
Figure 3.
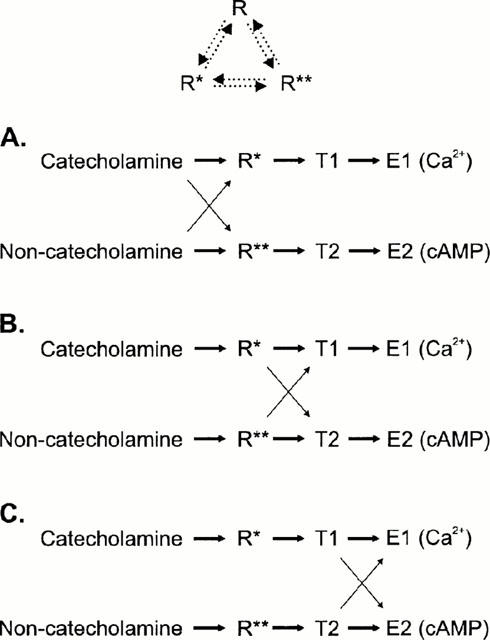
A schematic representation of the hypothesis with three somewhat different models, which are all possible based on the results. R is the inactive and R* and R** the two active receptor conformations, T1 and T2 are two signal transducers (e.g. G proteins) and E1 and E2 are the separate signal cascades which finally lead to a generation of a second messenger response (Ca2+ and cyclic AMP, respectively). The solid arrows indicate the direction of the response (the thicker the arrow, the stronger the coupling) and the dotted arrows in the top scheme the interconvertibility of the receptor conformations. In (A) a single receptor conformation only couples to a single transducer, which then couples to a single second messenger cascade. Catecholamines bind more strongly to R* whereas the other ligands bind more strongly to R** leading to a stronger activation of E1 (Ca2+) or E2 (cyclic AMP) response, respectively. In (B) the agonist trafficking of the receptor signals is accomplished at the level of receptor-transducer-coupling: the catecholamine-activated receptor (R*) is more effective in coupling to T1 (and thus to E1) whereas the non-catecholamine-activated receptor (R*) is more effective in its coupling to T2 (and thus to E2). In the third scheme, (C), a single receptor conformation only couples to a single transducer (as in A), which then couples to both second messenger cascades. The agonist trafficking of the receptor signals is brought about by the different ability of transducers to activate different cascades: the catecholamine-activated transducer T1 is more effective in coupling to E1 whereas the non-catecholamine-activated transducer T2 is more effective in its coupling to E2. The schemes (B) and (C) are essentially similar to the scheme presented in Berg et al. (1998), though somewhat more complex.