
Figure 4.
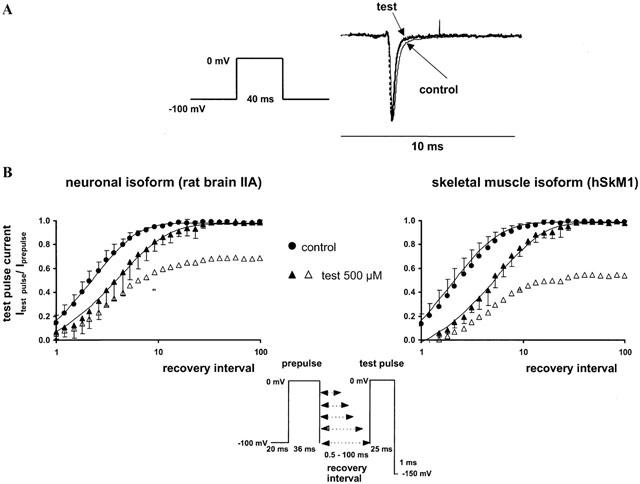
(A, upper panel) Acceleration of the sodium current decay phase by 500 μM 2,6-dimethylphenol in the skeletal muscle isoform. Representative current traces following a 40 ms depolarization to 0 mV in the control (solid line) and in the presence of 500 μM 2,6-dimethylphenol (test, dotted line). All currents were scaled to the same size for better comparison of the decay phase. (B, lower panel) Recovery from inactivation assessed by a two-pulse-protocol in the absence (control, circles) and presence of 500 μM 2,6-dimethylphenol (triangles). The abscissa represents the recovery time interval between prepulse and test pulse (up to 100 ms) on a logarithmic scale, and the ordinate represents the mean fractional current derived from at least three different experiments, having recovered from inactivation or inactivated channel block, respectively. Error bars are standard deviations. In the presence of 2,6-dimethylphenol, currents were normalized either to the prepulse in the presence of drug (filled symbols) or in the corresponding control experiment (empty symbols). Solid lines are bi-exponential fits (Equation 5) to the fractional current yielding the time constants of recovery, τrec. In the absence of the drug, recovery time constants τrec1 obtained for the neuronal and skeletal muscle isoform were 2.4±0.8 and 2.7±1.3 ms respectively. Additionally, we found a slow component τrec2 of 13.0±6.0 and 19.0±4.0 ms, which comprised 7 and 19% of the current amplitude. 2,6-dimethylphenol slightly increased the time constants of recovery from fast inactivation. τrec1 in the presence of 500 μM 2,6-dimethylphenol was 5.2±2.0 (P<0.01) and 6.0±1.6 ms (P=0.01); τrec2 was 17±8 and 20±10 ms. The amplitude of the slow component of recovery of about 20 ms increased up to 16±7 and 24±7% in the neuronal and skeletal muscle isoform in the presence of drug.