

Abstract
A report on the Ninth International Gap Junction Conference, Honolulu, USA, 4-9 August 2001.
Researchers working in the field of gap junctions, the membrane structures dedicated to direct cell-cell communication, might have felt, at times, as if they are children of a lesser god. The molecular cloning of connexins and their identification as the protein components of gap junctions heralded a new era, but compared to the progress achieved in deciphering other junctional structures, it looked as though cadherin and integrins were sprinting in a fast lane and leaving gap junctions behind. But gap junctions are not simple structures of contact between cells, they are in fact channels formed by the oligomerization of connexins: six connexins make a connexon or hemichannel, and two connexons from adjacent cells align in the extracellular space to make a full intercellular gap junction channel that allows direct communication between cells without using the extracellular space to exchange messages. Compared to classical ion channels, however, studies of intercellular ones have suffered from the lack of clear ionic selectivity and of specific toxins to probe their function. But if these aspects led to a grim mood among researchers, it has disappeared with the discovery that several genetic diseases are linked to mutations in connexin genes. The recent meeting on connexins and gap junctions, attended by the vast majority of people active in the field, confirmed some of the trends that have emerged and that are allowing us to make sense of the complexity of connexin language, and announced some of the new avenues that are beginning to be explored.
Connexin family members and nomenclature
With the completion of the human genome sequence, it appears that we have at least 20 connexins (Figure 1). Family members are usually distinguished by their expected molecular weight in kilodaltons, so that the best-known connexin (Cx), a protein of 43 kDa, is referred to as Cx43. It turns out that nature has produced a number of connexin proteins in the 29-32 kDa range, however, forcing the use of decimal figures to distinguish these family members. The main drawback of this nomenclature is that the corresponding orthologs in other species cannot be called by the same name. For example, human Cx30.2 (HCx30.2) has a shorter counterpart in the mouse, mCx29, whereas mCx30.2 (yes, there is one!) has its human counterpart in HCx31.9. To sort things out, Klaus Willecke (University of Bonn, Germany) and Dan Goodenough (Harvard Medical School, Boston, USA) proposed a new nomenclature in which connexins would be first ordered according to their molecular weight (from lighter to heavier) and then distinguished by progressive Arabic numbers, similar to the nomenclature adopted for keratins, fibroblast growth factors and other gene families. Human connexins would be taken as templates, as we have more complete information than for the mouse genome, so that Cx25 would become Cx1 and Cx62 would be renamed Cx20. In the case of genes with no counterparts in the human genome (for example, rat and mouse Cx33), the old nomenclature would be maintained. Although a majority of people participating in this discussion favored the change, some objections were raised and a decision on this issue was postponed. It is my personal view that, although the adoption of a new nomenclature may perturb (minimally) some of our daily routines, the advantages, including a satisfactory nomenclature of human genes, far outweigh the inconveniences.
Figure 1.
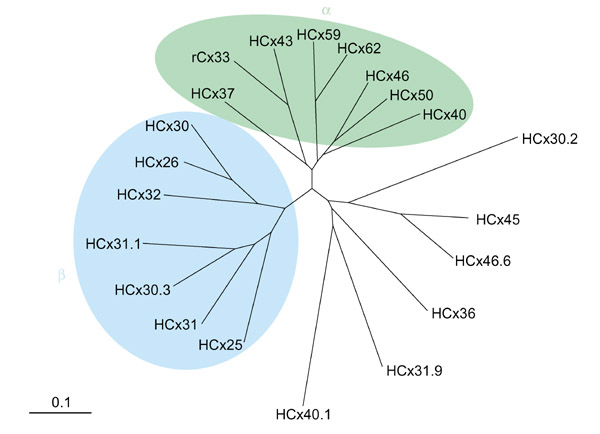
A molecular phylogenetic tree for human connexin protein sequences (HCx) and rat connexin 33 (rCx33). The divergent sequence portions (the intracellular loop and the carboxy-terminal domain) were removed from the multiple alignments. The distance matrices were then analyzed by the neighbor-joining method of Saitou and Nei (Mol Biol Evol 1987, 4:406-425), producing an unrooted molecular phylogenetic tree. The connexin family can be broadly separated into three main subgroups, the historical α (green) and β (blue) and a more divergent non-α non-β branch. A sequence divergence of 10% is indicated by the scale bar. The tree is provided courtesy of Daniele Condorelli (University of Catania, Italy).
Probing structure
Connexin topology calls for cytoplasmic amino and carboxyl termini, one cytoplasmic and two extracellular loops, and four transmembrane domains. One key question lingering over the connexin channel is the identification of the domains and amino acids that form the channel's pore. The classical approach of mutagenesis remains quite effective for addressing this issue; the laboratories of Vytas Verselis (Albert Einstein College of Medicine, USA) and Bruce Nicholson (State University of New York at Buffalo, USA) have used it and other methods to probe the extracellular and transmembrane regions of the pore, respectively. Jack Kronengold from the Verselis lab showed that negatively charged amino acids in the first extracellular loop of Cx46 are pore-lining residues, by demonstrating their accessibility to modification by methanethiosulfonate (MTS) added to either side of an open hemichannel. The resulting reductions of single-channel conductance occurred in a stepwise fashion, suggesting consecutive hits of the MTS on each connexin subunit. Martha Skerrett from the Nicholson lab used the substituted-cysteine accessibility method applied to intact gap junction channels to identify the third transmembrane domain as the main pore-lining domain on the basis of the reactivity to the thiol reagent maleimidobutyryl biocytin (MBB). Aside from studying two different regions of the pore, some other characteristics distinguished these studies. Kronengold used a smaller probe and measured single channel activity, which should have given him a better accuracy; on the other hand, Skerrett analyzed the behavior of intercellular channels, not hemichannels as in Kronengold's experiments, and it is possible that a complete gap junction channel is not the simple sum of two hemichannels.
Using a similar strategy, Luis Barrio (Hospital Ramón y Cajal, Madrid, Spain) identified in the second extracellular loop a calcium-binding site that blocks the fully voltage-gated opening of Cx32 hemichannels. Clearly, new tools are also necessary to hone in on channel structure. One approach was illustrated by Gina Sosinsky (University of California at San Diego, USA), who has used atomic force microscopy to obtain images that show exquisite detail at the extracellular surface of force-dissected gap junctions (that is, looking at the outside face of a hemichannel). This technique allows the sample to be viewed while the solutions bathing it are changed, and the connexon can be imaged under conditions in which the channel is either open or closed.
Probing function
The full panoply of genetic weapons available to investigate protein function - knockout, knock-in, RNA-mediated interference (RNAi) and antisense oligonucleotides - were on display at the meeting. Britta Doering (University of Bonn, Germany) reported the generation of several transgenic mouse lines with cell-type-specific deletion of Cx43, which allowed her to overcome the perinatal lethality of germline Cx43 deficiency. Mice with the Cx43 gene flanked by loxP sites were crossed with separate lines of transgenic animals in which Cre recombinase was driven by cell-type-specific promoters for astrocytes, endothelial cells, pancreatic β-cells, hepatocytes and cardiac myocytes. Of note, conditional deletion of Cx43 in the heart led to morphological abnormalities, albeit distinct from those described in Cx43-null mice, and death within the first two weeks after birth. These results were somewhat different from those presented by Greg Morley (Mount Sinai School of Medicine, New York, USA) who described mice with normal heart histology and contractile performance but markedly slowed conduction; these eventually died suddenly with spontaneous ventricular arrhythmia. Given that both groups crossed the same mouse strain expressing Cre under the control of the a myosin heavy chain with 'floxed' Cx43 animals, these discrepancies will no doubt be resolved in the near future.
The same two labs also presented further work addressing the role of Cx43 in heart function. Morley described the phenotype of mice with 'patchy' expression of Cx43 in the myocardium, which resulted in conduction defects and markedly depressed contractile function, and he hinted that conditional deletion of Cx43 in neural crest cells, which play a major role in cardiac morphogenesis, is in the pipeline. Willecke and Karen Maass have replaced wild-type Cx43 with a pH-insensitive Cx43 mutant truncated at amino acid 257, which also loses most of the phosphorylation sites that would otherwise be required for disruption of gap-junctional intercellular communication. Preliminary data indicate that these 'knock-in' mice die within the first week after birth, suggesting that the major regulatory domain in the carboxy-terminal tail of Cx43 is of vital importance.
The completed genomic sequences of the fruitfly and nematode worm proved that nobody had found connexins in invertebrates for the simple reason that there are none, and both these organisms use instead a different, but functionally equivalent, family of proteins, the innexins. In order to identify innexins involved in embryogenesis and wiring of the nervous system, Todd Starich (University of Minnesota, St Paul, USA) generated antibodies or green fluorescent protein fusions for 17 of the 25 innexins present in C. elegans, thus establishing an initial map of innexin expression. He reported that there are already three innexins expressed at the two-cell stage of nematode development, and that the same phenotype was observed following RNAi-based deletion of two innexins, strongly suggesting co-assembly of these two proteins into the same intercellular channel.
The usefulness of antisense techniques was discussed by Yossi Landesman (Harvard Medical School, Boston, USA), who explored the role of cell-cell communication in Xenopus development. Not only did he show that at least four maternal connexins are expressed in the oocyte and early embryo of Xenopus, but a striking phenotype was observed after depletion of Cx43 by morpholino antisense oligonucleotides: tadpoles were normally patterned, but paralyzed. The loss of locomotion could result from defects in sensory and/or motor pathways, which are being currently investigated.
Classical functional expression studies can also still yield vital information. Connexins are endowed with regulatory mechanisms whose functional significance is not obvious. One interesting case is posed by voltage gating, which is closure of connexin channels in response to transjunctional voltage. As Mike Bennett (Albert Einstein College of Medicine, New York, USA) put it in a review article some time ago, "voltage dependence of connexins is an arcane subject that may be of interest only to biophysicists, and a subset of them at that". Now Yang Qu and Gerhard Dahl (University of Miami, USA) have performed an elegant study to offer a fresh view of the issue. They have used paired Xenopus oocytes; in each pair, the pre-junctional cell expresses Cx46 and the post-junctional oocyte expresses Cx43 and a reporter gene, in this case the cystic fibrosis transmembrane conductance regulator (CFTR). They took advantage of the odd property of Cx46, which forms open hemichannels in Xenopus oocytes, to load the pre-junctional cell with either fluorescent probes or cAMP, and then measured the transit of test molecules through heterotypic Cx46/Cx43 channels at different transjunctional voltages (± 20 mV) that did not significantly reduce macroscopic conductance. When the voltage gate of Cx46 was activated, not only was the accumulation of tracers reduced, but the cAMP-activated chloride current carried by CFTR in the post-junctional cell was also reduced. These data provide compelling evidence that, within a range of transjunctional potentials, activation of the voltage gate selectively restricts metabolic coupling, but not electrical coupling. Thus, a possible physiological function of the voltage gate would be to modulate the passage of larger signaling molecules, such as cAMP, between cells without perturbing electrical coupling.
Connexins and diseases
The first disease to be associated with connexin mutations was the X-linked form of Charcot-Marie-Tooth (CMTX). One of the puzzling aspects of CMTX remains the exquisite restriction of symptoms to the peripheral nervous system, although the gene mutated in the disease, Cx32, is expressed in many other cells, including oligodendrocytes that are capable of forming normal myelin sheathes in the central nervous system (CNS). A popular explanation has been that most cell types express multiple connexins, which can functionally replace the mutated gene. Now the laboratories of David Paul (Harvard Medical School, Boston, USA) and Klaus Willecke have reported the identification of more connexins in Schwann cells and oligodendrocytes. Interestingly, Daniela Menichella (Harvard Medical School, Boston, USA) provided compelling evidence that oligodendrocytes are the major source of Cx47, and that a double knockout mouse produced by crossing Cx47-/- (phenotypically normal) mice with Cx32-/- mice resulted in a very severe phenotype resembling that of the shiverer mouse, in which the gene encoding myelin basic protein is deleted. Thus, Cx32 and Cx47 supply redundant activities necessary for oligodendrocyte-dependent myelination.
The most important connexin-based disease in terms of public health impact is a recessive form of deafness, DFNB1, which accounts for about half of the cases of inherited deafness in Western countries (1 in 2,000 children) and is caused by mutations in Cx26. The lethality of germline deletion of Cx26 in mice led Martine Cohen-Salmon (Institut Pasteur, Paris, France) to address the function of Cx26 in the inner ear, by generating mice with targeted Cx26 ablation in the sensory epithelial network. These mice exhibited hearing impairment and a progressive massive disorganization of the neuroepithelium of the cochlea, revealing that Cx26-containing epithelial gap junctions play an essential role in the function of the cochlea, probably linked to the elimination of glutamate and/or K+ ions released by the hair cells upon sound stimulation. In this context, Fabio Mammano (International School for Advanced Studies, Trieste, Italy) provided convincing electrophysiological evidence for a short recycling route of K+ ions that are taken up by supporting Deiters cells within the cochlea and then are shunted through gap junction channels back to the endolymph, via Hensen cells which have an apical K+/Cl- co-transport allowing for the maintenance of the endolymphatic potential. Two other connexins, Cx30 and Cx31, are responsible for dominant forms of deafness. Although Cx31 deficiency in mice does not result in an auditory deficit, Barbara Teubner (University of Bonn, Germany) found that homozygous Cx30-null mice do exhibit a severe hearing impairment. Cx26 and Cx30 are both expressed within the cochlea (in the spiral limbus, the spiral ligament, the stria vascularis and between supporting cells of the organ of Corti), but it looks as though they are not interchangeable, and a precise combination of both genes is needed for a normal auditory function.
Connexins are also involved in a number of skin disorders that are both clinically and genetically heterogeneous. Gabriela Richard (Thomas Jefferson University, Philadelphia, USA) identified a family with a novel Cx26 mutation associated with a disorder diagnosed as Bart-Pumphrey syndrome (deafness, palmoplantar keratoderma and nail abnormalities), stressing the clinical overlaps between defects in Cx26, Cx30, Cx30.3 and Cx31 that can be best summarized as: two connexins, one disorder; one connexin, many disorders. To gain insight into the pathophysiological mechanisms of these mutations, Ludovic Wiszniewski (University of Geneva, Switzerland) has used a three-dimensional skin-culture system, in which epidermal equivalents are reconstituted in vitro with stem cells from hair follicles. Using keratinocytes from DFNB1 patients (mutated Cx26), a reduced stratification of epidermal equivalents was observed, whereas keratinocytes from individuals affected with Clouston syndrome (mutated Cx30) exhibited a normal stratification but a thickened parakeratotic cornified layer. This is the first experimental evidence that different connexins in the skin perform separate tasks.
As one would expect, connexins may play a role not only in genetic diseases but also in cardiovascular pathology. Brenda Kwak (University of Geneva, Switzerland) presented intriguing data suggesting that a reduction of Cx43 expression in vivo (using Cx43+/- heterozygous mice) drastically reduced the progression of atherosclerotic lesions in mice lacking the low density lipoprotein (LDL) receptor, a model of atherosclerosis. Given that atherosclerosis is the leading cause of death in Western societies, this hypothesis will be examined in great detail in the near future.
The case of electrical synapses
Ultrastuctural and functional studies have, over the years, supported the notion that communication in the mammalian brain occurs overwhelmingly through chemical synapses. This monolithic view is now being amended by several reports on the distribution and functional roles of electrical synapses - gap junctions between adjacent neurons. John Rash (Colorado State University, Fort Collins, USA) has applied freeze-fracture immunogold labeling to the rat CNS, to examine the cell-type-specific expression of four connexins, including Cx36 which was identified as the first neuronal connexin isoform. After looking at over 5,000 labeled gap junctions, he concluded that gap junctions are present between neurons (where they are made of Cx36), between astrocytes (Cx43 and Cx30), and between astrocytes (Cx43 and Cx30) and oligodendrocytes (Cx32). Thus, Rash found no structural evidence of direct communication between neurons and glia, and suggested that neuron-glia coupling occurs via other routes. This is clearly a first expression map, as more connexins are being found in the CNS and specific antibodies are becoming available.
The cerebral cortex generates a variety of synchronized patterns of activity that vary with behavioral state. To determine the role of connexins in organizing these neuronal networks, Mike Deans and David Paul (Harvard Medical School, Boston, USA) constructed mice carrying histochemical reporters in place of Cx36, which is expressed in brain and retina. In collaboration with Barry Connors (Brown University, Providence, USA), they found that electrical synapses were common among inhibitory cortical interneurons in controls but were nearly absent from the knockouts. Furthermore, synchronous firing was weaker and spatially restricted, leading them to conclude that electrical synapses containing Cx36 are critical for the generation of widespread, synchronous inhibitory activity.
Electrical coupling is widespread among retinal neurons and regulates several aspects of visual function. The analysis of the retina of these mice showed several interesting features, such as expression in AII amacrine cells (interneurons in the rod photoreceptor pathway) and two types of ON-cone bipolar cells (see Figure 2), as well as reduced coupling between AII amacrine cells. The Willecke lab also presented similar data, together with Reto Weiler (University of Oldenburg, Germany). More importantly, in collaboration with Stewart Bloomfield (New York University School of Medicine, USA), Deans provided definitive physiological evidence that the loss of Cx36 disrupts AII amacrine to ON-cone bipolar cell coupling, which is a critical relay for transmission of the visual signal from rods to ganglion cells under starlight conditions. Novel insights into neuronal circuit organization are emerging from these studies, and one can easily anticipate that this outpouring of information will help to determine how electrical synapses and the networks they define are involved in brain function.
Figure 2.
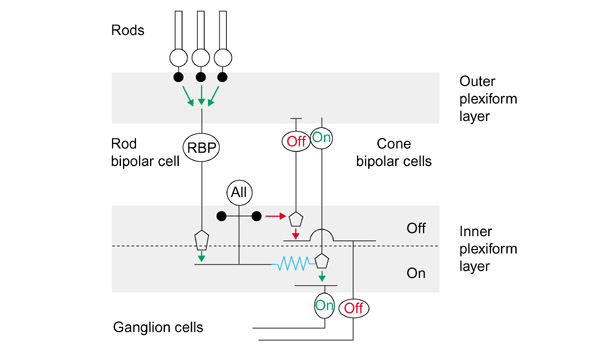
The mammalian retina uses electrical synapses in the vertical pathway from rod photoreceptors to ON-ganglion cells. In scotopic view, rod bipolar cells send signals to rod bipolar cells to excite AII amacrine cells through conventional synapses (green arrows). The AII cell forms electrical synapses (blue jagged line) with ON-cone bipolar cells, thus parasitizing the cone pathway. This pathway is lost in Cx36 knockout animals. Red arrows denote chemical inhibitory synapses. The diagram is modified from Strettoi, Dacheux and Raviola (J Comp Neurol 1994, 347:139-149).
In summary, this conference provided several clues that are contributing to our painstaking efforts to understand con-nexin syntax. New techniques, tools and mouse lines are being developed, other genetic approaches are becoming available as researchers working with worms and flies are studying the innexins, and naturally occurring mutations are illuminating the exquisitely controlled roles of connexins in different organs.
A collection of papers presented at the meeting will be published in a special issue of Cell Communication and Adhesion at the end of the year. The next International Gap Junction Conference will be held in Cambridge, UK, in 2003.