

Abstract
Functionally intact brain endothelial capillaries were isolated from porcine brain. p-Glycoprotein was localized at the lumenal membrane of intact capillaries by immunohistochemistry using a murine monoclonal antibody and a secondary FITC fluorescent labelled anti-mouse IgG. Western blot staining of p-glycoprotein in isolated endothelial cells confirmed the immunohistochemistry.
Excretion of the fluorescent labelled anthelmintic drug Ivermectin (BODIPY-Ivermectin) was studied in the isolated brain endothelial capillaries. Drug accumulation in the capillary lumen was visualized by fluorescence confocal laser scanning microscopy and was measured by image analysis. Secretion of BODIPY-Ivermectin into the capillary lumen exhibited characteristics of specific and energy-dependent transport. Steady state lumenal fluorescence intensity averaged 1.6 times cellular fluorescence and was reduced 3 – 4 times below cellular levels when metabolism was inhibited by NaCN.
BODIPY-Ivermectin secretion was inhibited in a concentration-dependent manner by unlabeled Ivermectin. In addition, lumenal but not cellular fluorescence intensity was significantly decreased when capillaries were incubated with PSC-833, Cyclosporin A or Verapamil, all inhibitors of p-glycoprotein. Conversely, unlabelled Ivermectin reduced the p-glycoprotein (Pgp)-mediated secretion of a fluorescent derivative of Verapamil, (BODIPY-Verapamil).
BODIPY-Ivermectin secretion was not affected in the presence of Leucotriene C4 (LTC4), a potent inhibitor of multidrug resistance related protein (mrp)-mediated transport processes. In addition, excretion of Fluorescein-Methotrexate, an mrp-substrate, was not inhibited by Ivermectin.
Uptake experiments with isolated porcine brain capillary cells showing increased cellular uptake of BODIPY-Ivermectin in the presence of unlabelled drug or PSC-833 supported the findings of a Pgp interaction in intact capillaries.
The data are consistent with BODIPY-Ivermectin and Ivermectin being transported across the lumenal membrane of brain capillaries. For the first time Pgp-interaction of Ivermectin at the blood brain barrier is demonstrated on a cellular level in an intact vascular tissue.
Keywords: Ivermectin, brain, capillary, blood – brain barrier, p-glycoprotein multidrug resistance related protein, confocal microscopy, active transport
Introduction
Ivermectin (22,23-dihydroavermectin B1a, Figure 1) is a semisynthetic analogue of avermectin B1, which belongs to a class of macrolide chemicals that act as agonists of γ-aminobutyric acid receptors in mammals and of glutamate-gated chloride channels in invertebrates (Bloom, 1996). Ivermectin is widely used in veterinary and human medicine for the treatment of parasitic infections. Although clinical safety has been proven, there are some contra-indications of the drug, e.g. meningitis, which may be associated with an impaired function of the blood – brain barrier. Earlier observations of side effects, such as central nervous system dysfunction (Vaughn et al., 1989; Paul et al., 1987) are now assumed to be related to an interaction of Ivermectin with p-glycoprotein (Lankas et al., 1997; Pouliot et al., 1997; Schinkel et al., 1994; Brown, 1998).
Figure 1.
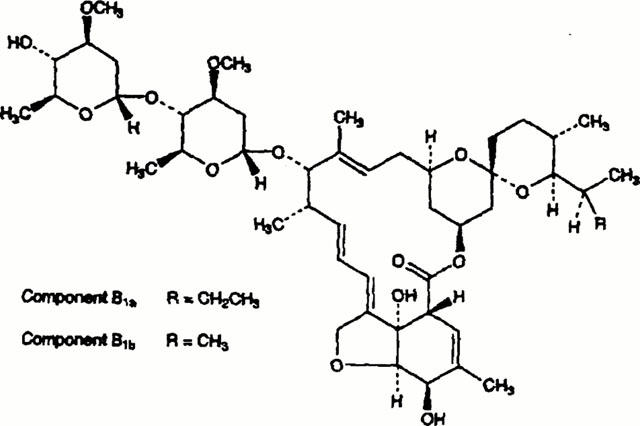
Molecular structure of Ivermectins (B1a: R=C2H5; B1b: R=CH3).
P-glycoprotein belongs to the family of membrane bound ATP-binding cassette (ABC) transport proteins and determines the pharmacokinetic behaviour of a variety, mainly cationic and lipophilic drugs (Gottesman & Pastan, 1993; Kusuhara et al., 1998; Müller & Jansen, 1997). It was originally discovered as an overexpressed protein in tumour cells with a multidrug resistant phenotype. Subsequently, it was also found at high levels in excretory or barrier tissues, such as in bile canalicular membranes of hepatocytes, the lumenal membrane of enterocytes in the gut, brush border membranes of renal proximal tubule or in brain capillary endothelium. In the latter, its exact localization is under debate: Some authors claimed that p-glycoprotein is not exactly localized within brain capillary endothelial cells, but in the foot processes of astrocytes surrounding the capillary wall (Pardridge et al., 1997; Golden & Pardridge, 1999; 2000).
In the present study, we used a model of freshly isolated, morphologically and functionally intact porcine brain capillaries and confocal laser scanning microscopy in order to investigate the excretory transport of a fluorescent Ivermectin derivative (BODIPY-Ivermectin). We further studied the ability of unlabelled Ivermectin to affect the excretion of fluorescent labelled drugs, which are handled by p-glycoprotein. The data show BODIPY-Ivermectin and Ivermectin being transported across the membrane of brain endothelial cells into the capillary lumen. Thus, for the first time Pgp-interaction of Ivermectin at the blood brain barrier is demonstrated on a cellular level in an intact vascular tissue.
Methods
BODIPY-Ivermectin, BODIPY-Verapamil and Fluoresceine-Methotrexate were purchased from Molecular Probes (Eugene, OR, U.S.A.). The fluorescent Cyclosporin-A derivative NBDL-CS was synthesized as described (Schramm et al., 1995). The p-glycoprotein blocker PSC-833 was obtained from Novartis AG, Basle, Switzerland. All other chemicals were purchased from commercial sources at the highest purity available.
Capillary isolation
Porcine brains were obtained from the local slaughterhouse and capillary vessels were isolated by mechanical disruption of cortical grey matter. Pieces of grey matter were gently homogenized by means of razor blades and a tight fitting teflon homogenizer in 3 volumes (v w−1) of buffer A (mM: NaCl 103, KCl 4.7, CaCl2 2.5, KH2PO4 1.2, MgSO4 1.2, HEPES 15). After addition of the same volume of dextran (final concentration 15%), the homogenate was centrifuged at 5800×g. The resulting pellet was resuspended in buffer B (corresponding to buffer A, supplemented with NaHCO3 (25 mM), glucose (10 mM), Na-pyruvate (1 mM), 0.5% (w v−1) bovine serum albumin, BSA) and filtered through a 200 μm nylon mesh. The filtrate was passed over a glass bead column and washed with 600 ml buffer B. The beads were carefully removed from the column and the adhering capillaries were removed by gentle agitation. The capillaries were centrifuged and the resulting pellet was resuspended in ice-cold, BSA-free Krebs-Henseleit-medium. The buffers were pregassed with carbogen (95% O2/5% CO2) and kept at 4°C.
Protein determination and analyses of enzymatic activities
The activities of the capillary marker enzymes alakaline phosphatase (AP; E.C.: 3.1.3.1.) and γ-glutamyl-transpeptidase (γ-GT, E.C.: 2.3.2.1.) were determined as quality markers of the purified capillaries. AP activity was measured by means of a commercial test kit (Sigma Diagnostics, St. Louis, MO, U.S.A.), the activity of γ-GT was determined following a earlier described method (Orlowsky & Meister, 1963). Protein concentrations were determined by the Bradford-method (Biorad, Munich, Germany).
Fluorescence microscopy
Individual capillaries were transferred to a aluminum foil-covered, teflon incubation chamber, which contained 0.5 – 1.0 ml of Krebs-Henseleit-medium with fluorescent compounds and added transport effectors. All concentrations are listed in the respective figure legends. The fluorescent compounds were dissolved in dimethylsulphoxide (DMSO) and added to the incubation medium. Earlier experiments had demonstrated that the concentrations of DMSO used (⩽1%) had no significant effect on uptake and excretion of the fluorescent labelled compounds. Analysis by HPTLC of BODIPY-Ivermectin and the other fluorescent compounds after extraction with ethanol out of the tissue gave no hint for degradation or transformation of the fluorescent compounds for at least the time of incubation (⩽1 h).
The chamber floor was a 2×2 cm glass cover slip to which the tissue could be easily viewed by means of an inverted confocal laser scanning microscope (Leica, Bensheim, Germany). The 488-nm laser line, and a 530 nm long-pass emission filter were employed. The tissue was viewed through a 63×water immersion objective. In order to circumvent photobleaching of the fluorescent compounds a low laser intensity was used (<30% of maximum intensity). Under the used conditions of photomultiplier gain settings the tissue autofluorescence was not detectable. The images were captured and analysed with the Scion Image software (Beta 3, Scion Corporation, Frederick, MD, U.S.A.). Normally, a field with several capillaries was selected and an epi-fluorescence image was aquired by averaging 4 – 8 frames. In earlier studies using fluoresceine as reference compound, it had been shown, that fluorescence intensity and fluoresceine concentration had an approximately linear relationship (Miller & Pritchard, 1991 and Miller, unpublished data). Here, the data are given as the average of measured pixel intensity rather than the estimated concentration of the fluorescent compounds. The intensities were measured as described for a similar system of renal proximal tubules (Miller, 1995; Schramm et al., 1995). From each capillary several adjacent cellular and lumenal areas were selected and the average pixel intensity for each area was calculated.
Cell isolation and cell culture
Porcine brain capillary endothelial cells were isolated and cultured as recently described (Gutmann et al., 1999b). Primary cultures were prepared according to Audus & Borchardt (1986) with the following modifications: Cortical grey matter from six freshly obtained porcine brains was minced and incubated in Medium 199, supplemented with L-glutamin (0.8 mM), Penicillin/Streptomycin (100 u ml−1; 100 μg ml−1), 100 μg ml−1 Gentamycin and HEPES, pH 7.4 (10 mM). Dispase (Roche, Mannheim, Germany) was added to a final concentration of 0.5% and the homogenate was incubated for 2 h at 37°C. Cerebral microvessels were obtained after centrifugation at 1000×g for 10 min and resupended in medium containing 15% dextran (Sigma, St. Louis, MO, U.S.A.) followed by centrifugation at 5800×g for 12 min. The microvessels were subsequently incubated in 20 ml medium containing 1 mg ml−1 collagenase-dispase (Boehringer) for 1.5 to 2 h. The resulting cell suspension was centrifuged for 10 min at 130×g and then, the cell pellet was resupended in 12 ml medium with 10% horse serum. This suspension was added to a discontinous Percoll (Pharmacia, Freiburg, Germany) gradient consisting of Percoll at 1.03 g ml−1 (20 ml) and 1.07 g ml−1 (15 ml). The percoll gradients were centrifuged at 1000×g for 10 min. Endothelial cells were enriched at the interface between the two Percoll solutions. Cells were collected and washed twice in medium containing 10% horse serum. The final cell suspension was kept in medium with 20% horse serum and 10% DMSO and stored in liquid nitrogen until use.
For uptake experiments, isolated endothelial cells were seeded with a density of 75,000 cells well−1 onto collagen (Roche, Mannheim, Germany) coated 96-well cell-culture plates. Cells were cultured under standard cell culture conditions using isolation medium without Gentamycin, but 10% horse serum. One day prior to transport experiments the medium was changed to 45% MEM, 45% F12-HAM, 100 μg ml−1 streptomycin, 100 μg ml−1 penicillin G, HEPES (10 mM), Glutamin (2 mM) and Hydrocortison (550 nM).
Western blot experiments
P-glycoprotein was detected by Western blot analysis using the monoclonal antibody (Mab) C219, (Alexis, Grünberg, Germany). The Pgp overexpressing cell line (MDR+)P388 was used as positive control for Western blots of Pgp. SDS-polyacrylamide gel electrophoresis was performed with a Mini-Protean II apparatus (Bio-Rad, Munich, Germany). Electrophoresis was carried out at a voltage of 80 V meanwhile the proteins where in the stacking gel and at a constant voltage of 120 V in the separating gel. Proteins were transferred electrophoretically to a nitro cellulose membrane using a Mini Trans-Blot cell (Bio-Rad) for 2.5 h at a constant amperage of 250 mA. The nitrocellulose membranes were incubated with antibody (1 : 20 dilution) in phosphate buffered saline containing 0.05% Tween 20, 1% BSA and 1% milk powder for 2 h at 37°C. Membranes were washed four times for 15 min and incubated 1 h at room temperature with horseradish peroxidase-conjugated rabbit anti-mouse IgG (1 : 1000; Roche, Mannheim, Germany) in posphate buffered saline containing 1% milk powder and 0.05% Tween 20. Then, the membranes were washed and p-glycoprotein was detected using enhanced chemiluminescence reagent (ECL; Amersham, Buckinghamshire, U.K.). Molecular weights were determined using commercially available standards.
Immunohistochemistry
For immunostaining, isolated brain capillaries were transferred onto slides and fixed for 20 min with 3% paraformaldehyde, 0.1% glutardialdehyde and 3.4% sucrose in PBS. After washing, the capillaries were permeabilized for 15 min with 1% (v v−1) Triton X-100 in PBS and again washed. Then, the capillaries were incubated for 1 h at 37°C in a humid chamber with the primary antibody (150 μl murine MAB C219, 1 : 20 dilution, Alexis, Grünberg, Germany). After washing the corresponding fluorochrome conjugated secondary antibody (150 μl FITC-conjugated rabbit anti-mouse IgG, 1 : 1000 dilution, Roche, Mannheim, Germany) was added for 1 h in a humid chamber in the dark.
Statistics
All pixels are given as mean±s.e.mean. The means were handled as statistically different, when the probability value (α) was less than 0.05 by use of an unpaired t-test.
Results
Capillary integrity
Brain endothelial capillaries were isolated from porcine brains by mechanical treatment of cortical grey matter. The capillary fragments were checked for morphological and enzymatic characteristics. Microscopic inspection yielded an average length of 150 – 350 μm of the capillary fragments. The endothelial marker enzymes alkaline phosphatase and γ-glutamyltranspeptidase showed an enrichment of activity compared to that of the brain homogenate of 32±1 and 19±4, respectively. Immunhistochemical assessment of p-glycoprotein in intact capillaries using a murine monoclonal antibody and a secondary FITC-conjugated rabbit anti-mouse antibody demonstrated a predominant localization of the protein at the lumenal membrane of the vessels (Figure 2A). Only a very faint staining could be seen at the ablumenal membrane of the capillaries. The presence of p-glycoprotein was confirmed by Western-blot analysis of 7-day-old isolated brain capillary endothelial cells kept in monolayer cultures (Figure 2B).
Figure 2.
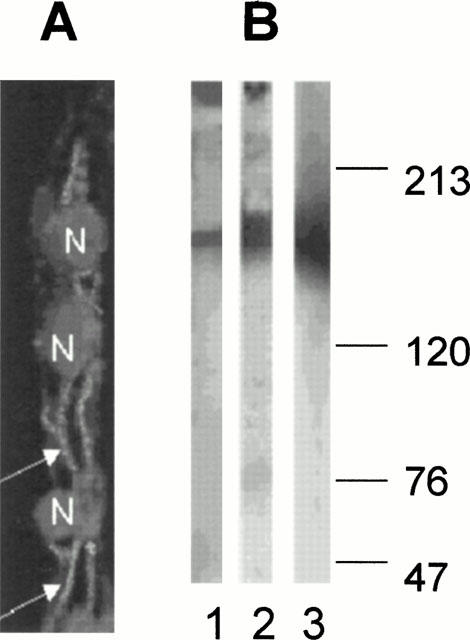
(A) Confocal image of porcine brain capillaries double stained with the monoclonal antibody versus Pgp and propidium iodide for nucleus (N) staining. Pgp staining can predominantly be observed at the lumenal membrane of the brain capillary endothelial cells (arrows). (B) Western blot detection of Pgp in isolated intact capillaries (1) and 7-day-old cultures of brain endothelial cells (2). Pgp-positive control (MDR+)-P388 cells (3). The numbers indicate molecular weight markers (in kD).
When the isolated capillaries were incubated with a marker for paracellular permeation (FITC-Dextran 4000) no fluorescence could be detected by confocal microscopy inside the capillary lumens within 60 – 80 min of incubation, indicating that the integrity of the tight junctions had been maintained during that time period. Thus, the isolated capillaries represent a significant barrier for the diffusion of large molecules across the endothelium, which is consistent with in vivo observations.
Transport studies
When porcine brain endothelial capillaries were incubated with micromolar concentrations of BODIPY-Ivermectin, the fluorescence started to accumulate in the cellular compartment within 3 – 5 min. After 5 – 10 min high fluorescence intensities could also be measured inside the capillary lumens. Steady state was reached within 25 – 30 min. Figure 3 shows a confocal image of a isolated capillary after 30 min incubation with 1 μM BODIPY-Ivermectin. Compared to the fluorescence intensity in the medium, which was almost undetectable under the chosen conditions, the intensities inside the cells were significantly higher. The fluorescence intensity levels inside the lumens at steady state exceeded those in the cells about 1.3 – 1.6 fold, suggesting uphill transport from bath to lumenal space. Such a pattern of fluorescence distribution under standard conditions had earlier been observed in a similar system of isolated, functionally intact renal proximal tubules and had been taken as an indication for active secretion from the cellular into the lumenal space (Schramm, 1995; Miller et al., 1997; Gutmann et al., 1999a; Fricker et al., 1999).
Figure 3.
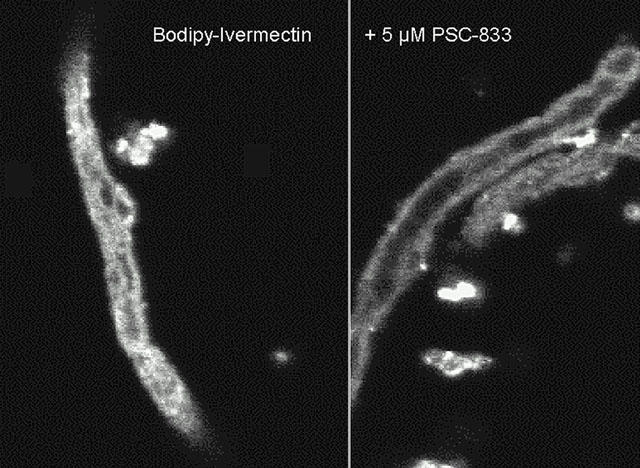
Confocal image of an isolated brain endothelial capillary incubated for 30 min with 1 μM BODIPY-Ivermectin (A). The fluorescent compound is excreted into the capillary lumen. When the capillary is pre-incubated with the p-glycoprotein blocker PSC-833 (B), BODIPY-Ivermectin is taken up into the cells, but not secreted any more.
When the cells were preincubated with 1 mM NaCN for 30 min, the steady state lumenal fluorescence was significantly reduced, whereas cellular fluorescence remained almost unchanged (Figure 4A). The ratio of lumenal to cellular fluorescence intensity decreased from 1.4±0.2 to 0.24±0.02 in control capillaries and in NaCN-exposed capillaries, respectively. The absolute cellular fluorescence intensity was comparable in control capillaries and in NaCN exposed tubules. This indicates, that only excretion but not uptake of BODIPY-Ivermectin is dependent on cellular metabolism.
Figure 4.
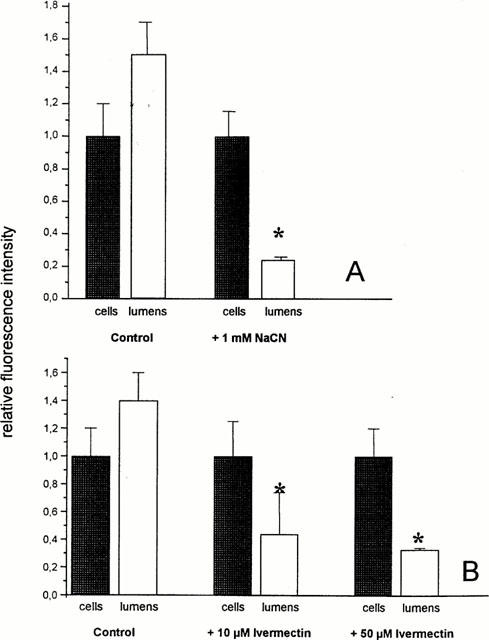
(A) Distribution of BODIPY-Ivermectin in control capillaries and in capillaries pre-incubated with 1 mM NaCN for 30 min (means±s.d.; n=12). (B) Distribution of BODIPY-Ivermectin in control capillaries and in capillaries incubated with increasing concentrations of unlabelled Ivermectin (means±s.d.; n=12). *Significantly different from control value (α<0.05).
In the presence of increasing concentrations of unlabelled Ivermectin a concentration dependent reduction of the lumenal fluorescence intensity could be seen, suggesting that transport of BODIPY-Ivermectin was specific and shared the same pathway as the unlabelled drug (Figure 4B).
When the capillaries were incubated with substrates and blockers of p-glycoprotein (PSC-833, Cyclosporin A, Verapamil) a significant decrease of lumenal, but not of cellular fluorescence could be seen (Figure 5A). This finding makes it very likely, that BODIPY-Ivermectin is secreted by p-glycoprotein across the lumenal membrane of brain capillaries. Conversely, when the transport of BODIPY-Verapamil, a substrate of p-glycoprotein was studied by confocal microscopy in the presence of unlabelled Ivermectin (Figure 5B), a reduction of the lumenal fluorescence was observed, whereas the cellular fluorescence intensity remained unchanged. To exclude that BODIPY-Ivermectin is excreted by another ABC-transport protein beside p-glycoprotein, we also studied the effect of Leucotriene C4, a substrate of proteins belonging to the mrp (multidrug resistance related protein) family, on BODIPY-Ivermectin excretion. LTC4 had no effect on cellular and lumenal fluorescence at steady state (Figure 6A). In addition, the excretion of Fluorescein-methotrexate, a fluorescent substrate of mrp, was not affected by unlabelled Ivermectin or PSC-833, but by LTC4 (Figure 6B).
Figure 5.
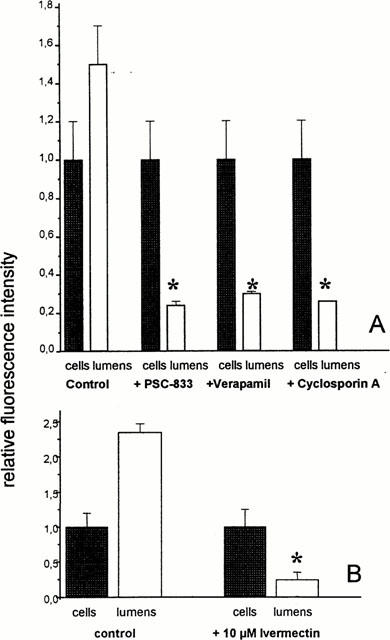
(A) Distribution of BODIPY-Ivermectin in control capillaries and capillaries incubated with substrates and blockers of p-glycoprotein (10 μM PSC-833, 10 μM Cyclosporin A, 50 μM Verapamil). Lumenal, but not cellular fluorescence is decreased (means±s.d.; n=10). (B) Distribution of the Pgp substrate BODIPY-Verapamil in the absence and in the presence of unlabelled Ivermectin. Lumenal fluorescence is significantly decreased (means±s.d; n=10). *Significantly different from control value (α<0.05).
Figure 6.
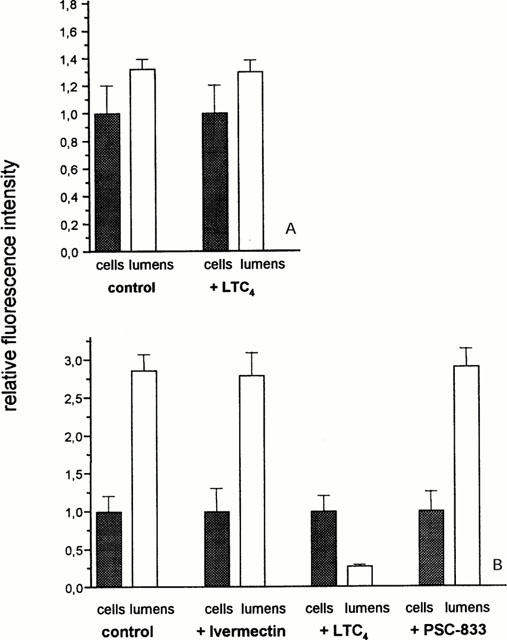
(A) Effect of the mrp-substrate LTC4 (0.3 μM) on cellular and lumenal distribution of BODIPY-Ivermectin (initial concentration in the incubation medium: 1 μM; means±s.d.; n=10). (B) Excretion of the mrp-substrate Fluorescein-Methotrexate (1 μM) in the absence and in the presence of BODIPY-Ivermectin (10 μM), LTC4 (0.3 μM) and PSC-833 (10 μM), respectively (means±s.d.; n=12). *Significantly different from control value (α<0.05).
In isolated brain capillary cells that were kept in monolayer cultures, the uptake of BODIPY-Ivermectin was significantly increased, when the cells were co-incubated with unlabelled Ivermectin or the p-glycoprotein blocker PSC-833 (Figure 7), supporting the finding in the capillaries of a p-glycoprotein interaction of BODIPY-Ivermectin.
Figure 7.
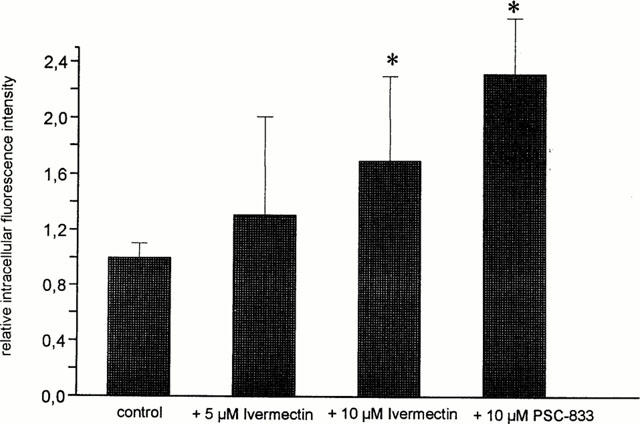
Uptake of BODIPY-Ivermectin into 7-day-old monolayer cultures of porcine brain capillary endothelial cells in the absence and in the presence of Ivermectin and the p-glycoprotein blocker PSC-833, respectively. Cells were incubated for 30 min with 10 μM BODIPY-Ivermectin (means±s.d.; n=6). *Significantly different from control value (α<0.05).
Discussion
The results of this study show, that a fluorescent labelled derivative of Ivermectin, BODIPY-Ivermectin, is transported from the incubation medium to the lumenal space of isolated functionally intact porcine brain capillaries by a process, that is concentrative, energy-dependent and specific. Since uptake into the endothelial cells was not affected by NaCN or one of the other compounds added, the pattern of transport is in accordance with the hypothesis of diffusional uptake of BI at the brain directed membrane of brain capillaries followed by a carrier-mediated and energy-dependent efflux across the lumenal membrane. Earlier observations in mdr1a knock-out mice make it very likely, that the underlying mechanism of that excretion is transport of BODIPY-Ivermectin and unlabelled drug, respectively, by p-glycoprotein: Treatment of mdr1a knock-out mice with Ivermectin exhibited a drastically increased neurotoxicity of the drug in these animals and significantly elevated drug concentrations in the brain of the ABC-transporter deficient animals compared to the concentration in brains of control animals (Schinkel et al., 1994; 1995). In a subpopulation of CF-1 mice showing mdr1-deficiency, concentration in the brain of Ivermectin or cyclosporin A (another p-glycoprotein substrate) was also higher in mdr (−/−) animals compared with mdr (+/+) mice after either i.v. or oral drug administration (Kwei et al., 1999). These mice, lacking placental p-glycoprotein, also showed an increased susceptibility of teratogenesis induced by an Ivermectin derivative (Lankas et al., 1998). In the same strain, the no-effect level after a single oral dose was about 100 μg kg−1, based on neurotoxicity, whereas other experimental animals, which were phylogenetically closer related to humans tolerated large doses of Ivermectin in the mg kg−1 range also after multiple dosing over 2 weeks (Brown, 1998). In humans, Ivermectin use is precluded in pregnant women, mothers nursing infants under 3 months old, children under 12 years old and people with CNS diseases like meningitis because of potential CNS side effects of the drug.
Our experiments demonstrate that fluorescent labelled Ivermectin is extruded out of intact brain capillaries by a concentrative effect and that this export process can be suppressed by p-glycoprotein substrates, whereas drug uptake remained unchanged. The excretion of BODIPY-Ivermectin seems to be p-glycoprotein specific. Transport was not inhibited by LTC4, a very potent inhibitor of proteins belonging to the p-glycoprotein related mrp-family, nor was secretion of Fl-methotrexate, which is mediated by mrp, inhibited by unlabelled Ivermectin. These data are consistent with our earlier observation of an interaction of BODIPY-Ivermectin and Ivermectin exclusively with p-glycoprotein in isolated functionally intact kidney tubules (Fricker et al., 1999). They also imply, that attention has to be paid to those drugs that are p-glycoprotein substrates and might exert CNS side effects during coadministration of Ivermectin. Examples of such drugs are HIV-protease inhibitors (Drewe et al., 1999; Gutmann et al., 1999a), PSC-833 or Cyclosporin A. For the latter, the effect of increased cerebral Ivermectin concentrations accompanied by neurotoxic side effects has been demonstrated in vivo in mice with an otherwise intact blood brain barrier (Marques-Santos et al., 1999).
The present findings further indicate, that p-glycoprotein is mainly located at the lumenal side of the capillary wall, as indicated by a strong and continous immunostaining. This is different from the hypothesis of an astrocyte related localization of Pgp in human tissue as described by others (Golden & Pardridge, 1999; 2000). Species differences, diverse antibodies may give an explanation for these different findings. The faint and irregular immunostainining at the outer surface of the capillaries does not completely exclude an abluminal localization of Pgp. However, the experimental setup of incubating isolated capillaries excludes the functional assessment of excretion into the incubation medium due to high dilution.
In addition, the study demonstrates, that isolated intact brain capillaries preserve functional activity of p-glycoprotein. Since the functional expression of transport proteins in cultured endothelial cells and cells lines may depend on the chosen culture conditions (El Hafny et al., 1997; Regina et al., 1998) the isolated capillaries represent an excellent in vitro tool to investigate the protective function of that ABC-transporter at the blood – brain barrier under conditions coming close to the in vivo situation.
Acknowledgments
Part of this work was supported by the Deutsche Forschungsgemeinschaft (grant GF/RU/6-2). P388 cells were kindly provided by Dr H. Gutmann, University Hospital Basle, CH.
Abbreviations
- AP
alkaline phosphatase
- BI
BODIPY-Ivermectin
- BSA
bovine serum albumin
- γ-GT
γ-glutamyl transpeptidase
- mrp
multidrug resistance related protein
- LTC4
Leucotriene C4
- Pgp
p-glycoprotein
References
- AUDUS K.L., BORCHARDT R.T. Characterization of an in vitro blood-brain barrier model system for studying drug transport and metabolism. Pharm. Res. 1986;3:81–87. doi: 10.1023/A:1016337202335. [DOI] [PubMed] [Google Scholar]
- BLOOM F.E. Goodman & Gilmann, The Pharmacological Basis of Therapeutics, 9th ed. McGraw-Hill: New York; 1996. Neurotransmission and the central nervous system; pp. 267–293. [Google Scholar]
- BROWN K.R. Changes in the use profile of Mectizan: 1987–1997. Annals Trop. Med. Parasitol. 1998;92 Suppl 1:S61–S64. doi: 10.1080/00034989859564. [DOI] [PubMed] [Google Scholar]
- DREWE J., GUTMANN H., FRICKER G., TÖRÖK M., BEGLINGER C., HUWYLER J. HIV protease inhibitor ritonavir: A more potent inhibitor of p-glycopotein than the cyclosporine analogue SDZ PSC-833. Biochem. Pharmacol. 1999;57:1147–1152. doi: 10.1016/s0006-2952(99)00026-x. [DOI] [PubMed] [Google Scholar]
- EL HAFNY B., CHAPPEY O., PICIOTTI M., DEBRAY M., BOVAL B., ROUX F. Modulation of P-glycoprotein activity by glial factors and retinoic acid in an immortalized rat brain microvessel endothelial cell line. Neurosci. Lett. 1997;236:107–111. doi: 10.1016/s0304-3940(97)00679-4. [DOI] [PubMed] [Google Scholar]
- FRICKER G., GUTMANN H., DROULLE A., DREWE J., MILLER D.S. Epithelial transport of anthelmintic Ivermectin in a novel model of isolated proximal kidney tubules. Pharm. Res. 1999;16:1570–1575. doi: 10.1023/a:1018956621376. [DOI] [PubMed] [Google Scholar]
- GOLDEN P.L., PARDRIDGE W.M. P-Glycoprotein on astrocyte foot processes of unfixed isolated human brain capillaries. Brain Res. 1999;819:143–146. doi: 10.1016/s0006-8993(98)01305-5. [DOI] [PubMed] [Google Scholar]
- GOLDEN P.L., PARDRIDGE W.M. Brain microvasular p-glycoprotein and a revised model of multidrug resistance in brain. Cell. Mol. Neurobiol. 2000;20:165–181. doi: 10.1023/a:1007093521681. [DOI] [PubMed] [Google Scholar]
- GOTTESMAN M.M., PASTAN I. Biochemistry of multidrug resistance mediated by the multidrug transporter. Annu. Rev. Biochem. 1993;62:385–427. doi: 10.1146/annurev.bi.62.070193.002125. [DOI] [PubMed] [Google Scholar]
- GUTMANN H., FRICKER G., DREWE J., TÖRÖK M., MILLER D.S. Interactions of HIV protease inhibitors with multiple ATP-dependent drug export proteins. Mol. Pharmacol. 1999a;56:383–389. doi: 10.1124/mol.56.2.383. [DOI] [PubMed] [Google Scholar]
- GUTMANN H., TÖRÖK M., FRICKER G., HUWYLER J., BEGLINGER C., DREWE J. Modulation of multidrug resistance protein expression in porcine brain capillary endothelial cells in vitro. Drug Metab. Dispos. 1999b;27:937–941. [PubMed] [Google Scholar]
- KUSUHARA H., SUZUKI H., SUGIYAMA Y. The role of p-glycoprotein and a canalicular multispecific organic anion transporter in the hepatobiliary excretion of drugs. J. Pharm. Sci. 1998;87:1025–1040. doi: 10.1021/js970100b. [DOI] [PubMed] [Google Scholar]
- KWEI G.Y., ALVARO R.F., CHEN Q., JENKINS H.J., HOP C.E., KEOHANE C.A., LY V.T., STRAUSS J.R., WANG R.W., WANG Z., PIPPERT T.R., UMBENHAUER D.R. Disposition of Ivermectin and cyclosporin A in CF-1 mice deficient in mdr1a P-glycoprotein. Drug Metab. Dispos. 1999;27:581–587. [PubMed] [Google Scholar]
- LANKAS G.R., CARTWRIGHT M.E., UMBENHAUER D. P-glycoprotein deficiency in a subpopulation of CF-1 mice enhances avermectin-induced neurotoxicity. Toxicol. Appl. Pharmacol. 1997;143:357–365. doi: 10.1006/taap.1996.8086. [DOI] [PubMed] [Google Scholar]
- LANKAS G.R., WISE L.D., CARTWRIGHT M.E., PIPPERT T., UMBENHAUER D.R. Placental P-glycoprotein deficiency enhances susceptibility to chemically induced birth defects in mice. Reprod. Toxicol. 1998;12:457–463. doi: 10.1016/s0890-6238(98)00027-6. [DOI] [PubMed] [Google Scholar]
- MARQUES-SANTOS L.F., BERNARDO R.R., DE PAULA E.F., RUMJANEK V.M. Cyclosporin A and Trifluoperazine, two resistance-modulating agents, increase Ivermectin neurotoxicity in mice. Pharmacol. Toxicol. 1999;84:125–129. doi: 10.1111/j.1600-0773.1999.tb00887.x. [DOI] [PubMed] [Google Scholar]
- MILLER D.S., PRITCHARD J.B. Indirect coupling of organic anion secretion to sodium in teleost (Paralichthys lethostigma) renal tabules. Am. J. Physiol. 1991;261:R1470–R1477. doi: 10.1152/ajpregu.1991.261.6.R1470. [DOI] [PubMed] [Google Scholar]
- MILLER D.S. Daunomycin secretion by killfish renal proximal tubules. Am. J. Physiol. 1995;269:R370–R379. doi: 10.1152/ajpregu.1995.269.2.R370. [DOI] [PubMed] [Google Scholar]
- MILLER D.S., FRICKER G., DREWE J. P-glycoprotein mediated transport of a fluorescent rapamycin derivative in renal proximal tubule. J. Pharmacol. Exp. Ther. 1997;82:440–444. [PubMed] [Google Scholar]
- MÜLLER M., JANSEN P.L. Molecular aspects of hepatobiliary transport. Am. J. Physiol. 1997;272:G1285–G1303. doi: 10.1152/ajpgi.1997.272.6.G1285. [DOI] [PubMed] [Google Scholar]
- ORLOWSKI M., MEISTER A. γ-Glutamyl-p-nitroanilide: A new convenient substrate for determination and study of L- and D-γ-glutamyltranspeptidase activities. Biochim. Biophys. Acta. 1963;73:679–681. doi: 10.1016/0006-3002(63)90348-2. [DOI] [PubMed] [Google Scholar]
- PARDRIDGE W.M., GOLDEN P.L., KANG Y.S., BICKEL U. Brain microvascular and astrocyte localization of p-glycoprotein. J. Neurochem. 1997;868:1278–1285. doi: 10.1046/j.1471-4159.1997.68031278.x. [DOI] [PubMed] [Google Scholar]
- PAUL A.J., TRANQUILLI W.J., SEWARD R.L., TODD K.S., JR, DIPIETRO J.A. Clinical observations in collies given Ivermectin orally. Am. J. Vet. Res. 1987;48:684–685. [PubMed] [Google Scholar]
- POULIOT J.F., L'HEUREUX F., LIU Z., PRICHARD R.K., GEORGES E. Reversal of P-glycoprotein associated multidrug resistance by Ivermectin. Biochem. Pharmacol. 1997;53:17–25. doi: 10.1016/s0006-2952(96)00656-9. [DOI] [PubMed] [Google Scholar]
- REGINA A., KOMAN A., PICIOTTI M., EL HAFNY B., CENTER MS., BERGMANN R., COURAUD PO., ROUX F. Mrp1 multidrug resistance-associated protein and P-glycoprotein expression in rat brain microvessel endothelial cells. J. Neurochem. 1998;71:705–715. doi: 10.1046/j.1471-4159.1998.71020705.x. [DOI] [PubMed] [Google Scholar]
- SCHINKEL A.H., SMIT J.J., VAN TELLINGEN O., BEIJNEN J.H., WAGENAAR E., VAN DEEMTER L., MOL C.A., VAN DER VALK M.A., ROBANUS MAANDAG E.C., TE RIELE H.P. Disruption of the mouse mdr1a p-glycoprotein gene leads to a deficiency in the blood brain barrier and to increased sensitivity to drugs. Cell. 1994;77:491–502. doi: 10.1016/0092-8674(94)90212-7. [DOI] [PubMed] [Google Scholar]
- SCHINKEL A.H., WAGENAAR E., VAN DEEMTER L., MOL C.A., BORST P. Absence of mdr1a p-glycoprotein in mice affects tissue distribution and pharmacokinetics of dexamethason, digoxin and cyclosporin A. J. Clin. Invest. 1995;96:1698–1705. doi: 10.1172/JCI118214. [DOI] [PMC free article] [PubMed] [Google Scholar]
- SCHRAMM U., FRICKER G., WENGER R., MILLER D.S. p-Glycoprotein mediated transport of a fluorescent cyclosporin analogue in teleost proximal tubules. Am. J. Physiol. 1995;268:F46–F52. doi: 10.1152/ajprenal.1995.268.1.F46. [DOI] [PubMed] [Google Scholar]
- VAUGHN D.M., SIMPSON S.T., BLAGBURN B.L., WHITMER W.L., HEDDENS-MYSINGER R., HENDRIX C.M. Determination of homovanillic acid, 5-hydroxyindoleacetic acid and pressure in the cerebrospinal fluid of collie dogs following administration of Ivermectin. Vet. Res. Commun. 1989;13:47–55. doi: 10.1007/BF00366852. [DOI] [PubMed] [Google Scholar]