
TABLE 3.
Effect of gin mutation copy number on mosaicism
|
gin/+ × +/+ outcrosses
|
gin/+ × gin/+ homoallelic crosses
|
|||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Single-patch mosaics
|
Multiple-patch mosaics
|
|||||||||
| A | B | C | D | E | F | G | H | I | J | |
| Mutation | % single-patch mosaics | % multiple-patch mosaics | % single-patch mosaics | % multiple-patch mosaics | % gin/gin single-patch mosaics
|
% gin/gin multiple- patch mosaics
|
Prs(gin/+)
|
Prs(gin/gin)
|
Prm(gin/+)
|
Prm(gin/gin)
|
| C − A | D − B | 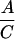 |
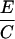 |
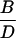 |
 |
|||||
| gin-1 | 0.2 | 0 (0/2564) | 0.7 | 0.3 | 0.5 | 0.3 | 0.29 | 0.71 | 0 | 1 |
| gin-2 | 0.2 | 0 (0/2622) | 1 | 1 | 0.8 | 1 | 0.20 | 0.80 | 0 | 1 |
| gin-3 | 0 (0/3171) | 0 (0/3171) | 0.4 | 0.3 | 0.4 | 0.3 | 0.00 | 1.00 | 0 | 1 |
| gin-4 | 0.2 | 0 (0/4224) | 0.3 | 0.4 | 0.1 | 0.4 | 0.67 | 0.33 | 0 | 1 |
| gin-5 | 0.7 | 0.1 (3/3039) | 1.3 | 0.5 | 0.6 | 0.4 | 0.54 | 0.46 | 0.2 | 0.8 |
| gin-6 | 0 (0/1584) | 0 (0/1584) | 0.7 | 0.7 | 0.7 | 0.7 | 0.00 | 1.00 | 0.0009 | 1 |
| gin-7 | 0.3 | 0 (0/2675) | 1 | 0 (0/786) | 0.7 | 0 | 0.30 | 0.70 | NA | NA |
| gin-8 | 0.1 | 0 (0/1617) | 0.9 | 0 (0/634) | 0.8 | 0 | 0.11 | 0.89 | NA | NA |
| gin-9 | 0.2 | 0 (0/2734) | 0.9 | 0.5 | 0.7 | 0.5 | 0.22 | 0.78 | 0 | 1 |
| gin-10 | 1.4 | 0.03 (2/5736) | 2 | 0.7 | 0.6 | 0.7 | 0.70 | 0.30 | 0.04 | 1 |
| gin-11 | 0.04 | 0 (0/2580) | 0.7 | 0.3 | 0.66 | 0.3 | 0.06 | 0.94 | 0 | 1 |
| gin-12 | 0.55 | 0 (0/2957) | 1.1 | 0.98 | 0.55 | 0.98 | 0.50 | 0.50 | 0 | 1 |
Heterozygote outcrosses were performed between gin/+ and +/+ parents, yielding 50% gin/+ and 50% +/+ progeny. Mosaics from these outcrosses (A and B) are presumed to have arisen among the gin/+ progeny. Homoallelic crosses between gin/+ parents yield 50% gin/+, 25% gin/gin, and 25% +/+ progeny. Mosaics from these crosses (C and D) are presumed to be due to both the gin/+ and the gin/gin progeny. The mosaics contributed by the gin/gin progeny (E and F) were calculated by subtracting the contribution of gin/+ embryos as determined from the outcrosses (A and B). The probability that single-patch mosaics from crosses between heterozygotes are due to gin heterozygosity, Prs(gin/+) (G), is the percentage of mosaic embryos arising among gin/+ progeny (A) divided by the percentage of mosaics among both gin/+ and gin/gin progeny (C). Derivation of probabilities of single-patch and multiple-patch mosaics Prm, with each genotype parallels this reasoning. The actual numbers of mosaic embryos and embryos scored are shown in parentheses when the percentage of mosaics approached 0. The data for columns A–D are from Table 2.