utrecht University, Utrecht, The Netherlands. This study was supported in part by Senter, Ministry of Economic Affairs, The Netherlands, through project SA 010111, entitled “Development of a biochip for the molecular diagnosis of pathogens in wildlife in South Africa.”
REFERENCES
- 1.Allsopp, M. T. E. P., T. Cavalier-Smith, D. T. de Waal, and B. A. Allsopp. 1994. Phylogeny and evolution of the piroplasms. Parasitology 108:147-152. [DOI] [PubMed] [Google Scholar]
- 2.Bekker, C. P. J., S. de Vos, A. Taoufik, O. A. E. Sparagano, and F. Jongejan. 2002. Simultaneous detection of Anaplasma and Ehrlichia species in ruminants and detection of Ehrlichia ruminantium in Amblyomma variegatum ticks by reverse line blot hybridization. Vet. Microbiol. 89:223-238. [DOI] [PubMed] [Google Scholar]
- 3.Chansiri, K., and N. Sarataphan. 2002. Molecular phylogenetic study of Theileria sp. (Thung Song) based on the thymidylate synthetase gene. Parasitol. Res. 88:S33-S35. [DOI] [PubMed] [Google Scholar]
- 4.Grobler, J. H. 1981. Parasites and mortality of sable antelope Hippotragus niger niger (Harris, 1838) in the Matopos, Zimbabwe. Koedoe 24:119-123. [Google Scholar]
- 5.Grootenhuis, J. G., A. S. Young, T. T. Dolan, and P. A. Stagg. 1979. Characteristics of Theileria species (eland) infection in eland and cattle. Res. Vet. Sci. 27:59-68. [PubMed] [Google Scholar]
- 6.Grootenhuis, J. G., W. I. Morrison, L. Karlstad, P.D. Sayer, A.S. Young, M. Murray, and R. D. Haller. 1980. Fatal theileriosis in eland (Taurotragus oryx), pathology of natural and experimental cases. Res. Vet. Sci. 29:219-229. [PubMed] [Google Scholar]
- 7.Hilton-Taylor, C. (compiler). 2000. 2000 IUCN red list of threatened species. World Conservation Union/Species Survival Commission, Gland, Switzerland, and Cambridge, United Kingdom.
- 8.Jardine, J. J. 1992. The pathology of cytauxzoonosis in a tsessebe (Damaliscus lunatus). J. S. Afr. Vet. Assoc. 63:49-51. [PubMed] [Google Scholar]
- 9.Kocan, A. A., K. M. Kocan, E. F. Blouin, and S. W. Mukolwe. 1992. A redescription of schizogony of Cytauxzoon felis in the domestic cat. Ann. N. Y. Acad. Sci. 653:161-167. [DOI] [PubMed] [Google Scholar]
- 10.Leighton, F. A. 2002. Health risk assessment of the translocation of wild animals. Rev. Sci. Tech. Off. Int. Epizoot. 21:187-195. [PubMed] [Google Scholar]
- 11.Levine, N. D. 1971. Taxonomy of the piroplasms. Trans. Am. Microsc. Soc. 90:2-33. [Google Scholar]
- 12.Martin, H., and D. W. Brocklesby. 1960. A new parasite of eland. Vet. Rec. 72:331-332. [Google Scholar]
- 13.Martinaglia, G. 1930. Red-water (babesiosis) in a sable antelope. J. S. Afr. Vet. Med. Assoc. 1:41-42. [Google Scholar]
- 14.McCully, R. M., M. E. Keep, and P. A. Basson. 1970. Cytauxzoonosis in a giraffe [Giraffa camelopardalis (Linnaeus, 1758)] in Zululand. Onderstepoort J. Vet. Res. 37:7-10. [PubMed] [Google Scholar]
- 15.McInnes, E. F., C. G. Stewart, B. L. Penzhorn, and D. G. A. Meltzer. 1991. An outbreak of babesiosis in imported sable antelope (Hippotragus niger). J. S. Afr. Vet. Assoc. 62:30-32. [PubMed] [Google Scholar]
- 16.Mehlhorn, H., and E. Schein. 1998. Redescription of Babesia equi Laveran, 1901 as Theileria equi Mehlhorn, Schein 1998. Parasitol. Res. 84:467-475. [DOI] [PubMed] [Google Scholar]
- 17.Neitz, W. O. 1957. Theileriosis, gonderioses and cytauxzoonoses. Onderstepoort J. Vet. Res. 27:275-430. [Google Scholar]
- 18.Neitz, W. O., and A. D. Thomas. 1948. Cytauxzoon sylvicaprae gen. nov., spec. nov., a protozoon responsible for a hitherto undescribed disease in the duiker [Sylvicapra grimmia (Linné)]. Onderstepoort J. Vet. Sci. Anim. Ind. 23:63-76. [PubMed] [Google Scholar]
- 19.Nijhof, A. M., B. L. Penzhorn, G. Lynen, J. O. Mollel, P. Morkel, C. P. J. Bekker, and F. Jongejan. 2003. Babesia bicornis sp. nov. and Theileria bicornis sp. nov.: tick-borne parasites associated with mortality in the black rhinoceros. J. Clin. Microbiol. 41:2249-2254. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Norval, R. A. I. 1977. Tick problems in relation to land utilization in Rhodesia. Rhod. Vet. J. 8:33-38. [Google Scholar]
- 21.Preston, P. M., F. R. Hall, E. J. Glass, J. D. Campbell, M. A. Darghouth, J. S. Ahmed, B. R. Shiels, R. L. Spooner, F. Jongejan, and C. G. Brown. 1999. Innate and adaptive immune responses co-operate to protect cattle against Theileria annulata. Parasitol. Today 15:268-274. [DOI] [PubMed] [Google Scholar]
- 22.Spitalska, E., M. Riddell, H. Heyne, and O. A. Sparagano. 2005. Prevalence of theileriosis in Red Hartebeest (Alcelaphus buselaphus caama) in Namibia. Parasitol. Res. 97:77-79. [DOI] [PubMed] [Google Scholar]
- 23.Stoltsz, W. H., and M. T. Dunsterville. 1992. In vitro establishment and cultivation of a Cytauxzoon sp. (Theileria sp.) from a sable antelope (Hippotragus niger, Harris 1838). J. S. Afr. Vet. Ass. 63:182. (Abstract.) [Google Scholar]
- 24.Thomas, S. E., D. E. Wilson, and T. E. Mason. 1982. Babesia, Theileria and Anaplasma spp. infecting sable antelope, Hippotragus niger (Harris, 1838), in southern Africa. Onderstepoort J. Vet. Res. 49:163-166. [PubMed] [Google Scholar]
- 25.Todd, J. L., and S. B. Wolbach. 1912. Parasitic protozoa from the Gambia. J. Med. Res. 26:195-218. [PMC free article] [PubMed] [Google Scholar]
- 26.Wilson, D. E., and S. M. Hirst. 1977. Ecology and factors limiting roan and sable antelope populations in South Africa. Wildlife Monograph 54. Wildlife Society, Washington, D.C.
- 27.Wilson, D. E., R. C. Bartsch, R. D. Bigalke, and S. E. Thomas. 1974. Observations on mortality rates and disease in roan and sable antelope on nature reserves in the Transvaal. J. S. Afr. Wildl. Mgmt. Assoc. 4:203-206. [Google Scholar]