

Abstract
Agrobacterium tumefaciens genetically transforms plant cells by transferring a portion of the bacterial Ti-plasmid, the T-DNA, to the plant and integrating the T-DNA into the plant genome. Little is known about the T-DNA integration process, and no plant genes involved in integration have yet been identified. We characterized an Arabidopsis mutant generated by T-DNA insertional mutagenesis, rat5, that is resistant to Agrobacterium root transformation. rat5 contains two copies of T-DNA integrated as a tandem direct repeat into the 3′ untranslated region of a histone H2A gene, upstream of the polyadenylation signal sequence. Transient and stable β-glucuronidase expression data and assessment of the amount of T-DNA integrated into the genomes of wild-type and rat5 Arabidopsis plants indicated that the rat5 mutant is deficient in T-DNA integration. We complemented the rat5 mutation by expressing the RAT5 histone H2A gene in the mutant plant. Overexpression of RAT5 in wild-type plants increased Agrobacterium transformation efficiency. Furthermore, transient expression of a RAT5 gene from the incoming T-DNA was sufficient to complement the rat5 mutant and to increase the transformation efficiency of wild-type Arabidopsis plants.
Keywords: T-DNA transformation, haplo-insufficiency
Agrobacterium tumefaciens is a Gram-negative soil bacterium that has been exploited by plant biologists to introduce foreign DNA into plants. Although best known for this practical application, the actual mechanism of DNA transfer from bacteria to plants is not completely understood. The biological processes that take place within the bacteria during and before DNA transfer have been partially characterized. The DNA that is transferred from Agrobacterium to the plant cell is a segment of the Ti, or tumor-inducing, plasmid called the T-DNA (transferred DNA). Virulence (vir) genes responsible for T-DNA processing and transfer lie elsewhere on the Ti plasmid (1, 2). The role of vir genes in T-DNA processing, the formation of bacterial channels for export of T-DNA, and the attachment of bacteria to the plant cell are reasonably well understood. In contrast, not much is known about the role of plant factors in T-DNA transfer and integration within the plant cell. Several plant factors that may be involved in these processes, a karyopherin-α, a cyclophilin, and a type 2C protein phosphatase, recently have been identified (refs. 3 and 4; Y. Tao, P. Rao, and S.B.G., unpublished work). Other evidence for the involvement of plant factors in T-DNA transfer and integration comes from our identification of several ecotypes and mutants of Arabidopsis that are resistant to Agrobacterium transformation (5, 6).
We recently identified several T-DNA-tagged mutants of Arabidopsis that are highly recalcitrant to Agrobacterium root transformation (6). We called these rat mutants (resistant to Agrobacterium transformation). In most of these mutants Agrobacterium transformation is blocked at an early step, either during bacterial attachment to the plant cell or before T-DNA nuclear import. In some of the mutants, however, the T-DNA integration step is most likely blocked. T-DNA does not encode enzymes necessary for DNA integration. The only Agrobacterium proteins that may play a role in T-DNA integration are VirD2 and VirE2 that associate with the T-DNA. VirD2 covalently attaches to the 5′ end of the T-strand (7–9) and is believed to play a role in T-DNA integration (10, 11). Plant proteins may interact with VirD2 to help integrate the T-DNA (11). The role of VirE2 protein in T-DNA integration is not clear. VirE2, a single-stranded DNA binding protein, presumably coats the T-strand and prevents nucleolytic degradation (12–14). Integrated T-DNA delivered from VirE2 mutant Agrobacterium cells is often severely truncated at the 3′ end (15). T-DNA integration does not take place by homologous recombination, the most common method of foreign DNA integration in prokaryotes and lower eukaryotes, because no extensive homology between the T-DNA and target sequences has been found. T-DNA therefore integrates by illegitimate recombination (16–19), the predominant mechanism of DNA integration into the genomes of higher plants (20–22). However, plant factors involved in illegitimate recombination of T-DNA into the plant genome have not yet been identified.
Here we report the characterization of a T-DNA tagged Arabidopsis mutant, rat5, that is recalcitrant to Agrobacterium root transformation. We show that in rat5 a histone H2A gene is disrupted. Complementation analysis and RAT5 overexpression indicate that histone H2A plays a role in Agrobacterium transformation. We also show that the T-DNA integration step of transformation is blocked in the rat5 mutant. We hypothesize that histone H2A (RAT5) plays an important role in illegitimate recombination of T-DNA into the plant genome.
Materials and Methods
Nucleic Acid Manipulation.
Total plant genomic DNA was isolated according to Dellaporta et al. (23). Restriction endonuclease digestions, agarose gel electrophoresis, plasmid isolation, and DNA blot analysis were conducted as described (24).
Plasmid Rescue.
Genomic DNA (5 μg) of rat5 was digested to completion with SalI. The digested DNA was extracted with phenol/chloroform and precipitated with ethanol. The DNA was self-ligated in a final volume of 500 μl in 1× ligation buffer (Promega) with 3 units of T4 DNA ligase at 16°C for 16 hr. The ligation mixture was precipitated with ethanol, transformed into electrocompetent Escherichia coli DH10B cells (mcrBC−; Life Technologies, Gaithersburg, MD) by electroporation (25 μF, 200 Ω, and 2.5 kV) and plated on LB medium containing ampicillin (100 μg/ml). Colonies were lifted onto a nylon membrane, the bacteria were lysed, and DNA was denatured in situ (24). A radio-labeled left border sequence (3.0-kbp EcoRI fragment of pE1461) was used as a hybridization probe to identify a plasmid containing the left border. By restriction fragment analysis a plasmid containing both the left border and plant junction DNA was identified. The plant junction fragment was confirmed by hybridizing the junction fragment to wild-type plant DNA. A restriction map of this plasmid, containing the left border-plant junction DNA, was made. A 1.7-kbp EcoRI fragment that contained plant DNA plus 75 bp of the left border sequence was subcloned into pBluescript, resulting in pE1509, and sequenced at the Purdue University sequencing center.
Growth of Agrobacterium and in Vitro Root Inoculation of Arabidopsis thaliana.
These were performed as described (6).
Construction of the Binary Vectors pKM4 and pKM5.
The plasmid pE1509 containing the 1.7-kbp junction fragment was digested with EcoRI to release the junction fragment. The 5′ overhanging ends were filled in by using the Klenow fragment of DNA polymerase I and deoxynucleotide triphosphates. The T-DNA binary vector (pE1011) pGTV-HPT (25) was digested with the enzymes SacI and SmaI, releasing the promoterless gusA gene from pGTV-HPT. The 3′ overhanging sequence of the larger fragment containing the origin of replication and the hygromycin resistance gene (hpt) was removed by using the 3′–5′ exonuclease activity of Klenow DNA polymerase, and the resulting 1.7-kbp blunt end fragment was ligated to the blunt ends of the binary vector. The resulting plasmid containing the 1.7-kbp fragment in the correct orientation (pAnos downstream of the histone H2A gene) was named pKM4 (strain E1547).
An approximately 9-kbp wild-type genomic SacI fragment containing the histone H2A gene (RAT5) from a lambda genomic clone was cloned into the SacI site of pBluescript. This 9-kbp SacI fragment subsequently was released from pBluescript by digestion with SacI and was cloned into the SacI site of the binary vector pGTV-HPT, resulting in the plasmid pKM5 (strain E1596). Both pKM4 and pKM5 were transferred by triparental mating (26) into the nontumorigenic Agrobacterium strain GV3101 (27), resulting in the strains A. tumefaciens At1012 and At1062, respectively.
Germ-Line Transformation of Arabidopsis.
Germ-line transformations were performed as described (28). Transgenic plants were selected on B5 medium containing 20 μg/ml hygromycin.
Results
Characterization of the rat5 Mutant.
rat5, an Arabidopsis T-DNA tagged mutant, previously was identified as resistant to Agrobacterium root transformation (6). An in vitro root inoculation assay was performed by using the wild-type Agrobacterium strain A208 (At10). After 1 month, the percentage of root bundles that formed tumors was calculated. Greater than 90% of the root bundles of the wild-type plants (ecotype Ws) formed large green teratomas. In contrast, fewer than 10% of the root bundles from the rat5 plants responded to infection, forming small yellow calli (Fig. 1A). A homozygous rat5 plant (pollen donor) was crossed to a wild-type plant (egg donor), and the resulting F1 progeny were tested for susceptibility to Agrobacterium transformation. This analysis indicated that rat5 appears dominant, although further analysis (see below) indicates that rat5 is haplo-insufficient (ref. 6; Fig. 1A). F2 progeny analysis indicated that kanamycin resistance segregated 3:1, indicating that a single locus had been disrupted by the mutagenizing T-DNA. Earlier, we showed that kanamycin resistance cosegregated with the rat5 phenotype, indicating that a gene involved in Agrobacterium transformation had likely been mutated by the T-DNA insertion (6).
Figure 1.
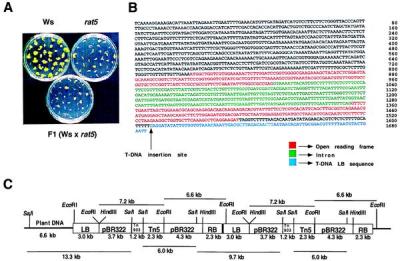
Characterization of the rat5 mutant. (A) Stable transformation of wild-type Arabidopsis ecotype Ws, the rat5 mutant, and the F1 progeny. Sterile root segments were infected with A. tumefaciens A208. Two days after cocultivation, the roots were transferred to Murishige and Skoog medium lacking phytohormones and containing timentin. Tumors were scored after 4 weeks. (B) Sequence of the rat5/T-DNA junction region. (C) Pattern of T-DNA integration in rat5. LB, T-DNA left border; RB, T-DNA right border; pBR322, pBR322 sequences containing the β-lactamase gene and ColE1 origin of replication; Tn903, kanamycin resistance gene for E. coli selection; Tn5, kanamycin resistance gene for plant selection. Five micrograms of genomic DNA from the rat5 mutant was digested with either EcoRI or SalI and was blotted onto a nylon membrane. An EcoRI–SalI fragment of pBR322 was used as the hybridization probe. Restriction fragment sizes shown above the T-DNA were detected by EcoRI digestion and the sizes shown below the T-DNA were detected by SalI digestion.
Recovery of a T-DNA-Plant Junction from rat5.
The T-DNA integration pattern in the rat5 mutant was determined by DNA blot analyses. There are two copies of T-DNA integrated as a direct tandem repeat into the genome of the rat5 mutant (Fig. 1C).
A left border T-DNA-plant junction was recovered from rat5 by using a plasmid rescue technique, and a restriction endonuclease map was constructed. An approximately 1.7-kbp EcoRI fragment containing both plant and left border DNA was subcloned into pBluescript and sequenced (Fig. 1B). Analysis of this junction region indicated that the T-DNA had inserted into the 3′ untranslated region (UTR) of a histone H2A gene (Fig. 1B). We further characterized the histone H2A genes of Arabidopsis by isolating and sequencing numerous cDNA and genomic clones (K.S.M., H. Yi, and S.B.G., unpublished work). Six different gene variants of histone H2A were identified. We identified a lambda genomic clone containing the wild-type histone H2A gene corresponding to RAT5. DNA sequence analysis of this genomic clone indicated that in rat5 the T-DNA had inserted upstream of the consensus polyadenylation signal (AATAA). DNA blot analysis of Ws and rat5 DNA indicated that the T-DNA insertion in rat5 did not cause any major rearrangements in the plant DNA immediately around the site of insertion (data not shown). However, these experiments cannot rule out the existence of rearrangements many kilobases from the T-DNA insertion site. Such rearrangements resulting from Agrobacterium-mediated transformation have been reported (29). We hypothesize that disruption of the 3′ UTR of the RAT5 histone H2A gene is the sole cause for the rat phenotype in the rat5 mutant.
Complementation of the rat5 Mutant with a Histone H2A Gene (RAT5).
Two different constructions were made to perform complementation analyses of the rat5 mutant. First, a nopaline synthase (NOS) terminator (3′ NOS) was fused to the 3′ region of the 1.7-kbp junction fragment (the sequence of this 1.7-kbp fragment is shown in Fig. 1B). This construction contains the RAT5 histone H2A gene with its own promoter and a 3′ NOS. This fragment was cloned into the plasmid pGTV-HPT (25) containing a hygromycin resistance gene between the left and the right T-DNA borders, resulting in pKM4 (Fig. 2A). For the second construction, a 9-kbp SacI genomic fragment of wild-type Ws DNA containing a RAT5 histone H2A gene plus at least 2-kbp sequences upstream and downstream of RAT5 was cloned into pGTV-HPT, resulting in pKM5 (Fig. 2B). pKM4 and pKM5 were transferred into the nontumorigenic Agrobacterium strain GV3101, resulting in strains A. tumefaciens At1012 and At1062, respectively.
Figure 2.
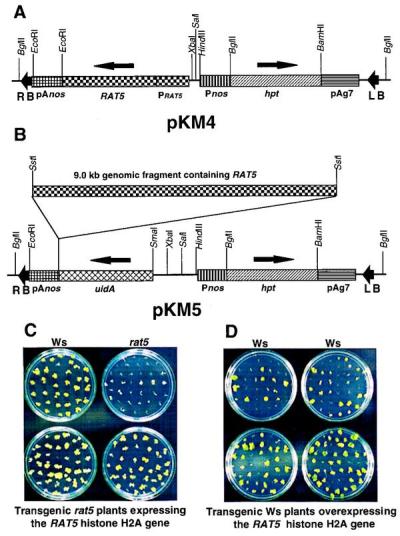
Complementation of the rat5 mutant and overexpression of RAT5 in wild-type Arabidopsis plants. Maps of the binary vectors pKM4 (A) and pKM5 (B). RB, T-DNA right border; LB, T-DNA left border; pAnos, NOS polyadenylation signal sequence; RAT5, coding sequence of the RAT5 histone H2A gene; PRAT5, promoter sequence of the RAT5 histone H2A gene; Pnos, NOS promoter; hpt, hygromycin resistance gene; pAg7, agropine synthase polyadenylation signal sequence; uidA, promoterless gusA gene. Arrows above the RAT5, uidA, and hpt genes indicate the direction of transcription. (C) Complementation of the rat5 mutant. rat5 mutant plants were transformed with an Agrobacterium strain containing the binary vector pKM4 (At1012). Hygromycin-resistant transgenic plants were obtained and were self-pollinated to obtain T2 plants. Sterile root segments of T2 plants expressing RAT5, wild-type Ws plants, and rat5 mutant plants were infected with the tumorigenic strain A. tumefaciens A208. Two days after cocultivation, the roots were moved to Murishige and Skoog medium lacking phytohormones and containing timentin. Tumors were scored after 4 weeks. (D) Tumorigenesis assay of Ws transgenic plants overexpressing the RAT5 histone H2A gene. Ws plants were transformed with A. tumefaciens At1012 containing the binary vector pKM4. Hygromycin-resistant transgenic plants were obtained and were self-pollinated to obtain T2 plants. Sterile root segments of T2 plants overexpressing RAT5 and wild-type Ws plants were infected at low bacterial density with A. tumefaciens A208. After 2 days cocultivation, the roots were moved to MS medium lacking phytohormones and containing timentin. Tumors were scored after 4 weeks.
Strains At1012 and At1062 were used to transform rat5 plants by using a germ-line transformation method (28), and transgenic rat5 plants were selected for resistance to hygromicin. These transgenic plants were allowed to self-fertilize and T1 seeds were collected. Six transgenic lines obtained by transformation with At1012 (the histone H2A with 3′ NOS) were selected and their seeds were germinated in the presence of hygromycin. Tumorigenesis assays were performed as described (6) on at least five different plants from each of the six transgenic lines. The results indicated that in five of the six transgenic rat5 lines tested, the tumorigenesis-susceptibility phenotype was recovered (Fig. 2C; Table 1). Teratomas incited on the roots of these plants appeared similar to tumors generated on a wild-type plant. One of the transgenic plants tested did not recover the tumorigenesis-susceptibility phenotype, probably because of an inactive transgene. Transgenic T1 plants of rat5 obtained by transformation with At1062 (containing a gene RAT5 from the wild-type plant) also were tested for restoration of the tumorigenesis-susceptibility phenotype. Some of these plants also were able to recover the tumorigenesis-susceptibility phenotype, indicating complementation of the rat5 mutation (data not shown). Hygromycin-resistant transgenic plants generated by transforming the rat5 mutant with pGPTV-HPT alone did not form tumors upon infection with A. tumefaciens A208 (data not shown).
Table 1.
Complementation of the rat5 mutant and overexpression of RAT5 in wild-type (Ws) Arabidopsis plants
| Line | % Root bundles with tumors | Tumor morphology |
|---|---|---|
| rat5 complementation with At1012 (T2 plants) | ||
| Ws | 98 ± 2 | Large, green |
| rat5 | 21 ± 6 | Small, yellow |
| rat5 At1012-1 | 64 ± 30 | Large + small, green |
| rat5 At1012-2 | 17 ± 4 | Small, yellow |
| rat5 At1012-3 | 70 ± 20 | Large + medium, green |
| rat5 At1012-4 | 86 ± 6 | Large, green |
| rat5 At1012-5 | 82 ± 10 | Large, green |
| rat5 At1012-6 | 92 ± 5 | Large, green |
| Overexpression of RAT5 in Ws (T2 plants)* | ||
| Ws | 35 ± 14 | Large, green |
| Ws At1012-1 | 69 ± 27 | Large, green |
| Ws At1012-2 | 68 ± 25 | Large, green |
| Ws At1012-3 | 64 ± 13 | Large, green |
| Ws At1012-4 | 63 ± 20 | Large, green |
At least five plants were tested for each mutant, and 40–50 root bundles were tested for each plant.
Agrobacterium was diluted to a concentration 100-fold lower than that normally used, and single root segments were separated.
To confirm the genetic basis of the complementation experiment, a cosegregation analysis was performed on one of the rat5 transgenic lines (rat5 At1012–6) obtained by transformation of the rat5 mutant with A. tumefaciens At1012. To examine the cosegregation of the complementing T-DNA containing the RAT5 gene with the tumorigenesis-susceptibility phenotype, seeds from a T2 plant homozygous for the rat5 mutation but heterozygous for hygromycin resistance were germinated and grown on B5 medium without selection. Roots of these plants subsequently were tested for hygromycin resistance and susceptibility to crown gall tumorigenesis. All plants that were sensitive to hygromycin were also resistant to tumor formation in a manner similar to that of the rat5 mutant. Of the 25 hygromycin-resistant plants, at least eight were susceptible to tumorigenesis. However, 17 hygromycin-resistant plants remained recalcitrant to Agrobacterium-mediated transformation. It is likely that these plants are heterozygous with respect to the complementing RAT5 gene and did not express this gene to a level high enough to restore susceptibility to tumorigenesis. This possibility corresponds to our finding that the rat5 mutation is haplo-insufficient, and that therefore one active copy of RAT5 is not sufficient to permit Agrobacterium-mediated transformation. Our molecular and genetic data strongly indicate that in the rat5 mutant disruption of a histone H2A gene is responsible for the tumorigenesis-deficiency (rat) phenotype.
The rat5 Mutant Is Haplo-Insufficient.
Our ability to complement the rat5 mutant suggested that rat5 is haplo-insufficient rather than dominant. To test this, we transformed wild-type Ws plants with A. tumefaciens GV3101(pE1553) and selected six independent T2 plants for analysis. pE1553 is identical to pKM4 except that the nos poly(A) signal was deleted by digestion of pKM4 with EcoRI. pE1553 thus contains the mutant rat5 gene with 75 bp of T-DNA inserted 46 nt after the stop codon. Some 98.1 ± 1% of the root segments from control Ws plants showed teratomas. Root segments from the six transgenic plants gave a similarly high infection frequency (data not shown). Thus, the mutant rat5 gene does not confer a dominant rat phenotype upon wild-type plants, and we conclude that rat5 is haplo-insufficient rather than dominant.
Overexpression of a Histone H2A (RAT5) Gene in Wild-Type Plants Improves the Efficiency of Agrobacterium Transformation.
To determine further whether the RAT5 gene plays a role in Agrobacterium-mediated transformation, we used A. tumefaciens At1012 to generate several transgenic Arabidopsis plants containing additional copies of the RAT5 histone H2A gene. These transgenic plants were self-pollinated, T1 seeds were collected, and T2 plants were germinated in the presence of hygromycin. Tumorigenesis assays were performed on at least five plants from each of four different transgenic lines. Because ecotype Ws is highly susceptible to Agrobacterium transformation, we altered the tumorigenesis assay to detect subtle differences between the transformation-susceptible wild-type plant and transgenic wild-type plants overexpressing RAT5. These alterations included inoculation of root segments with a 100-fold lower concentration (2 × 107 colony-forming units/ml) of bacteria than that normally used (2 × 109 colony-forming units/ml), and spreading individual root segments rather than bundles of root segments on Murishige and Skoog medium to observe tumor production. The results, shown in Table 1 and Fig. 2D, indicate that transgenic plants overexpressing RAT5 are approximately twice as susceptible to root transformation as are wild-type Ws plants. These data indicate that overexpression of RAT5 can increase susceptibility to transformation.
Transient Expression of Histone H2A Is Sufficient to Permit Transformation of rat5 and to Increase the Transformation Efficiency of Wild-Type Ws Plants.
We tested whether expression of the RAT5 histone H2A gene from the incoming T-DNA would complement the rat5 mutant. Although transformation of this mutant with an Agrobacterium strain harboring pGPTV-HYG (lacking a histone H2A gene) resulted in only a few, slow-growing calli on hygromycin selection medium, Agrobacterium strains harboring pKM4 or pKM5 incited rapidly growing hygromycin-resistant calli on 60 ± 21% and 54 ± 22% of the rat5 root segment bundles, respectively. In addition, when wild-type plants were infected (at low bacterial density) with a tumorigenic Agrobacterium strain (A208) harboring pKM4, 78 ± 8% of the root segments developed tumors, compared with 36 ± 9% of the root segments infected with a tumorigenic bacterial strain harboring pGPTV-HYG. These transformation experiments indicate that Agrobacterium strains containing the binary vectors pKM4 or pKM5 are able to transform rat5 mutant plants at relatively high efficiency, and on wild-type plants are 2-fold more tumorigenic, and are better able to incite hygromycin-resistant calli, than are Agrobacterium strains containing the “empty” binary vector pGPTV-HYG. We speculate that transiently produced histone H2A can improve the stable transformation efficiency of plants by Agrobacterium.
The rat5 Mutant Is Deficient in T-DNA Integration.
We previously showed that Agrobacterium-mediated transformation of the Arabidopsis rat5 mutant results in a high efficiency of transient transformation but a low efficiency of stable transformation (6). This result suggested that rat5 is likely deficient in T-DNA integration. We therefore tested this hypothesis directly. Root segments from Ws and rat5 plants were inoculated with A. tumefaciens GV3101 harboring the T-DNA binary vector pBISN1. pBISN1 contains a gusA-intron gene under the control of a “super-promoter” (30, 31). Two days after cocultivation, the root segments were transferred to callus-inducing medium containing timentin (100 μg/ml) to kill the bacteria. Three days after infection, a few segments were stained for β-glucuronidase (GUS) activity by using the chromogenic dye X-gluc. As reported previously (6), both the wild-type and the rat5 mutant showed high levels of GUS expression (approximately 90% of the root segments stained blue; Fig. 3A). The remaining root segments were allowed to form calli on callus-inducing medium containing timentin to kill Agrobacterium, but lacking any antibiotic for selection of plant transformation. After 4 weeks, numerous calli derived from at least five different Ws and rat5 plants were stained with X-gluc. Of the Ws calli sampled, 92 ± 12% showed large blue staining areas, whereas only 26 ± 10% of the rat5 calli showed GUS activity, and most of these blue staining regions were small (Fig. 3A). These data indicate that although the rat5 mutant can transiently express the gusA gene at high levels, it fails to stabilize gusA expression.
Figure 3.
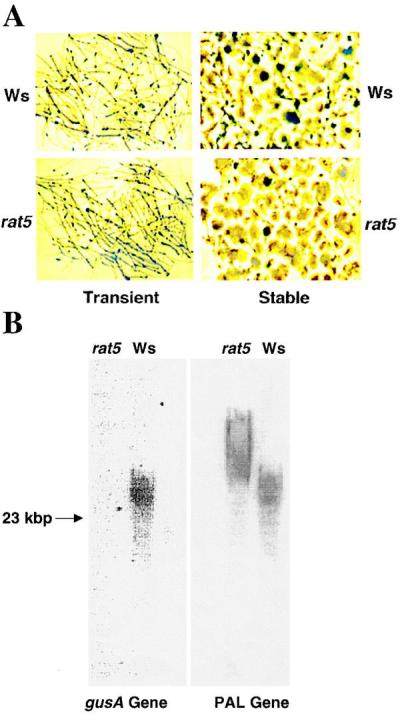
T-DNA integration assays of rat5 and Ws plants. (A) Transient and stable GUS expression in Ws and rat5. Sterile root segments of Ws and rat5 plants were infected with the nontumorigenic Agrobacterium strain GV3101 containing the binary vector pBISN1 (21). Two days after cocultivation, the roots were transferred to callus-inducing medium (CIM) containing timentin. Three days after infection, half of the segments were stained with X-gluc to determine the efficiency of transient GUS expression. The other group of segments was allowed to form calli on CIM. After 4 weeks, these calli were stained with X-gluc to determine the efficiency of stable GUS expression. (B) T-DNA integration in rat5 and Ws plants. Suspension cells were derived from the calli generated from Ws and rat5 root segments infected with the nontumorigenic Agrobacterium strain GV3101 containing the binary vector pBISN1. The suspension cell lines were grown for 3 weeks (without selection for transformation) in the presence of timentin or cefotaxime to kill Agrobacterium. Genomic DNA was isolated from these cells, subjected to electrophoresis through a 0.6% agarose gel, blotted onto a nylon membrane, and hybridized with a gusA gene probe. After autoradiography, the membrane was stripped and rehybridized with a phenylalanine ammonia-lyase (PAL) gene probe to determine equal loading of DNA in each lane.
We generated suspension cell lines from these Ws and rat5 calli, and after an additional month assayed the amount of T-DNA (using as a hybridization probe the gusA-intron gene located within the T-DNA of pBISN1) integrated into high molecular weight plant DNA from Ws and rat5 calli, as described (5, 11). Fig. 3B shows that although we could easily detect T-DNA integrated into the genome of wild-type Ws plants, we were unable to detect T-DNA integrated into the rat5 genome. These data demonstrate that rat5 is deficient in T-DNA integration. To demonstrate equal loading of plant DNA in each of the lanes, we stripped the gusA probe from the blot and rehybridized the blot with an Arabidopsis phenylalanine ammonia-lyase gene probe.
Discussion
To identify plant genes involved in Agrobacterium-mediated transformation, we screened a T-DNA-tagged Arabidopsis library for mutants that are resistant to Agrobacterium transformation (rat mutants). There are several steps in which plant genes are likely involved in the Agrobacterium-mediated transformation process. First, plant-encoded factors could be involved in the initial step of bacterial attachment to the plant cell surface. Mutants and ecotypes that are deficient in bacterial attachment have been identified (5, 6), and genes involved in bacterial attachment currently are being characterized. The next step is the transfer of T-strands from the bacteria to plant cells across the plant cell wall and membrane. Once the T-DNA/T-complex enters the cytoplasm of the plant cell, plant factors are required to transport the T-complex to the nucleus. The isolation of one such putative factor recently has been reported. Ballas and Citovsky (4) showed that a plant karyopherin α (AtKAPα) can interact with VirD2 nuclear localization sequences (NLS) in a yeast two-hybrid interaction system and presumably is involved in nuclear translocation of the T-complex. Using a similar approach, a tomato type 2C protein phosphatase, DIG3, that can interact with the VirD2 NLS was identified (Y. Tao, P. Rao, and S.B.G., unpublished work). Unlike AtKAPα, DIG3 plays a negative role in nuclear import. After the T-DNA/T-complex enters the nucleus, it must integrate into the plant chromosome. Plant chromosomal DNA is packaged into nucleosomes consisting primarily of histone proteins. The incoming T-DNA may have to interact with this nucleosome structure during the integration process. However, T-DNA may preferentially integrate into transcribed regions of the genome (32, 33). These regions are believed to be temporarily free of histones. How exactly T-DNA integration takes place is unknown. Recent reports have implicated involvement of VirD2 protein in the T-DNA integration process (10, 11). Plant proteins are also likely to be involved in this process.
Evidence for the involvement of plant factors in the T-DNA integration process comes from our ability to identify ecotypes and mutants of Arabidopsis that are deficient in T-DNA integration and hence recalcitrant to transformation (5, 6). We previously partially characterized an Arabidopsis T-DNA tagged mutant, rat5, that is deficient in T-DNA integration and is resistant to Agrobacterium-mediated root transformation. We showed that the T-DNA cosegregated with the tumorigenesis deficiency phenotype in the F2 segregating population of a cross between the wild-type Ws and rat5 (6). In this study we used both genetic and DNA blot analyses to demonstrate that there are two copies of T-DNA integrated as a tandem repeat at a single locus in rat5 (Fig. 1C). We additionally determined that there are no major rearrangements in the rat5 plant DNA immediately surrounding the T-DNA insertion site. These data strongly suggest that in rat5 the T-DNA had inserted into a gene necessary for Agrobacterium-mediated transformation. The sequence of the T-DNA left border-plant junction indicated that the T-DNA had inserted into the 3′ UTR of a histone H2A gene (Fig. 1B). This insertion is upstream of the consensus polyadenylation signal. By screening a Ws cDNA library and sequencing 20 different histone H2A cDNA clones, and by performing a computer database search, we have shown that there are at least six different histone H2A genes (K.S.M., H. Yi, and S.B.G., unpublished work). These genes encode proteins that are greater than 90% identical at the amino acid sequence level. Thus, the histone H2A genes comprise a small multigene family in Arabidopsis. At this point we are unable to explain the precise mechanism by which mutation of one of these genes can cause the rat phenotype.
There are several reports showing that mutation of one or two genes of a multigene family is sufficient to result in a detectable alteration in a phenotype. For example, mutation of one of the eight active Arabidopsis actin genes can be deleterious to the plant (34). The rat5 mutant does not have any other obvious morphological or developmental alterations compared with the wild-type parent. However, because of the deficiency in root transformation by Agrobacterium, it is possible that only a particular variant of H2A (RAT5) is involved in the transformation process. Earlier we hypothesized that although the rat5 mutation appears to be a loss-of-function mutant, its dominant nature could result from a gene dosage effect (6). Our ability to complement the rat5 mutant with a RAT5 gene, and the lack of a mutant phenotype displayed by wild-type plants transformed with the mutant rat5 gene, suggest that although the mutant appears dominant when rat5 plants are crossed to wild-type Ws plants, the mutant plant is actually haplo-insufficient. Because the T-DNA inserted into the 3′ UTR of the RAT5 gene, the effect of the mutation cannot be on the structure and consequent altered function of the RAT5 protein. There are several reports showing that the 3′ UTR of histone H2A mRNA is important for stability of the mRNA (35–38) and localization of mRNA on polyribosomes (39). Changes in histone H2A gene dosage also can alter transcription of other genes in yeast (40). It is possible that changes in dosage of this particular histone H2A variant can alter the chromatin structure of the plant target site and affect T-DNA integration. In yeast, reduced levels of histone proteins can alter retrotransposon (Ty) target site preference and disrupt an asymmetric Ty insertion pattern (41).
There are several reports of an inserted T-DNA not causing a particular mutant phenotype (42–47). To determine whether the T-DNA insertion caused the rat5 phenotype, a complementation analysis was performed. By expressing the RAT5 gene in the rat5 mutant we were able to restore the transformation proficiency phenotype (Fig. 2C; Table 1). This complementation test was confirmed by showing that the restored tumorigenesis phenotype cosegregated with the complementing RAT5 gene. These data indicate that a histone H2A (RAT5) is involved in Agrobacterium-mediated transformation. However, the rat5 mutant was not deficient in Agrobacterium-mediated germ-line transformation (48). This result demonstrates a requirement for histone H2A (RAT5) in the root tissue but not in the germ-line tissues for Agrobacterium-mediated transformation. To determine whether histone H2A plays a role in Agrobacterium-mediated transformation, the RAT5 gene was overexpressed in wild-type (ecotype Ws) Arabidopsis plants. Transgenic plants overexpressing RAT5 were approximately 2-fold more susceptible to Agrobacterium root transformation than were wild-type plants (Fig. 2D; Table 1). We also showed that transient expression of RAT5 was sufficient to complement partially the mutation in rat5 and also to increase the efficiency of T-DNA transformation of wild-type Ws plants. The presence of the RAT5 gene in a binary vector not only increased the efficiency of transformation to a phenotype encoded by the binary vector T-DNA, but also increased the transformation efficiency to a stable phenotype (tumorigenesis) encoded by the T-DNA of the Ti plasmid. These data indicate that histone H2A (RAT5) plays a role in Agrobacterium-mediated transformation. In the future it may be possible to increase the susceptibility of a plant to Agrobacterium transformation by either the transient or stable expression of RAT5.
Earlier we suggested that the rat5 mutant is likely deficient in T-DNA integration (6). Here we show direct evidence, by looking at the amount of T-DNA integrated in both Ws and rat5 plants, that the rat5 mutant is deficient in T-DNA integration (Fig. 3). The exact mechanism of involvement of histone H2A (RAT5) in T-DNA integration warrants further investigation. It will be of interest to determine whether mutations in other members of the histone H2A gene family result in the rat phenotype, and whether overexpression of these other family members can compensate for loss of RAT5 function in the rat5 mutant plant.
Acknowledgments
This work was supported by grants from the National Science Foundation (IBN-9630779) and the Consortium for Plant Biotechnology Research.
Abbreviations
- T-DNA
transferred DNA
- UTR
untranslated region
- GUS
β-glucuronidase
- NOS
nopaline synthase
Footnotes
References
- 1.Sheng J, Citovsky V. Plant Cell. 1996;8:1699–1710. doi: 10.1105/tpc.8.10.1699. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Zupan J R, Zambryski P. Crit Rev Plant Sci. 1997;16:279–295. [Google Scholar]
- 3.Deng W, Chen L, Wood D W, Metcalfe T, Liang X, Gordon M P, Comai L, Nester E W. Proc Natl Acad Sci USA. 1998;95:7040–7045. doi: 10.1073/pnas.95.12.7040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Ballas N, Citovsky V. Proc Natl Acad Sci USA. 1997;94:10723–10728. doi: 10.1073/pnas.94.20.10723. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Nam J, Matthysse A G, Gelvin S B. Plant Cell. 1997;8:873–886. [Google Scholar]
- 6.Nam J, Mysore K S, Zheng C, Knue M K, Matthysse A G, Gelvin S B. Mol Gen Genet. 1999;261:429–438. doi: 10.1007/s004380050985. [DOI] [PubMed] [Google Scholar]
- 7.Herrera-Estrella A, Chen Z-M, Van Montagu M, Wang K. EMBO J. 1988;7:4055–4062. doi: 10.1002/j.1460-2075.1988.tb03299.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Ward E R, Barnes W M. Science. 1988;242:927–930. [Google Scholar]
- 9.Howard E A, Winsor B A, De Vos G, Zambryski P. Proc Natl Acad Sci USA. 1989;86:4017–4021. doi: 10.1073/pnas.86.11.4017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Tinland B, Schoumacher F, Gloeckler V, Bravo Angel A M, Hohn B. EMBO J. 1995;14:3585–3595. doi: 10.1002/j.1460-2075.1995.tb07364.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Mysore K S, Bassuner B, Deng X-B, Darbinian N S, Motchoulski A, Ream W, Gelvin S B. Mol Plant-Microbe Interact. 1998;11:668–683. doi: 10.1094/MPMI.1998.11.7.668. [DOI] [PubMed] [Google Scholar]
- 12.Citovsky V, De Vos G, Zambryski P. Science. 1988;240:501–504. doi: 10.1126/science.240.4851.501. [DOI] [PubMed] [Google Scholar]
- 13.Christie P J, Ward J E, Winans S C, Nester E W. J Bacteriol. 1988;170:2659–2667. doi: 10.1128/jb.170.6.2659-2667.1988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Christie P J, Ward J E, Gordon M P, Nester E W. Proc Natl Acad Sci USA. 1989;86:9677–9681. doi: 10.1073/pnas.86.24.9677. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Rossi L, Hohn B, Tinland B. Proc Natl Acad Sci USA. 1996;93:126–130. doi: 10.1073/pnas.93.1.126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Matsumoto S, Ito Y, Hosoi T, Takahashi Y, Machida Y. Mol Gen Genet. 1990;224:309–316. doi: 10.1007/BF00262423. [DOI] [PubMed] [Google Scholar]
- 17.Gheysen G, Villarroel R, Van Montagu M. Genes Dev. 1991;5:287–297. doi: 10.1101/gad.5.2.287. [DOI] [PubMed] [Google Scholar]
- 18.Mayerhofer R, Koncz-Kalman Z, Nawrath C, Bakkeren G, Crameri A, Angelis K, Redei G R, Schell J, Hohn B, Koncz C. EMBO J. 1991;10:697–704. doi: 10.1002/j.1460-2075.1991.tb07999.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Ohba T, Yoshioka Y, Machida C, Machida Y. Plant J. 1995;7:157–164. doi: 10.1046/j.1365-313x.1995.07010157.x. [DOI] [PubMed] [Google Scholar]
- 20.Britt A B. Annu Rev Plant Physiol Plant Mol Biol. 1996;47:75–100. doi: 10.1146/annurev.arplant.47.1.75. [DOI] [PubMed] [Google Scholar]
- 21.Offringa R, de Groot M J A, Haagsman H J, Does M P, van den Elzen P J M, Hooykaas P J J. EMBO J. 1990;9:3077–3084. doi: 10.1002/j.1460-2075.1990.tb07504.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Paszkowski J, Baur M, Bogucki A, Potrykus I. EMBO J. 1988;7:4021–4026. doi: 10.1002/j.1460-2075.1988.tb03295.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Dellaporta S L, Wood J, Hicks J B. Plant Mol Biol Rep. 1983;1:19–22. [Google Scholar]
- 24.Sambrook M A, Fritsch E F, Maniatis T. Molecular Cloning: A Laboratory Manual. Plainview, New York: Cold Spring Harbor Lab. Press; 1982. [Google Scholar]
- 25.Becker D, Kemper E, Schell J, Masterson R. Plant Mol Biol. 1992;20:1195–1197. doi: 10.1007/BF00028908. [DOI] [PubMed] [Google Scholar]
- 26.Ditta G, Stanfield S, Corbin D, Helinski D R. Proc Natl Acad Sci USA. 1980;77:7347–7351. doi: 10.1073/pnas.77.12.7347. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Koncz C, Schell J. Mol Gen Genet. 1986;204:383–396. [Google Scholar]
- 28.Bent A F, Clough S J. In: Plant Molecular Biology Manual. Gelvin S B, Verma D P S, editors. Vol. 3. Dordrecht, the Netherlands: Kluwer; 1998. p. B7/1. 14. [Google Scholar]
- 29.Nacry P, Camilleri C, Courtail B, Caboche M, Bouchez D. Genetics. 1998;149:641–650. doi: 10.1093/genetics/149.2.641. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Ni M, Cui D, Einstein J, Narasimhulu S, Vergara C E, Gelvin S B. Plant J. 1995;7:661–676. [Google Scholar]
- 31.Narasimhulu S B, Deng X-B, Sarria R, Gelvin S B. Plant Cell. 1996;8:873–886. doi: 10.1105/tpc.8.5.873. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Koncz C, Martini N, Mayerhofer R, Koncz-Kalman Z, Korber H, Redei G P, Schell J. Proc Natl Acad Sci USA. 1989;86:8467–8471. doi: 10.1073/pnas.86.21.8467. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Herman P L, Jacobs A, Van Montagu M, Depicker A. Mol Gen Genet. 1990;224:248–256. doi: 10.1007/BF00271558. [DOI] [PubMed] [Google Scholar]
- 34.Gilliland L U, McKinney E C, Asmussen M A, Meagher R B. Genetics. 1998;149:717–725. doi: 10.1093/genetics/149.2.717. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Eckner R, Birnstiel M L. Nucleic Acids Res. 1992;20:1023–1030. doi: 10.1093/nar/20.5.1023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Brinstiel M L, Busslinger M, Strub K. Cell. 1985;41:349–359. doi: 10.1016/s0092-8674(85)80007-6. [DOI] [PubMed] [Google Scholar]
- 37.Hentschel C C, Birnstiel M L. Cell. 1981;25:301–313. doi: 10.1016/0092-8674(81)90048-9. [DOI] [PubMed] [Google Scholar]
- 38.Marzluff W F. Gene Exp. 1992;2:93–97. [PMC free article] [PubMed] [Google Scholar]
- 39.Piper P W, Aamand J L. J Mol Biol. 1989;208:697–700. doi: 10.1016/0022-2836(89)90159-9. [DOI] [PubMed] [Google Scholar]
- 40.Clark-Adams C D, Norris D, Osley M A, Fassler J S, Winston F. Genes Dev. 1988;2:150–159. doi: 10.1101/gad.2.2.150. [DOI] [PubMed] [Google Scholar]
- 41.Rinckel L A, Garfinkel D J. Genetics. 1996;142:761–776. doi: 10.1093/genetics/142.3.761. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Azpiroz-Leehan R, Feldmann K A. Trends Genet. 1997;13:152–156. doi: 10.1016/s0168-9525(97)01094-9. [DOI] [PubMed] [Google Scholar]
- 43.Errampalli D, Patton D, Castle L, Mickelson L, Hansen K, Schnall J, Feldmann K, Meinke D. Plant Cell. 1991;3:149–157. doi: 10.1105/tpc.3.2.149. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Feldmann K A. Plant J. 1991;1:71–82. [Google Scholar]
- 45.Franzmann L H, Yoon E S, Meinke D W. Plant J. 1995;7:341–350. [Google Scholar]
- 46.Koncz C, Nemeth K, Redei G P, Schell J. Plant Mol Biol. 1992;20:963–976. doi: 10.1007/BF00027166. [DOI] [PubMed] [Google Scholar]
- 47.Van Lijsebettens M, Vanderhaegen R, Van Montagu M. Theor Appl Genet. 1991;81:277–284. doi: 10.1007/BF00215734. [DOI] [PubMed] [Google Scholar]
- 48.Mysore, K. S., Ranjith-Kumar, C. T. & Gelvin, S. B. (2000) Plant J., in press. [DOI] [PubMed]