
Figure 1. Simulated entorhinal place cells from Eq. 1 (Adapted from Howard et al, 2005).
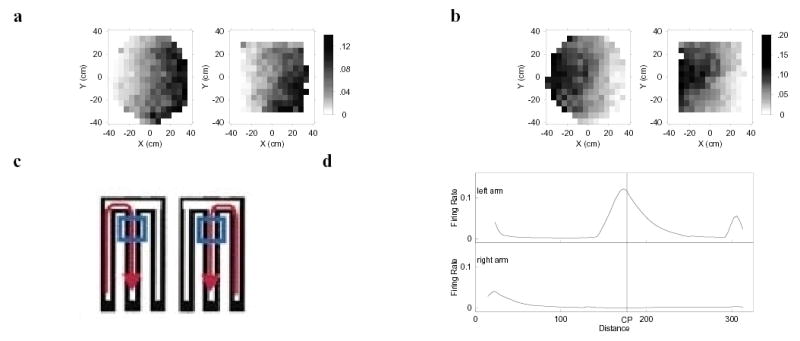
a,b. Simulated entorhinal place fields from the open field. Like entorhinal cells (Quirk et al, 1992), the simulated cells showed noisy place fields that were correlated across similar environments. The cell in a took input from a head direction cell with a preferred direction that pointed towards the east. The cell in b received input from a head direction cell with a preferred direction that pointed west-by-northwest. c. Frank et al, (2000) trained rats to run in a W-maze in which the animal successively visited a food well in each arm in sequence. This paradigm provides the opportunity to examine firing of cells in the middle arm of the maze as part of different trajectories. d. Simulated entorhinal cells showed retrospective coding that distinguished the path the animal had taken to reach a point on the middle arm. A representative simulated retrospective cell shows different firing on the center arm on inbound paths depending on which arm the animal was coming from. The choice points, CP, is a point along the path that is several centimeters from the top of the middle arm. The choice point is indicated by the top of the box in c. Because position and heading are the same in both cases (see c), this indicates that the model, like the entorhinal cortex, is responding to more than position per se. See Howard et al 2005 for details.