

Abstract
Centella asiatica (CeA) is a creeping plant growing in damp places in India and other Asian countries. The leaves of CeA are used for memory enhancement in the Ayurvedic system of medicine, an alternative system of medicine in India. In this study, we have investigated the effect during the rat growth spurt period of CeA fresh leaf extract treatment on the dendritic morphology of hippocampal CA3 neurons, one of the regions of the brain concerned with learning and memory. Neonatal rat pups (7 days old) were fed with 2, 4 or 6 ml kg−1 body weight of fresh leaf extract of CeA for 2, 4 or 6 weeks. After the treatment period the rats were killed, their brains were removed and the hippocampal neurons were impregnated with silver nitrate (Golgi staining). Hippocampal CA3 neurons were traced using a camera lucida, and dendritic branching points (a measure of dendritic arborization) and intersections (a measure of dendritic length) were quantified. These data were compared with data for age-matched control rats. The results showed a significant increase in the dendritic length (intersections) and dendritic branching points along the length of both apical and basal dendrites in rats treated with 4 and 6 ml kg−1 body weight per day of CeA for longer periods of time (i.e. 4 and 6 weeks). We conclude that the constituents/active principles present in CeA fresh leaf extract have a neuronal dendritic growth stimulating property; hence, the extract can be used for enhancing neuronal dendrites in stress and neurodegenerative and memory disorders.
Keywords: Centella asiatica, dendritic branches, dendritic intersections, growth spurt period, hippocampal CA3 neurons
Introduction
Ayurveda, an alternative system of medicine in India, uses a number of plants for the treatment of a variety of diseases. The Medhya rasayana are a group of medicines in Ayurveda known to act on the nervous system. These drugs contain mainly extracts from plants such as Centella asiatica (CeA), Acorus calamus, jatamansi and Bacopa monnieri. These medhya rasayana have been claimed to improve mental ability (1).
CeA is a herb that grows in wet places throughout India. It is used in Ayurvedic preparations either as whole plants or as leaves in the fresh or extract form (1). CeA has been shown to be very useful in improving learning and memory (2–4). It is also used as a tonic for promoting brain growth and improving memory (5). In addition, the plant is used in children with learning difficulties to improve general mental ability and in people suffering from cognitive disorders (3,6–9). Though the fresh leaf juice (extract) of CeA has been claimed to improve learning and memory in different clinical studies (2,3,6–8), there is no evidence to show the effect of this plant extract on the brain regions involved in learning and memory, namely the hippocampus (10–12), amygdala and limbic cortex. The cornu ammonis (CA) region, particularly the CA3 subregion of the hippocampus, is the key structure of the brain involved in learning and memory (13–16).
We hypothesized that treatment with fresh leaf juice from CeA would bring about structural changes in the hippocampal CA3 pyramidal neurons in young growing rats. Since active brain growth occurs during the growth spurt (preweaning) period (17), our study was carried out on neonatal rats. We have aimed to conduct the experiment in the way explained in the classic texts of Ayurveda (1), i.e. using fresh leaf juice rather than extraction.
Methods
Rats
Neonatal Wistar rat pups (7 days old) of both sexes bred and maintained in our central animal research facility were used in this experiment. The rats were fed food and water ad libitum and maintained in a 12:12 h dark and light cycle. The room temperature was kept constant at 25°C. All experiments were carried out with prior approval from the institutional animal ethical committee. Only the minimum required number of rats were used, and they were handled in a humane way.
Experimental Groups
Rat pups were assigned into 2, 4 and 6 week treatment groups. Pups in each of these groups were divided into 2, 4 and 6 ml kg−1 body weight dose groups (n = 8 for each dose). Each rat pup in the given dosage group was fed through gastric intubation with a given amount of fresh leaf extract of CeA daily for 2, 4 or 6 weeks. Along with these experimental groups, a normal control group and a vehicle (saline) control group (n = 8 in both groups) were also maintained.
Extraction and Administration of CeA Leaf Juice
The CeA was identified by Professor P. Venugopal Tantry, Department of Botany, Vijaya College, Mulky, Karnataka, India. A voucher specimen number 525PP has been entered in the registry in the Department of Pharmocognosy, Manipal College of Pharmaceutical Sciences, Manipal, India. For this experiment, we cultivated the plant in uniform soil conditions in order to maintain the same source of plant throughout the experiment. Fresh, 15–20-day-old leaves of CeA were collected in the morning. Fresh leaf juice was extracted from these leaves after washing, air-drying and homogenizing in a glass vessel and finally filtering through a sterile gauge cloth. Leaves were extracted maximally, so that from a given weight of leaves (5.0 g), a known volume of juice was extracted (1.63 ± 0.15 g, n = 6). Since soil water was kept uniform we could extract the same volume of juice from a given weight of leaves on different days. Further, we have established that the dry weight of a given volume (1 ml) of juice prepared on different days was the same (0.079 ± 0.01 g, n = 6). The fresh leaf juice so obtained was fed to the rat pups through a gastric capillary tube attached to a 1 ml hypodermic syringe. Since the volume of juice to be fed to each individual rat pup was very little, the dose was blended with an appropriate volume of saline for convenient feeding. Control rats remained undisturbed in their home cage, and saline control rats were fed with a volume of saline equivalent to the volume of extract that the age-matched experimental rats received on each day.
Since standard extraction procedures (which involve boiling in water, ethyl alcohol or other organic solvents) may alter the structure of bioactive principles, we avoided them. Though there may have been minor variation in our daily preparations, it would have been minimal as leaves of equal maturation were collected from the same place on all days. This minor daily variation would have been compensated for by the long period (2, 4 or 6 weeks) of treatment. It has been shown in a recent study that a CeA plant extract obtained from ethanol extraction was different from one obtained from water extraction in terms of its biological activity (18).
Rapid Golgi Staining Procedure (19)
At the end of the extract treatment period (2, 4 or 6 weeks), the rats were deeply anesthetized with ether and killed, their brains were removed rapidly and the hippocampi were dissected and fixed in rapid Golgi fixative. Tissue was processed for rapid Golgi staining as detailed earlier (19). In brief, tissues were fixed for 5 days in Golgi fixative and impregnated with 0.75% aqueous silver nitrate for 48 h. Sledge microtome sections of 120 μm thickness were taken, dehydrated, cleared and mounted with DPX mounting media.
Camera Lucida Tracing
Slides were coded before camera lucida tracing and quantification to avoid experimenter bias. Decoding was performed after the completion of data collection and analysis. From each rat, 8–10 hippocampal CA3 neurons were traced using the camera lucida, and their dendritic branching points and dendritic intersections were quantified. Those neurons with minimal overlap of dendrites, heavily impregnated with silver nitrate and without truncate dendrites were selected for tracing.
Quantification of Dendritic Branching Points and Dendritic Intersections
The concentric circle method of Sholl (20) was used for dendritic quantification. On a transparent sheet, concentric circles were drawn. The radial distance between two adjacent concentric circles was 20 μm. For dendritic quantification, the sheet with concentric circles was placed on the camera lucida trace of the neuron in such a way that the center of the cell body of the neuron coincided with the center of the concentric circles. The number of branching points between two successive concentric circles, i.e. within each successive 20 μm radial circle, was counted. The dendritic intersection is the point where a dendrite intersects the given concentric circle. Both branching points and intersections were counted up to a radial distance of 100 μm from the center of the soma. The mean number of dendritic branching points in each concentric zone/neuron and the number of dendritic intersections at each concentric circle/neuron were calculated. This method of scoring was applied to both apical and basal dendritic quantification.
Data Analysis
Data were analyzed using analysis of variance (ANOVA) followed by Bonferroni's post test using Graph Pad in Stat (GPIS) software, version 1.13.
Results
The rats treated with all doses of CeA remained healthy throughout the treatment period. They gained more body weight than the control and saline treated rats (data not shown). Moreover, the rats fed with CeA leaf extract were more active than control rats, and learning and memory were significantly better in rats treated with higher doses for longer periods (21).
Hippocampal CA3 Neuronal Dendritic Quantification
The hippocampal CA3 neuronal dendritic analysis in CeA leaf extract treated rats showed a significant increase in dendritic length and branching in both apical and basal dendrites, particularly at higher doses (4 and 6 ml kg−1 CeA) for longer durations (4 and 6 weeks). Figures 1 and 6 illustrate the dendritic arborization of hippocampal CA3 neurons in control rats and rats treated for 4 and 6 weeks with CeA leaf extract. There was no difference in dendritic length and branching pattern between control and saline treated rats, suggesting that daily handling of the rats (handling stress and vehicle) itself did not alter the dendritic pattern. On this basis, only comparisons between normal control and experimental groups are detailed below and in all figures and tables.
Figure 1.

Representative photomicrographs and camera lucida tracings of hippocampal CA3 neurons from control rats (A1–2) and rats treated with CeA for 6 weeks [2 ml kg−1 CeA (B1–2); 4 ml kg−1 CeA (C1–2); 6 ml kg−1 CeA (D1–2)]. Note the significant increase in dendritic length and branches in rats treated with 4 and 6 ml kg−1 CeA for 6 weeks. Scale bar = 20 μm.
Significant Increase After 6 Weeks
There was a significant increase in the dendritic intersections and branching points in the CeA leaf extract treated rats at different radial distance from the soma. Figure 1 shows photomicrographs and camera lucida tracings of normal control rats (A1–2) and rats treated with 2, 4 and 6 ml kg−1 of CeA leaf extract for 6 weeks (B1–2, C1–2 and D1–2, respectively). Note that these Golgi stained (silver nitrate impregnated) neurons have a significantly increased number of dendrites extending beyond 100 μm from the soma in the 4 and 6 ml kg−1 CeA leaf extract treated rats.
Apical Dendritic Intersections
Both 4 and 6 ml kg−1 CeA groups treated for 6 weeks showed a significant increase in the apical dendritic intersections at the 80 μm (77.07 and 112.62% increases) and 100 μm (137.0 and 151.62% increases) concentric circles (80 μm concentric circle: 3.01 ± 0.8 in normal controls versus 5.33 ± 0.95 in the 4 ml kg−1 CeA group, P < 0.01, and 6.4 ± 0.86 in the 6 ml kg−1 CeA group, P < 0.001; 100 μm concentric circle: 3.08 ± 1.15 in normal controls versus 7.31 ± 1.0 in the 4 ml kg−1 CeA group, P < 0.001, and 7.75 ± 0.54 in the 6 ml kg−1 CeA group, P < 0.001). In addition, the 6 ml kg−1 CeA group showed a significantly increased number of dendritic intersections (69.2% increase) at the 60 μm concentric circle (2.5 ± 0.31 in normal controls versus 4.23 ± 0.82 in the 6 ml kg−1 group, P < 0.001). There were no significant changes in the dendritic intersections at any of the concentric circles in the 2 ml kg−1 CeA group when compared with the control group (Fig. 2).
Figure 2.
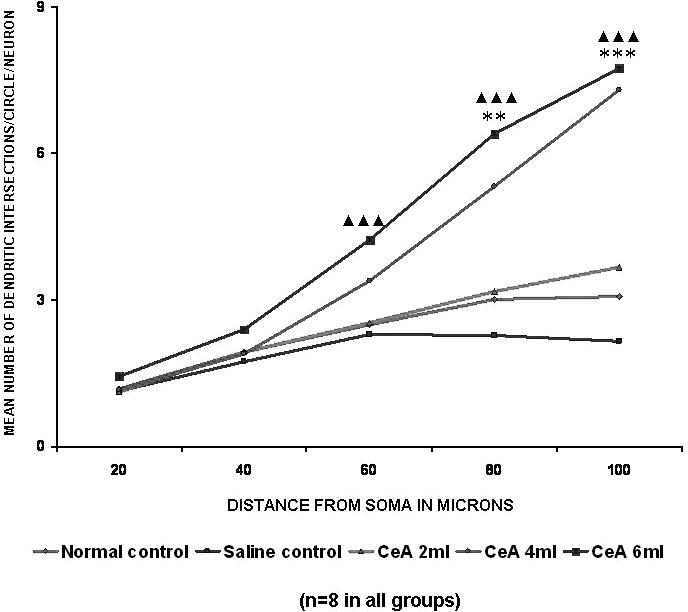
Apical dendritic intersections of hippocampal CA3 neurons in rats treated with 2, 4 and 6 ml kg−1 body weight of CeA for 6 weeks and in age-matched control and saline treated rats. Each point represents the mean of 8–10 neurons from each rat (SD not shown). F-value: 1.27, 3.17, 17.22, 37.49 and 64.43 at 20, 40, 60, 80 and 100 μm distance from the soma, respectively. Note that there is a significant increase in the dendritic intersections in rats treated with 4 and 6 ml kg−1 CeA. Normal control versus 4 ml kg−1 CeA: **P < 0.01, ***P < 0.001; normal control versus 6 ml kg−1 CeA: ▴▴▴P < 0.001 (one-way ANOVA, Bonferroni's test).
Basal Dendritic Intersections
The 2, 4 and 6 ml kg−1 CeA groups showed a significant increase in the basal dendritic intersections at the 60 μm (53.16, 111.86 and 108.06% increases) and 80 μm (79.26, 169.54 and 190.72% increases) concentric circles (60 μm concentric circle: 6.32 ± 1.71 in normal controls versus 9.68 ± 1.32 in the 2 ml kg−1 CeA group, P < 0.05, 13.39 ± 1.21 in the 4 ml kg−1 CeA group, P < 0.001, and 13.15 ± 2.33 in the 6 ml kg−1 CeA group, P < 0.001; 80 μm concentric circle: 4.63 ± 1.53 in normal controls versus 8.30 ± 2.77 in the 2 ml kg−1 CeA group, P < 0.05, 12.48 ± 0.92 in the 4 ml kg−1 CeA group, P < 0.001, and 13.46 ± 1.96 in the 6 ml kg−1 CeA group, P < 0.001). In addition, the 4 and 6 ml kg−1 CeA groups also showed a significantly increased number of dendritic intersections at the 40 μm (64.71 and 65.17% increases) and 100 μm (262.44 and 331.83% increases) concentric circles (40 μm concentric circle: 6.49 ± 1.28 in normal controls versus 10.69 ± 0.99 in the 4 ml kg−1 CeA group, P < 0.001, and 10.72 ± 1.67 in the 6 ml kg−1 CeA group, P < 0.01; 100 μm concentric circle: 2.45 ± 0.82 in normal controls versus 8.88 ± 1.63 in the 4 ml kg−1 CeA group, P < 0.001, and 10.58 ± 2.46 in the 6 ml kg−1 CeA group, P < 0.001) (Fig. 3).
Figure 3.
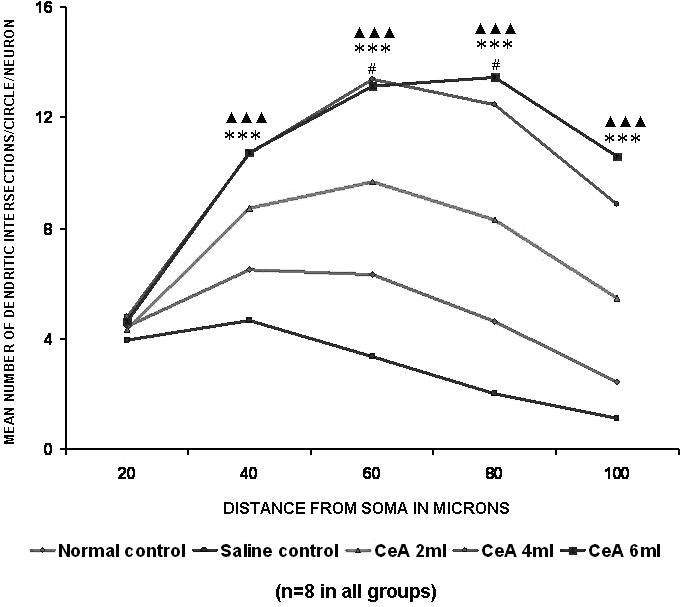
Basal dendritic intersections of hippocampal CA3 neurons in rats treated with 2, 4 and 6 ml kg−1 body weight of CeA for 6 weeks and in age-matched control and saline treated rats. Each point represents the mean of 8–10 neurons from each rat (SD not shown). F-value: 0.73, 38.64, 58.37, 60.64 and 33.0 at 20, 40, 60, 80 and 100 μm distance from the soma, respectively. Note that there is a significant increase in the dendritic intersections in rats treated with 2, 4 and 6 ml kg−1 CeA. Normal control versus 2 ml kg−1 CeA: #P< 0.05; normal control versus 4 ml kg−1 CeA: ***P < 0.001; normal control versus 6 ml kg−1 CeA: ▴▴▴P < 0.001 (one-way ANOVA, Bonferroni's test).
Apical Dendritic Branching Points
Both the 4 and 6 ml kg−1 CeA groups showed a significant increase in the apical dendritic branching points in the fourth (60–80 μm) zone [1.22 ± 0.61 in normal controls versus 2.49 ± 0.59 in the 4 ml kg−1 CeA group (104.09% increase), P < 0.05, and 2.69 ± 0.56 in the 6 ml kg−1 CeA group (120.49% increase), P < 0.01] and the fifth (80–100 μm) zone [0.88 ± 0.56 in normal controls versus 2.81 ± 0.66 in the 4 ml kg−1 CeA group (219.31% increase), P < 0.01, and 2.57 ± 0.87 in the 6 ml kg−1 CeA group (192.04% increase), P < 0.01]. In addition, in the third (40–60 μm) zone, the 6 ml kg−1 CeA group showed a significantly increased number of dendritic branching points [0.92 ± 0.3 in normal controls versus 2.09 ± 0.63 in the 6 ml kg−1 CeA group (127.17% increase), P < 0.01]. However, no significant change was observed in the dendritic branching points in any of the concentric zones in the 2 ml kg−1 CeA group (Fig. 4).
Figure 4.
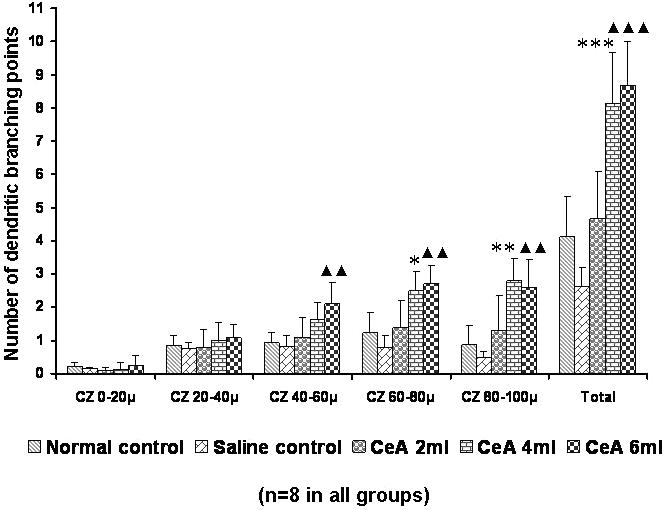
Apical dendritic branching points in the hippocampal CA3 neurons of rats treated with 2, 4 and 6 ml kg−1 body weight of CeA for 6 weeks and in age-matched control and saline treated rats at concentric zones (CZs) and total number of branching points. Each value represents the mean + SD of 8–10 neurons from each rat. F-value: 0.91, 0.98, 9.70, 15.63, 16.24 and 35.43 at the 0–20, 20–40, 40–60, 60–80 and 80–100 μm CZs and total number of branching points, respectively. Note the significant increase in dendritic branching points in 4 and 6 ml kg−1 CeA treated rats compared with control rats. Normal control versus 4 ml kg−1 CeA: *P < 0.05, **P < 0.01, ***P < 0.001; normal control versus 6 ml kg−1 CeA: ▴▴P < 0.01, ▴▴▴P < 0.001 (one-way ANOVA, Bonferroni's test).
Basal Dendritic Branching Points
All the groups treated with CeA showed a significant increase in the dendritic branching points in the second (20–40 μm) zone [2.83 ± 0.58 in normal controls versus 4.71 ± 0.67 in the 2 ml kg−1 CeA group (66.43% increase), P < 0.05, 6.0 ± 0.63 in the 4 ml kg−1 CeA group, P < 0.001, and 6.09 ± 1.25 in the 6 ml kg−1 CeA group (27.38% increase), P < 0.001] and the third (40–60 μm) zone [1.37 ± 0.83 in normal controls versus 3.45 ± 0.43 in the 2 ml kg−1 CeA group (151.82% increase), P < 0.01, 4.29 ± 0.73 in the 4 ml kg−1 CeA group (213.14% increase), P < 0.001, and 4.76 ± 1.04 in the 6 ml kg−1 CeA group (247.44% increase), P < 0.001]. In the fourth (60–80 μm) zone both the 4 and 6 ml kg−1 CeA groups showed a significantly increased number of dendritic branching points [0.7 ± 0.59 in normal controls versus 1.99 ± 0.56 in the 4 ml kg−1 CeA group (184.28% increase), P < 0.01, and 2.57 ± 0.51 in the 6 ml kg−1 CeA group (267.42% increase), P < 0.001]. In the fifth (80–100 μm) zone the 6 ml kg−1 CeA group showed a significantly increased number of dendritic branching points (0.08 ± 0.14 in normal controls versus 0.99 ± 0.49 in the 6 ml kg−1 CeA group, P < 0.01) (Fig. 5).
Figure 5.
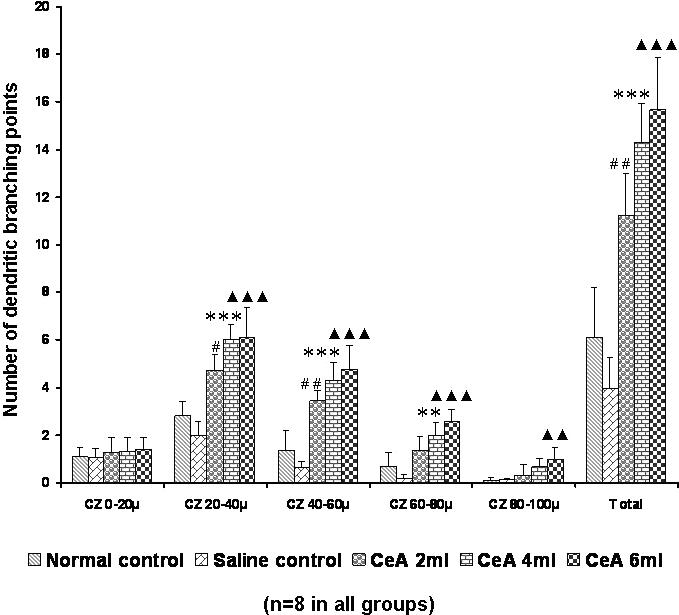
Basal dendritic branching points in the hippocampal CA3 neurons of rats treated with 2, 4 and 6 ml kg−1 body weight of CeA for 6 weeks and in age-matched control and saline treated rats at concentric zones (CZs) and total number of branching points. Each value represents the mean + SD of 8–10 neurons from each rat. F-value: 0.74, 44.64, 51.38, 28.66, 10.06 and 61.49 at the 0–20, 20–40, 40–60, 60–80 and 80–100 μm CZs and total number of branching points, respectively. Note the significant increase in dendritic branching points in 2, 4 and 6 ml kg−1 CeA treated rats compared with control rats. Normal control versus 2 ml kg−1 CeA: #P < 0.05, ##P < 0.01; normal control versus 4 ml kg−1 CeA: **P < 0.01, ***P < 0.001; normal control versus 6 ml kg−1 CeA: ▴▴P < 0.01, ▴▴▴P < 0.001 (one-way ANOVA, Bonferroni's test).
Results of 4 Weeks' Treatment Group
There was a significant increase in the dendritic intersections and branching points in the CeA leaf extract treated rats at different radial distances from the soma. Figure 6 shows photomicrographs and camera lucida tracings of normal control rats (A1–2) and rats treated with 2, 4 and 6 ml CeA leaf extract for 4 weeks (B1–2, C1–2, D1–2, respectively). Note that these Golgi stained (silver nitrate impregnated) neurons have a significantly increased number of dendrites extending beyond 100 μm from the soma in the 4 and 6 ml kg−1 CeA leaf extract treated rats.
Figure 6.

Representative photomicrographs and camera lucida tracings of hippocampal CA3 neurons from control rats (A1–2) and rats treated with CeA for 4 weeks [2 ml kg−1 CeA (B1–2); 4 ml kg−1 CeA (C1–2); 6 ml kg−1 CeA (D1–2)]. Note the significant increase in dendritic length and branches in rats treated with 4 and 6 ml kg−1 CeA for 6 weeks. Scale bar = 20 μm.
Apical Dendritic Intersections
Both the 4 and 6 ml kg−1 CeA groups showed a significant increase in dendritic intersections at the 80 μm (35.16 and 66.33% increases) and 100 μm (77.47 and 78.82% increases) concentric circles (80 μm concentric circle: 4.01 ± 0.47 in normal controls versus 5.42 ± 1.01 in the 4 ml kg−1 CeA group, P < 0.05, and 6.67 ± 0.73 in the 6 ml kg−1 CeA group, P < 0.001; 100 μm concentric circle: 3.73 ± 0.81 in normal controls versus 6.62 ± 1.07 in the 4 ml kg−1 CeA group, P < 0.01, and 6.67 ± 0.93 in the 6 ml kg−1 CeA group, P < 0.001). In addition, the 6 ml kg−1 CeA group showed a significantly increased number of dendritic intersections at the 60 μm concentric circle (53.08% increase) (3.24 ± 0.64 in normal controls versus 4.96 ± 1.35 in the 6 ml kg−1 CeA group, P < 0.05). There was no significant change in the dendritic intersections in any of the concentric circles in the 2 ml kg−1 CeA group when compared with the normal control group (Fig. 7).
Figure 7.
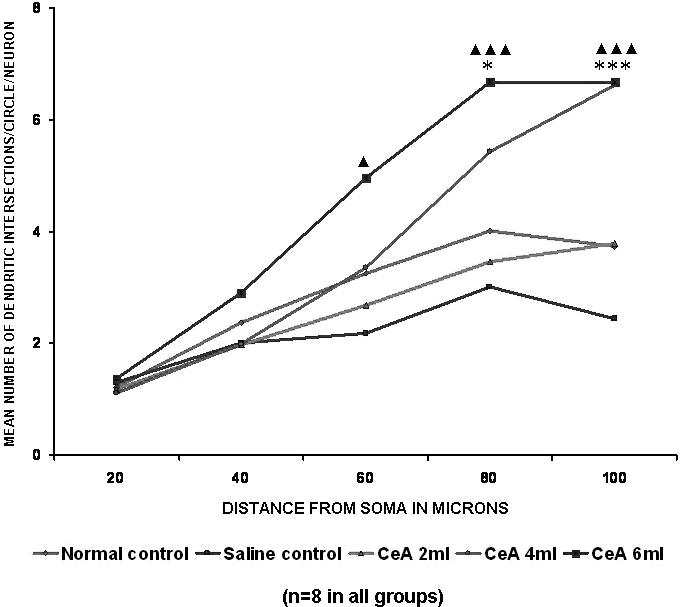
Apical dendritic intersections of hippocampal CA3 neurons in rats treated with 2, 4 and 6 ml kg−1 body weight of CeA for 4 weeks and in age-matched control and saline treated rats. Each point represents the mean of 8–10 neurons from each rat (SD not shown). F-value: 1.38, 3.23, 13.28, 40.8 and 39.64 at 20, 40, 60, 80 and 100 μm distance from the soma, respectively. Note that there is a significant increase in the dendritic intersections in rats treated with 4 and 6 ml kg−1 CeA. Normal control versus 4 ml kg−1 CeA: *P < 0.05, ***P < 0.001; normal control versus 6 ml kg−1 CeA: ▴P < 0.05, ▴▴▴P < 0.001 (ANOVA, Bonferroni's test).
Basal Dendritic Intersections
Both the 4 and 6 ml kg−1 CeA groups showed a significant increase in dendritic intersections at the 60 μm (67.75 and 101.09% increases) and 80 μm (161.20 and 133.01% increases) concentric circles (60 μm concentric circle: 6.42 ± 1.96 in normal controls versus 10.77 ± 0.68 in the 4 ml kg−1 CeA group, P < 0.01, and 12.91 ± 1.79 in the 6 ml kg−1 CeA group, P < 0.001; 80 μm concentric circle: 4.15 ± 1.94 in normal controls versus 10.84 ± 0.89 in the 4 ml kg−1 CeA group, P < 0.001, and 9.67 ± 2.19 in the 6 ml kg−1 CeA group, P < 0.01). In addition, the 6 ml kg−1 CeA group showed a significantly increased number of dendritic intersections in the 40 μm concentric circle [6.67 ± 0.9 in normal controls versus 10.7 ± 2.52 in the 6 ml kg−1 CeA group (60.0% increase), P < 0.01]. The 4 ml kg−1 CeA group showed a significantly increased number of dendritic intersections at the 100 μm concentric circle [1.91 ± 1.38 in normal controls versus 9.03 ± 1.69 in the 4 ml kg−1 CeA group (372.77% increase), P < 0.001]. There were no significant changes in the dendritic intersections at any of the concentric circles in the 2 ml kg−1 CeA group when compared with the normal control group (Fig. 8).
Figure 8.
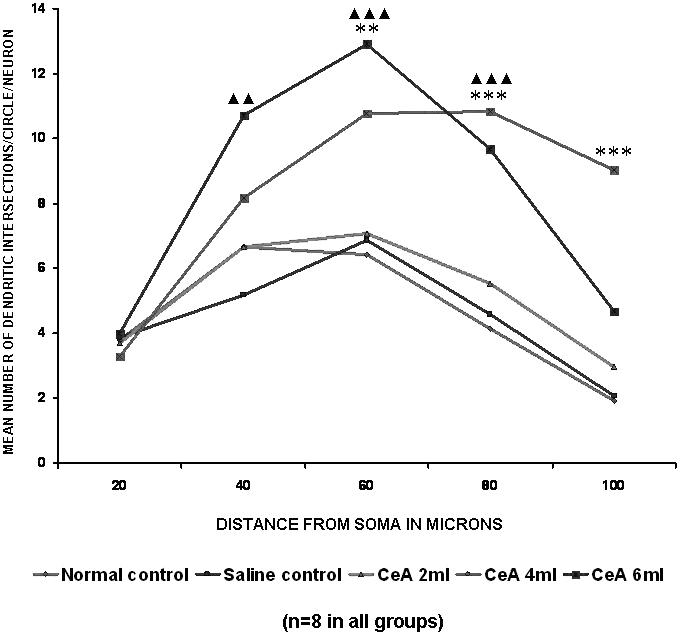
Basal dendritic intersections of hippocampal CA3 neurons in rats treated with 2, 4 and 6 ml kg−1 body weight of CeA for 4 weeks and in age-matched control and saline treated rats. Each point represents the mean of 8–10 neurons from each rat (SD not shown). F-value: 0.13, 21.05, 32.24, 27.22 and 24.78 at 20, 40, 60, 80 and 100 μm distance from the soma, respectively. Note that there is a significant increase in the dendritic intersections in rats treated with 4 and 6 ml kg−1 CeA. Normal control versus 4 ml kg−1 CeA: **P < 0.01, ***P < 0.001; normal control versus 6 ml kg−1 CeA: ▴▴P < 0.01, ▴▴▴P < 0.001 (ANOVA, Bonferroni's test).
Apical Dendritic Branching Points
The 4 and 6 ml kg−1 groups showed a significant increase in the apical dendritic branching points in the fourth (60–80 μm) zone [1.38 ± 0.24 in normal controls versus 2.3 ± 0.44 in the 4 ml kg−1 CeA group (66.66% increase), P < 0.05, and 2.97 ± 0.48 in the 6 ml kg−1 CeA group (115.21% increase), P < 0.001]. However, in the third (40–60 μm) zone only the 6 ml kg−1 CeA group showed a significantly increased number of dendritic branching points [1.21 ± 0.29 in normal controls versus 2.58 ± 0.52 in the 6 ml kg−1 CeA group (113.22% increase), P < 0.01]. In the fifth (80–100 μm) zone only the 4 ml kg−1 CeA group showed an increased number of dendritic branching points [0.96 ± 0.37 in normal controls versus 1.88 ± 0.53 in the 4 ml kg−1 CeA group (95.83% increase), P < 0.05]. No significant change was observed in the dendritic branching points in any of the concentric zones in the 2 ml kg−1 CeA group (Fig. 9).
Figure 9.
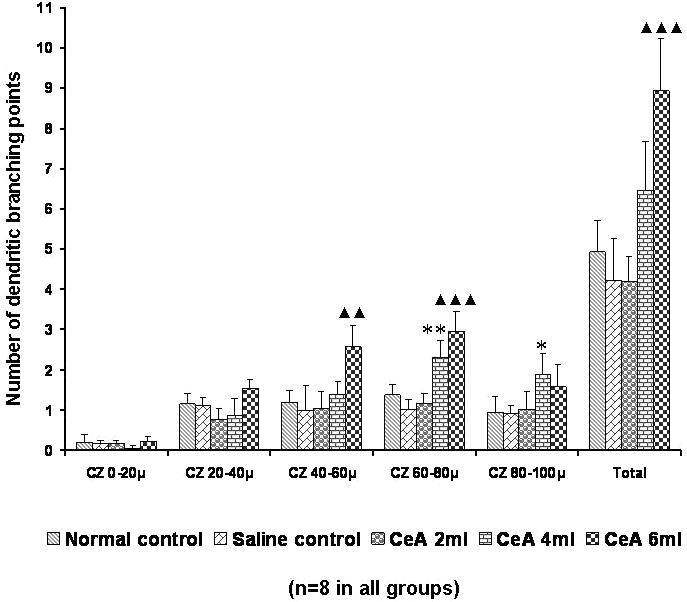
Apical dendritic branching points in the hippocampal CA3 neurons of rats treated with 2, 4 and 6 ml kg−1 body weight of CeA for 4 weeks and in age-matched control and saline treated rats at concentric zones (CZs) and total number of branching points. Each value represents the mean + SD of 8–10 neurons from each rat. F-value: 3.85, 9.04, 16.97, 45.88, 7.94 and 30.8 at the 0–20, 20–40, 40–60, 60–80 and 80–100 μm CZs and total number of branching points, respectively. Note the significant increase in dendritic branching points in 4 and 6 ml kg−1 CeA treated rats when compared with control rats. Normal control versus 4 ml kg−1 CeA: *P < 0.05, **P < 0.01; normal control versus 6 ml kg−1 CeA: ▴▴P < 0.01, ▴▴▴P < 0.001 (one-way ANOVA, Bonferroni's test).
Basal Dendritic Branching Points
Both the 4 and 6 ml kg−1 CeA groups showed a significant increase in dendritic branching points in the second (20–40 μm) zone [3.16 ± 0.82 in normal controls versus 4.46 ± 0.72 in the 4 ml kg−1 CeA group (41.13% increase), P < 0.05, and 6.36 ± 0.76 in the 6 ml kg−1 CeA group (101.26% increase), P < 0.001], the third (40–60 μm) zone [1.63 ± 0.56 in normal controls versus 3.37 ± 0.52 in the 4 ml kg−1 CeA group (106.74% increase), P < 0.01, and 4.14 ± 0.56 in the 6 ml kg−1 CeA group (153.98% increase), P < 0.001] and the fourth (60–80 μm) zone [0.34 ± 0.28 in normal controls versus 1.51 ± 0.35 in the 4 ml kg−1 CeA group (344.11% increase), P < 0.05, and 1.81 ± 0.86 in the 6 ml kg−1 CeA group (432.35% increase), P < 0.05]. No significant changes were observed in the dendritic branching points in any of the concentric zones in the 2 ml kg−1 CeA group (Fig. 10).
Figure 10.
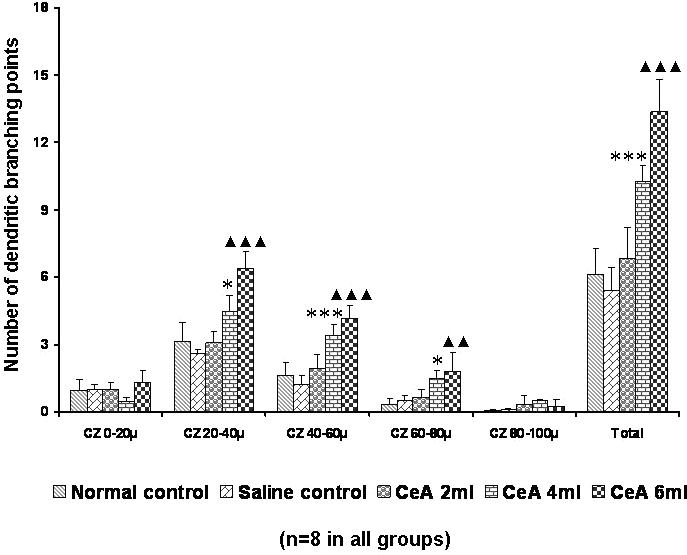
Basal dendritic branching points in the hippocampal CA3 neurons of rats treated with 2, 4 and 6 ml kg−1 body weight of CeA for 4 weeks and in age-matched control and saline treated rats at concentric zones (CZ) and total number of branching points. Each value represents the mean + SD of 8–10 neurons from each rat. F-value: 5.68, 44.84, 43.84, 15.44, 4.35 and 65.57 at the 0–20, 20–40, 40–60, 60–80 and 80–100 μm CZ and total number of branching points, respectively. Note the significant increase in dendritic branching points in 4 and 6 ml kg−1 CeA treated rats compared with control rats. Normal control versus 4 ml kg−1 CeA: *P < 0.05, ***P < 0.001; normal control versus 6 ml kg−1 CeA: ▴▴P < 0.01, ▴▴▴P < 0.001 (one-way ANOVA, Bonferroni's test).
Results of 2 Weeks' Treatment Group
There was no significant change in either apical or basal dendritic intersections or in branching points of hippocampal CA3 neurons in rats treated with CeA for 2 weeks in any of the dose groups (2, 4 and 6 ml kg−1; data not shown).
Discussion
CeA fresh leaf juice treatment at 4 ml kg−1 body weight for 4 and 6 weeks showed a 35–137% increase in apical dendritic intersections and a 64–372% increase in basal dendritic intersections in the outer concentric zones. The increase in the dendritic intersections of apical dendrites in rats treated with 6 ml kg−1 CeA for 4 and 6 weeks is higher than for the 4 ml kg−1 CeA group (53–780%). However, the increase in basal dendritic intersections in rats treated with 6 ml kg−1 CeA for 4 weeks was the same as that for the 4 ml kg−1 CeA group (60–331%). These changes were also seen in the outer concentric zones. The dendritic branching points showed an increase in number in outer zones in the 4 ml kg−1 CeA group (66–95%) and the 6 ml kg−1 CeA group (113–115%) in apical dendrites. Similar changes were also seen in basal dendrites.
There are several reports in the literature indicating dendritic modifications brought about by various factors leading to improved learning and memory. Repeated exposure to enriched environments has been shown to increase the density of spines and dendritic complexity in certain brain structures (22). After induction of long-lasting functional enhancement of synapses in the CA1 area, new spines appear on post-synaptic dendrites, whereas in control regions on the same dendrites or in slices where long-term potentiation is blocked, no significant spine growth occurs (23). Thus, dendritic structural reorganization is the key feature in learning and memory.
The memory-enhancing property of fresh leaf extracts of CeA in neonatal rats has been reported before (21). Nalini et al. (24) reported the memory-enhancing effect of aqueous extract of CeA in adult rats. In addition to its effect on behavior, asiatic acid, asiaticoside 6 and SM2, which are the active components of the plant, have a neuroprotective property. They show a strong inhibition of beta amyloid and free radical induced cell death in B103 cell cultures and hippocampal slices (25). The changes taking place in the dendritic morphology of hippocampal neurons reported in this article may be due to these neuroprotective active components and the key neuronal basis for improved learning and memory in these rats.
There have been reports where a few other herbal extracts have been proved to act on the central nervous system, thereby enhancing learning and memory. A recent study has shown that B. monnieri improves memory in humans (26–30). Clitoria ternatea and jatamansi have also been reported to be excellent memory enhancers (28,31). Clitoria ternatea has been shown to enhance acetylcholine content in the rat hippocampus (32).
The dendrites of hippocampal CA3 neurons receive extrinsic and intrinsic inputs from the different parts of the limbic system, which is concerned with learning and memory. The extrinsic afferents are mainly from the entorhinal cortex, septal area, mamillary body and nuclei of the brain stem (33,34). The CA3 neurons also receive a significant number of intrinsic afferents from the axons of dentate granule cells and the contralateral CA3 regions (35,36). Alterations such as increases or decreases in dendritic length and increases or decreases in dendritic branches in these neurons may result in alterations in learning and memory behavior.
The increased dendritic intersections and branches of hippocampal CA3 neurons in animals treated with 4 and 6 ml kg−1 of CeA per day for 4 and 6 weeks in the present study suggest that such doses of plant extract were adequate to induce structural changes in these neurons. Naturally, such changes will have a profound effect on behavior because of the additional dendrites that are available on these neurons for the formation of new synapses. It can be noted from the results that a significant number of additional dendrites are formed closer to the soma of neurons, i.e. within 60 μm. This results in more rapid and effective conduction of impulses, which may be one of the reasons for the enhanced learning and memory in these animals reported earlier (21). Such an increase in dendrites during the neonatal period has been shown to be effective in enhancing learning and memory (37).
Thus, from the present study it is evident that administration of fresh leaf extracts of CeA during the growth spurt (neonatal) period facilitates dendritic growth in the hippocampal CA3 neurons. Increased dendritic arborization may be the neuronal basis for the improved learning and memory reported earlier (21).
REFERENCES
- 1.Sharma PV. Dravyaguna Vignana. 13th edition. New Delhi, India: Chaukhamba Publications Vishwa Bharati Academy; 1992. pp. 3–5. [Google Scholar]
- 2.Sivarajan VV. Ayurvedic Drugs and Their Plant Sources. New Delhi, India: Oxford and IBH Publishing Company; 1994. Indira Balachandran; p. 97. 289, 290. [Google Scholar]
- 3.Dash PK, Mistry IU, Rao AR, Patel KS. Role of medhya rasayana in school children. Ayu. 1996;12:15. [Google Scholar]
- 4.Satyavati GV, Gupta AK, Tandon N. Medicinal Plants of India. 1st edition. New Delhi: Indian Council of Medical Research; 1976. pp. 18–21.pp. 216–20. [Google Scholar]
- 5.Anbuganapathi GA. Synergetic effect of vallarai and brahmi on learning ability of albino mice and school children. Paper presented at the International Seminar on Recent Trends in Pharmaceutical Sciences; Ootacamund. February 1995. pp. 18–20. [Google Scholar]
- 6.Rajagopalan V. Effect of Ayushman-8 in manasa mandata (mental retardation). Paper presented at the Seminar on Research in Ayurveda and Siddha; New Delhi. CCRAS; March 1995. pp. 20–2. [Google Scholar]
- 7.Shah LP. An open clinical trial of mentat in hyperkinetic children. Probe. 1992;31:125–9. [Google Scholar]
- 8.Appa Rao MVR, Srinivasan K, Rao KT. The effect of mandookaparni (Centella asiatica) on the general mental ability (medhya) of mentally retarded children. J Res Indian Med. 1973;8:9–16. [Google Scholar]
- 9.Joshi H, Parle M. Brahmi rasayana improves learning and memory in mice. Evid Based Complement Alternat Med. 2006;3:79–85. doi: 10.1093/ecam/nek014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Galani R, Weiss I, Cassel JC, Kelche C. Spatial memory habituation and reactions to spatial and nonspatial changes in rats with selective lesions of the hippocampus, the entorhinal cortex or the subiculum. Behav Brain Res. 1998;96:1–12. doi: 10.1016/s0166-4328(97)00197-6. [DOI] [PubMed] [Google Scholar]
- 11.Batch ME, Hawkins RQ, Osman M, Kandel ER, Mayfond M. Impairment of spatial but not contextual memory in Ca MKII mutant mice with a selective loss of hippocampal LTP in the range of theta sequency. Cell. 1995;81:905–15. doi: 10.1016/0092-8674(95)90010-1. [DOI] [PubMed] [Google Scholar]
- 12.Kesner RP, Hopkins RD. Short-term memory for duration and distance in humans: role of the hippocampus. Neuropsychology. 2001;15:58–68. doi: 10.1037//0894-4105.15.1.58. [DOI] [PubMed] [Google Scholar]
- 13.Jensen O, Lisman JE. Hippocampal CA3 region predicts memory sequences accounting for the phase precession of place cells. Learn Mem. 1996;3:279–87. doi: 10.1101/lm.3.2-3.279. [DOI] [PubMed] [Google Scholar]
- 14.Lassale JM, Bataille T, Halley H. Reversible inactivation of the hippocampal mossy fiber synapses in mice impairs spatial learning, but neither consolidation nor memory retrieval, in the Morris navigation task. Neurobiol Learn Mem. 2000;73:243–57. doi: 10.1006/nlme.1999.3931. [DOI] [PubMed] [Google Scholar]
- 15.Christian EP, Deadwyler SA. Behavioral functions and hippocampal cell types: evidence for 2 non-over lapping populations in the rat. J Neurophysiol. 1986;55:331–48. doi: 10.1152/jn.1986.55.2.331. [DOI] [PubMed] [Google Scholar]
- 16.Stubley Weatherly L, Harding JW, Wright JW. Effect of discrete kainic acid induced hippocampal lesions on spatial and contextual learning in rats. Brain Res. 1996;716:29–38. doi: 10.1016/0006-8993(95)01589-2. [DOI] [PubMed] [Google Scholar]
- 17.Dobbing J, Sands J. Comparative aspects of the brain growth spurt. Early Human Dev. 1979;3:79–83. doi: 10.1016/0378-3782(79)90022-7. [DOI] [PubMed] [Google Scholar]
- 18.Soumyanath A, Zhong YP, Gold SA, Yu X, Koop DR, Bourdette D, et al. Centella asiatica accelerates nerve regeneration upon oral administration and contains multiple active fractions increasing neurite elongation in-vitro. J Pharm Pharmacol. 2005;57:1221–9. doi: 10.1211/jpp.57.9.0018. [DOI] [PubMed] [Google Scholar]
- 19.Rao BSS, Desiraju T, Raju TR. Neuronal plasticity induced by self stimulation rewarding experiences in rats—a study on alteration in dendritic branching in pyramidal neurons of hippocampus and motor cortex. Brain Res. 1993;627:216–24. doi: 10.1016/0006-8993(93)90324-g. [DOI] [PubMed] [Google Scholar]
- 20.Sholl DA. The Organization of the Cerebral Cortex. London: Methun; 1956. [Google Scholar]
- 21.Rao Mohandas KG, Rao Muddanna S, Rao Gurumadhva S. Centella asiatica (linn) induced behavioural changes during growth spurt period in neonatal rats. Neuroanatomy. 2005;4:18–23. [Google Scholar]
- 22.Moser MB. Making more synapses: a way to store information. Cell Mol Life Sci. 1999;55:593–600. doi: 10.1007/s000180050317. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Engert F, Bonhoeffer T. Dendritic spine changes associated with hippocampal long-term synaptic plasticity. Nature. 1999;399:66–70. doi: 10.1038/19978. [DOI] [PubMed] [Google Scholar]
- 24.Nalini K, Aroor AR, Karanth KS, Rao A. Effect of Centella aciatica fresh leaf extract on learning and memory and biogenic amine turnover in albino rats. Fitoterapia. 1992;63:232–8. [Google Scholar]
- 25.Mook Jung I, Shin JE, Yun SH, Huh K, Koh JY, Park HK, et al. Protective effects of asiaticoside derivatives against beta amyloid neurotoxicity. J Neurosci Res. 1999;58:417–25. [PubMed] [Google Scholar]
- 26.Roodenrys S, Booth D, Bulzomi S, Phipps A, Micallef C, Smoker J. Chronic effects of Brahmi (Bacopa monnieri) on human memory. Neuropsychopharmacology. 2002;27:279–81. doi: 10.1016/S0893-133X(01)00419-5. [DOI] [PubMed] [Google Scholar]
- 27.Stough C, Lloyd J, Clarke J, Downey LA, Hutchison CW, Rodgers T, et al. The chronic effects of an extract of Bacopa monnieri on cognitive function in healthy human subjects. Psychopharmacology. 2001;156:481–4. doi: 10.1007/s002130100815. [DOI] [PubMed] [Google Scholar]
- 28.Indurwade NH, Biyani KR. Evaluation of comparative and combined depressive effect of Brahmi, Shankapushpi and Jatamansi in mice. Indian J Med Sci. 2000;54:339–41. [PubMed] [Google Scholar]
- 29.Shukia B, Khanna NK, Godhwani JL. Effect of Brahmi rasayan on central nervous system. J Ethnopharmacol. 1987;21:65–74. doi: 10.1016/0378-8741(87)90095-x. [DOI] [PubMed] [Google Scholar]
- 30.Singh HK, Dhawan BN. Effect of Bacopa monnieri (Linn) extract on avoidance responses in rat. J Ethnopharmacol. 1982;5:205–14. doi: 10.1016/0378-8741(82)90044-7. [DOI] [PubMed] [Google Scholar]
- 31.Rai KS, Murthy KD, Karanth KS, Rao MS. Clitoria ternatea root extract treatment during growth spurt period enhances learning and memory in rats. Indian J Physiol Pharmacol. 2001;45:305–13. [PubMed] [Google Scholar]
- 32.Rai KS, Murthy KD, Karanth KS, Nalili K, Rao MS, Srinivasan KK. Clitoria ternatea root extract enhances acetylcholine content in rat hippocampus. Fitoterapia. 2002;73:685–9. doi: 10.1016/s0367-326x(02)00249-6. [DOI] [PubMed] [Google Scholar]
- 33.Van Hoesen GW, Pandya DN. Some connections of the entorhinal cortex (area 28) and perirhinal (area 35) cortices of rhesus monkey, III Efferent connections. Brain Res. 1975;95:39–59. doi: 10.1016/0006-8993(75)90206-1. [DOI] [PubMed] [Google Scholar]
- 34.Raisman G, Cowan WM, Powell TPS. The extrinsic afferent commissural and association fibers of the hippocampus. Brain. 1965;88:963–96. [Google Scholar]
- 35.Hamlyn LH. The fine structure of the mossy fiber endings in hippocampus of the rabbit. J Anat. 1962;96:112–20. [PMC free article] [PubMed] [Google Scholar]
- 36.Amaral DG, Witter MP. The three dimensional organization of the hippocampal formation. A review of anatomical data. Neuroscience. 1989;31:571–91. doi: 10.1016/0306-4522(89)90424-7. [DOI] [PubMed] [Google Scholar]
- 37.Tang AC. Neonatal exposure to novel environment enhances hippocampal dependent memory function during infancy and adulthood. Learn Mem. 2001;8:257–64. doi: 10.1101/lm.43101. [DOI] [PMC free article] [PubMed] [Google Scholar]