
Figure 1.
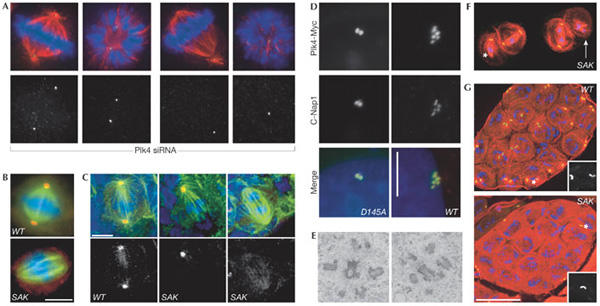
Plk4/Sak regulates centrosome duplication. (A) Small interfering RNA targeting of human Plk4 induces progressive aberrations in spindle morphology and a stepwise reduction in centriole numbers (green, centrin; red, α-tubulin; blue, DNA). (B,C) Drosophila Sak is required for centrosome duplication in S2 cells (B) and larval brain cells from Sak flies (C) (green, α-tubulin; blue, DNA; red, γ-tubulin). Scale bars, 5 μm. (D) Staining for the centriole marker C-Nap1 (red) and the transfected Plk4–myc fusion protein (green) reveals an increase in centriole number following overexpression of wild-type (WT), but not catalytically inactive (D154A) Plk4–Myc in human U2OS cells. Scale bar, 10 μm. (E) A transmission electron micrograph of Plk4–Myc-expressing U2OS cells 48 h after induction of transcription shows the centriole content of the cells. (F) Primary Sak spermatocytes in meiosis I stained to reveal microtubules (red), GFP-PACT (a centriolar marker) and DNA (blue). The asterisk marks an unfocused pole that lacks a centriole whereas a highly disorganized spindle that completely lacks centrioles is indicated by the white arrow. (G) Wild-type and SAK primary spermatocyte cysts stained as in (F). There are 16 cells in each cyst but only the wild-type cyst has a centriole pair in each cell. Scale bar, 50 μm. Panels (B,C,F,G) are reprinted from Bettencourt-Dias et al (2005) with permission from Elsevier. Panels (A,D,E) reprinted from Habedanck et al (2005) with permission from Nature Publishing Group.