

Abstract
The relationship between sensory sensitivity and reading performance was examined to test the hypothesis that the orthographic and phonological skills engaged in visual word recognition are constrained by the ability to detect dynamic visual and auditory events. A test battery using sensory psychophysics, psychometric tests, and measures of component literacy skills was administered to 32 unselected 10-year-old primary school children. The results suggest that children's sensitivity to both dynamic auditory and visual stimuli are related to their literacy skills. Importantly, after controlling for intelligence and overall reading ability, visual motion sensitivity explained independent variance in orthographic skill but not phonological ability, and auditory FM sensitivity covaried with phonological skill but not orthographic skill. These results support the hypothesis that sensitivity at detecting dynamic stimuli influences normal children's reading skills. Vision and audition separately may affect the ability to extract orthographic and phonological information during reading.
Learning to read a language depends on acquiring an understanding of both its spoken properties (phonology) and its written form (orthography). In alphabetic languages such as English, printed characters (or graphemes) correspond to phonemes, the smallest meaningful units of sound that amalgamate to constitute spoken words. However, unlike other languages such as Spanish or German where the relationship between letters and sounds is relatively invariant, readers of English are presented with the problem that most letter combinations can be mapped to more than one phoneme. Phoneme identity in English depends much more on the preceding and subsequent letter context than in other alphabetic languages with more regular grapheme-phoneme mappings (1). Grapheme-phoneme correspondence rules are therefore useful in learning to read unknown regular words such as “market,” but they do not provide sufficient information for successfully decoding exception words like “quay.” Identification of exception words requires orthographic coding skill, the ability to use and identify familiar letter sequences with minimal aid from phonological information (2, 3). Phonological processing may occur during orthographic (de)coding, but the result of such processing is not sufficient to determine the identity of a lexical string (3). Which word features comprise orthographic information is inconsistently applied in the literature, and no standardized measure of orthographic coding skill yet exists. Orthographic information could encompass any of a number of sources in visual word recognition such as letter order or letter frequency in a given position within a word (4), the perceived pronunciation of a letter string (5), and the rules that constrain phonemic and graphemic information within words (6). Therefore the grain size of orthographic information can range from the single letter, through syllables and onset-rimes (7), to whole word forms (for review see ref. 8).
A large body of research has shown the importance of phonological skill in learning to read (for review see ref. 9). Competent phonological awareness, characterized by the ability to switch attention from a word's meaning to an analysis of its acoustic characteristics (10), has been demonstrated experimentally by using a number of paradigms that require word sound identification and/or manipulation (11–13). Proficiency on such tasks can reliably discriminate good from poor readers (14, 15) and can predict the future literacy skill of prereaders (16). Phonological representations in memory have been argued to result either from a mechanism specific to higher-level linguistic functions such as the categorical perception of speech segments (17, 18) or from more basic auditory processing skills (19, 20) such as those engaged when following the acoustic modulations that are present in speech (21). In either case it is clear that the accurate detection of the fine-grained distinctions characteristic of phonemes in spoken language is important for the development of proficient reading skills.
In addition to proficient phonological skills, orthographic sensitivity is highly important to the acquisition of reading skills (2, 22, 23). The strong correlation between measures of orthographic skill and reading ability remains after accounting for the large amount of statistical covariance with phonological ability and after removing variance attributable to individual differences in print exposure (2). Orthographic skill is therefore an acknowledged source of independent variance in literacy skills (2, 24). This observation has been largely confirmed by twin studies (refs. 3 and 25, cf. ref. 12). Olson and colleagues showed that significant variance in both phonological and orthographic skill can be attributed to heritable factors, and that each of these factors contributes independently to variability in word recognition.
Studies of word recognition deficits in persons with specific reading disabilities have been used as evidence for orthographic and phonological coding as separable and independent processes in word recognition (26, 27). Although most current developmental and connectionist models of single-word reading propose that orthographic and phonological processes rarely operate autonomously (28–33), both routes are important for the development of competent reading skills. Because the nature of the letter sound mappings in English is inconsistent, most successful methods for teaching reading have used training on both printed letter and word sounds (i.e., both orthographic and phonological coding) (16, 34–36).
In addition to their poor literacy skills, dyslexic readers have been found to be less sensitive to dynamic sensory stimuli, both auditory and visual (for review see refs. 37–40). Deficits for detecting such rapidly presented or rapidly changing stimuli are argued to play a direct role in dyslexic's problems with word decoding (19–21). Although this sensory hypothesis is still controversial (17, 18), substantial empirical evidence suggests that insensitivity to the temporal auditory changes characteristic of many phonemes is associated with their phonological problems (refs. 19–21 and 41, cf. ref. 42]. Similarly there is good evidence that difficulties in detecting visual changes such as coherent motion hinders the ability to extract letter position information during the early stages of visual print analysis (43, 44). For example, poor visual motion sensitivity is associated with reduced performance on a word/anagram decision task requiring the extraction of orthographic information about letter position (43, 44). Temporal visual processing also has been linked to nonword reading ability, a sensitive measure of phonological skill (21, 45–47).
Our hypothesis is that sensitivity to dynamic visual and auditory stimuli influences the development of literacy skills in normal readers. We have focused on single-word decoding measures because accessing the lexicon of visual/phonological word forms while reading is the primary constraint on proficient reading comprehension (for review see ref. 3). However, this sensory processing hypothesis has been derived mainly from experiments involving selected dyslexic and control groups rather than unselected children who are learning to read. These previous results therefore might apply only to selected adults and not to the abilities of normal children. In this study therefore we have tested whether visual and auditory transient sensitivity is a useful predictor of reading and spelling ability in unselected primary school children. In a class of 10-year-olds, just about to leave primary school, we have compared sensitivity to simple visual motion and auditory FM stimuli with reading and spelling, phonological and orthographic decoding proficiency. If sensory processing is important for the development of reading in general across the normal range then it should covary with reading skill. Furthermore, visual processing skills would be predicted to covary most strongly with measures of orthographic sensitivity, whereas auditory sensitivity would be most related to phonological task performance.
Methods
Subjects.
Our experimental protocol was administered in accordance with the guidelines of the Declaration of Helsinki and had local ethics committee approval. The subjects were 32 unselected children, around the age of 10 years, who comprised all of the students in a single, mainstream primary school classroom. None of the children had any history of learning disability at the time of testing. The entire battery of tests took approximately 1¼ hr to complete. The group was equally divided between males and females, and all of the participants were native English speakers.
Psychometric Assessments.
The children's cognitive, reading, and spelling abilities were estimated by using standardized measures of achievement from the British Abilities Scales (BAS) (48). Cognitive measures sampled both verbal (similarities) and nonverbal (matrices) performance as well as short-term verbal memory (digit span). These measures comprise three of the four subscales suggested for determining a child's short form intelligence quotient (48). BAS reading and spelling subscales were used to assess literacy skill. Table 1 shows the average performance of our sample on these measures.
Table 1.
Averaged group performance for the psychometric measures used in the study
| Measure (units) | Mean (SD) |
|---|---|
| Age (months) | 118.4 (4.5) |
| BAS average (t-score) | 51.9 (6.0) |
| BAS reading (t-score) | 55.8 (10.6) |
| BAS spelling (t-score) | 52.5 (7.5) |
| Exception word naming (% correct) | 65.0 (15.8) |
| Nonword naming (% correct) | 73.6 (20.2) |
| Spoonerisms (% correct) | 62.3 (20.9) |
| Pseudohomophone identification (% correct) | 82.6 (12.2) |
| 2-Hz FM detection (mod. index) | 4.4 (2.8) |
| 240-Hz FM detection (mod. index) | 4.1 (2.8) |
| Coherent motion detection (% coherence) | 18.6 (10.0) |
In addition to standardized achievement tests, other tasks were used to assess children's orthographic and phonological decoding skills, as follows.
The nonword and exception word naming tasks came from the Castles and Coltheart battery (26). The tasks comprised 30 each of irregularly spelled words (e.g., “colonel”) and nonwords (e.g., “tegwop”). The latter are called nonwords because, although they lack meaning, they can be pronounced by applying grapheme to phoneme correspondence rules. The children attempted to name each word, which was presented visually to them in a list, and were instructed to proceed as quickly as possible without making errors. Percent correct for each list was recorded.
The Spoonerisms section of the Phonological Assessment Battery (13) assesses children's ability to manipulate phonemes in words presented to them orally and therefore does not involve any visual processing of print. It has three sections that contain increasingly difficult phoneme elisions: simple phoneme deletion and substitution (e.g., replace the first sound in “dog” with “l” to make “log”); complex phoneme deletion and substitution (e.g., replace the first sound in “lip” with the first sound in “pig” to make “pip”); and Spoonerisms (e.g., swap the first sounds of “little, pup” to make “pittle, lup”). Each child was given a maximum of 3 min to complete each section of 10 items. Percent of items correct was recorded.
The ability to discriminate real words from pseudohomophones (e.g., “rain” vs. “rane”) was used as a measure of orthographic sensitivity (3, 49). This is considered a test of orthographic skill because phonological analysis alone cannot discriminate between the pseudohomophone and the real word target. Eighty-eight word/pseudohomophone pairs were presented on a computer screen in 18-point Geneva font. The children were instructed to view both words and decide which one was spelled correctly, guessing when necessary. The stimulus duration of each word pair was not restricted but the children were told that each response was being timed, and therefore it was important to proceed as quickly as possible without sacrificing accuracy. Number correct and response time for the 80 targets, after eight practice trials, was recorded by computer.
Psychophysical Paradigms.
Detection thresholds were measured for 2 Hz and 240 Hz FM of a 1-kHz tonal carrier, by using a standard two-interval, two-alternative forced-choice paradigm. In both cases, the target interval contained sinusoidal FM; defined in Eq. 1 and the other interval contained a pure tone as defined by Eq. 2.
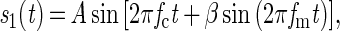 |
1 |
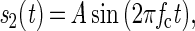 |
2 |
where Sn(t) = the signal as a function of time, A = the intensity of the signal, fc = frequency of the carrier (1 kHz in this study), fm= modulation rate, t = time, β = the modulation index (the modulation depth expressed as the frequency deviation in Hz divided by the modulation rate).
All sounds were generated with Tucker-Davis Technologies (Gainesville, FL) System II equipment. Signals were generated digitally and passed to 16-bit digital to analogue converters with a sampling rate of 40 kHz. Stimuli were low-pass filtered at 15 kHz, attenuated, and presented through calibrated Sennheiser HD40 headphones in a quiet room. Each of the two stimulus intervals was 1,000 ms in duration and separated by a 500-ms silent inter-stimulus interval. Sounds in each interval were gated on and off with 20-ms Gaussian rise and fall times. Absolute thresholds for a 1-kHz tone had been measured for all subjects to ensure they had no hearing loss at this frequency. Sounds were presented at a comfortable 60-dB hearing level. Subjects were required to report verbally which interval, first or second, contained the modulation. They were given feedback about their performance on the computer screen. Before collection of data, subjects were given a short period of practice, about five supra-threshold trials. This procedure allowed the experimenter to estimate the subject's threshold, to set the appropriate range of stimulus depths for the psychometric function, and to ensure the child understood the experimental protocol. Ten trials were performed at each of six stimulus depths, chosen to span the subject's threshold. The percentage of correct responses was plotted against the stimulus depths, to produce a psychometric function, which was fitted with a Weibull function (50). Detection threshold was taken to be the stimulus depth at which subjects responded correctly on 75% of the trials.
The coherent motion stimuli were similar to those used in other studies of children and infants (51). They comprised two patches of 300 high luminance (80.6 cd/m2), white dots (1 pixel), presented on the dark background (0.98 cd/m2) of a LCD laptop computer display. Michelson contrast [(Lmax − Lmin)/(Lmax + Lmin)] between the luminance (L) of the dots and the background was held at a constant 97.6%. Binocular viewing was conducted in a darkened room, illuminated only by the computer display. All luminance measures were obtained with an OptiCal digital photometer (Cambridge Research Systems, Cambridge, U.K.). At the constant viewing distance of 57 cm, each patch subtended 10 × 14° visual angle, separated horizontally by 5°.
The percentage of coherently moving dots (angular velocity = 15.1°/sec) within a given software animation frame (50 ms) was determined algorithmically and varied adaptively to the subject's detection threshold. This threshold was defined as the proportion of dots required for the subject to detect reversing motion in either the left or right panel. The coherent dots changed direction every 1,000 ms throughout the 2,500-ms stimulus interval. The noncoherent dots moved randomly between frames in a Brownian manner. To eliminate the possibility of detecting the direction of coherent motion by following the trajectory of a single dot, each dot had a fixed lifetime of five animation frames (250 ms) after which it would disappear before being regenerated at a random place within the stimulus patch. Percentage of coherent motion is corrected for a finite dot lifetime of five frames. When all dots are moving coherently during an animation frame, this is described as 80% coherence.
The children were asked to visually inspect each stimulus patch carefully and report which patch contained coherent motion by pointing with their finger. The experimenter recorded these responses by pressing an appropriate key on the computer. Coherent motion was varied to the subject's motion detection threshold by a 3-dB-up-1-dB-down, two alternative forced-choice staircase procedure (52) with a constant starting value of 70% coherence. This value is well above the detection threshold for even those with the poorest motion sensitivity. This staircase procedure decreases the motion coherence by a factor of 1.122 (1 dB) for every correct response whereas incorrect responses result in the coherence value being raised by a factor of 1.412 (3 dB), where dB is defined by Eq. 3.
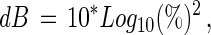 |
3 |
where % = percent coherent dot motion.
Threshold estimates were determined by taking the geometric average of the last 8 of 10 reversal points within a given series of trials. Each series was repeated twice with the arithmetic mean of these two estimates defined as the subject's coherent motion detection threshold.
There is an extensive literature showing that both good and poor readers in the age range of our study can reliably complete protocols of visual and auditory psychophysics [for review see refs. 40 and 53]. Nevertheless we also monitored each child's performance to ensure that they understood the tests and could complete them reliably by giving them practice trials at stimulus values that were well above their detection threshold. For the coherent motion task, each staircase was run twice. The correlation between these threshold estimates was 0.71, indicating an acceptable test-retest reliability. For both FM tasks, the equal interval design included modulation rates that were set well above the child's threshold. Hence, each child was exposed to at least one modulation rate at which she or he could respond at 100% correct over 10 stimulus presentations.
Results
Can Dynamic Sensory Sensitivity Predict Reading Skill?
Kolmogorov-Smirnov Z tests showed that the distribution of each of the measured variables did not significantly differ from normal. Pearson product moment correlations therefore were justified to determine the nature of the relationship between the various study measures (Table 2).
Table 2.
Pearson product-moment correlations between study measures
| MAT | SIM | DIG | READ | SPEL | NON | IRR | SPO | ORT | MOT | FM2 | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| SIM | 0.54** | ||||||||||
| DIG | 0.23 | 0.24 | |||||||||
| READ | 0.43* | 0.44* | 0.25 | ||||||||
| SPEL | 0.41* | 0.39* | 0.35 | 0.79*** | |||||||
| NON | 0.48** | 0.37* | 0.45* | 0.82*** | 0.79*** | ||||||
| IRR | 0.32 | 0.39* | 0.38* | 0.83*** | 0.67*** | 0.71*** | |||||
| SPO | 0.48** | 0.38* | 0.44* | 0.69*** | 0.71*** | 0.83*** | 0.63*** | ||||
| ORT | 0.32 | 0.38* | 0.34 | 0.88*** | 0.80*** | 0.76*** | 0.86*** | 0.61** | |||
| MOT | 0.51** | 0.40* | 0.36* | 0.31 | 0.49** | 0.37* | 0.40* | 0.48** | 0.48** | ||
| FM2 | 0.43* | 0.45* | 0.45* | 0.67*** | 0.70*** | 0.80*** | 0.69*** | 0.70*** | 0.66*** | 0.39* | |
| FM240 | 0.30 | 0.26 | 0.41* | 0.49** | 0.53** | 0.59** | 0.51** | 0.55** | 0.40* | 0.30 | 0.43* |
MAT, matrices; SIM, similarities; DIG, digit span; READ, reading; SPEL, spelling; NON, nonword reading; IRR, exception word reading; SPO, Spoonerisms decoding; ORT, word-pseudohomophone discrimination; MOT, visual coherent motion sensitivity; FM2, 2-Hz auditory FM sensitivity; FM240, 240-Hz auditory FM sensitivity. *, P ≤ 0.05; **, P ≤ 0.01, ***, P ≤ 0.001.
Children's performance on the orthographic word-pseudohomophone discrimination task was the strongest statistical predictor of their literacy skills, as measured by single-word reading and spelling. It alone could explain over 78% of the variance in reading skill and over 63% of the variance in spelling ability (see Table 2). The phonological nonword naming test accounted for 5% of the remaining variance in reading and an additional 8% of the variance in spelling after removing the effects of word-pseudohomophone discrimination. Thus, when combined, these two variables accounted for over 83% of the variability in children's reading [F(2,29) = 74.6, P < 0.001] and over 71% of the variance in children's spelling [F(2,29) = 36.1, P < 0.001]. None of the other psychometric variables were found to be significant predictors of reading or spelling after the variance attributable to these two factors was removed. Therefore, consistent with previous research, we have verified the strong predictive power of orthographic and phonological tests to predict literacy skill. We also replicated the finding that, although there is substantial statistical covariation between the separate phonological and orthographic measures (minimum r = 0.61, Table 2), they are not entirely redundant statistically (2, 3, 54). That is, both orthographic and phonological tests account for unique variance in normal children's literacy skills.
The purpose of our study was to examine whether sensory sensitivity, either auditory, visual, or both, can predict normal children's literacy skills. If they do, can these sensory skills be related to particular aspects of word reading, namely phonological and orthographic decoding of print? It is apparent that that our sensory tasks were significant predictors of the literacy skills of the children we studied (Table 2). Threshold for 2-Hz FM was the strongest single predictor of both reading (45%) and spelling (49%) skill. This finding rivals the predictive power of the majority of the literacy skill measures we administered. When the three measures of auditory and visual sensory processing were entered together as predictors of reading and spelling ability in hierarchical regression analyses, they could account for over 51% of reading skill [F(3,28) = 9.4, P < 0.001] and over 59% of spelling skill [F(3,28) = 13.5, P < 0.001]. Sensitivity to 240-Hz FM predicted a small, but significant, proportion of independent variance in reading (5%) but not spelling. Coherent motion detection accounted for additional independent variance in spelling (8%) but not reading, and motion detection alone was a significant predictor (24%) of spelling ability, but not reading skill (Table 2). These results confirm previous findings demonstrating a moderate to strong relationship between dynamic sensory processing and reading skills (21). These findings also suggest that auditory and visual processing may separately influence various literacy skills such as single-word reading and spelling. Most importantly they provide direct evidence for a relationship between sensory processing skills and the literacy skills of normal children rather than in selected dyslexic readers compared with controls.
Does Sensory Processing Explain Independent Variance in Children's Literacy Skills?
Our hypothesis is that the mechanism by which sensory processing influences reading skill occurs in the early stages of text processing; hence it should affect the ability to extract orthographic and phonological information while reading. Although this bottom-up hypothesis is difficult to test directly, we can examine the relationship between sensory and linguistic processing by removing top-down effects, such as overall reading skill and intelligence. Such effects may obscure important covariance between our sensory and lexical processing measures. Reading ability was removed because the reciprocal nature of the causal relationship between reading and phonological awareness (55, 56) may obscure the relationship between our sensory and phonological processing measures. Each of our component language tasks, with the exception of the Spoonerisms measure, also involves the processing of print for the successful completion of the task. Reading ability is strongly related to print exposure (2, 25); hence removing the overall effects of reading skill also may account for the variance it shares with print exposure.
Similarly, individual differences in intelligence, including short-term memory, may account for significant inter-subject variability in psychophysical task performance (53) and tests of phonological ability (57). Consistent with this evidence, our data showed that children's performance on a single BAS test of cognitive ability could account for up to 23% of the variation in the psychophysical tasks and also in the literacy measures (Table 2). Variance attributable to overall reading skill and intelligence was removed in regression analyses to ensure that any predictive relationships among variables found in subsequent analyses were not caused by intersubject differences in these measures.
Accounting for overall differences in intelligence and reading skill serves a second purpose as it removes shared variance between the orthographic and phonological performance measures. Coltheart and Leahy (54) have reported a high correlation (r = 0.69) between nonword reading and exception word reading in a large sample of children. We found a similarly strong correlation (r = 0.71) between these measures (Table 2). One explanation for this result is that a substantial proportion of the variance common to each measure is accounted for by a third factor such as reading ability, intelligence, or print exposure. This prediction was confirmed when the effects of overall reading skill and intelligence were removed; the highest intercorrelation between any orthographic and phonological measure dropped to 0.10, whereas the correlation between the two orthographic measures (r = 0.48) and between the two phonological measures (r = 0.58) remained strong.
Table 3 shows the partial correlation between our measures when the variance attributable to reading ability and intelligence was removed. Before this step the two measures of orthographic processing (pseudohomophone detection and irregular word reading) and phonological processing (nonword reading and spoonerisms), which are highly correlated with one another (Table 2), were scaled equally and combined by linear principal components analysis (PCA). PCA provides a method for optimally scaling two related variables. The first principal component for each pair of tests was used as a unitary measure of orthographic and phonological decoding skill, respectively. Multiple regression analyses were used to test the relationship between these scaled measures and sensitivity on our sensory tasks. When 2-Hz FM, 240-Hz FM, the component orthographic measure and coherent motion detection, were entered as predictor variables of phonological processing, 2-Hz FM could account for 24% of the variance. No other variable accounted for significant additional variance. In contrast, when the auditory measures, the component phonological measure, and motion detection were entered as predictor variables of orthographic skill, only motion detection accounted for significant variance (20%). No other variable made a significant contribution beyond that accounted for by motion detection performance alone. Together these analyses suggest that normal children's orthographic and phonological word decoding skills covary with their temporal sensitivity to visual and auditory stimuli, respectively. Orthographic skill covaried with coherent motion detection, which is a sensitive measure of visual magnocellular function (58–60); whereas phonological skill was predicted by 2-Hz FM (but not 240-Hz FM) sensitivity.
Table 3.
Partial correlations between measures after removing variance attributable to individual differences in intelligence and reading skill
| ORT | PHO | MOT | FM2 | |
|---|---|---|---|---|
| PHO | 0.08 | |||
| MOT | 0.45** | 0.21 | ||
| FM2 | 0.30 | 0.49** | 0.09 | |
| FM240 | 0.08 | 0.34 | 0.09 | 0.07 |
ORT, principal component (PC) of irregular word reading and word-pseudohomophone discrimination; PHO, PC of nonword reading and Spoonerisms; MOT, FM2, and FM240 as in Table 2. **, P ≤ 0.01.
Discussion
Among other possibilities, our results are consistent with the hypothesis that sensitive visual magnocellular processing is important for orthographic aspects of single-word reading. The magnocellular retino-cortical pathway has been shown to be important for visual search (61) and for the accurate letter position encoding necessary for skilled reading (43, 44). This pathway has been identified as one prominent locus of the visual deficit associated with developmental dyslexia (62, 63). The association between reading disability and poor visual magnocellular function raises the possibility that failure to visually process print correctly contributes directly to reading difficulties (43, 44). Conversely, high magnocellular sensitivity may be associated with better reading skills. Our data provide support for these hypotheses; we found that measures of normal children's orthographic skill covaried with their coherent motion threshold. At least three mechanisms can be proposed to account for this relationship. First, magnocellular input to the dorsal visual stream is considered to be important for encoding information about the spatial position of objects (64, 65). Thus, magnocellular sensitivity may influence representations of the spatial position of letters with respect to one another. Children who are good motion detectors are less likely to mistake anagrams for real words in a word anagram (e.g., “ocean” vs. “oaecn”) decision task (43). Second, magnocellular function seems to influence selective attention in visual search (61, 66, 67), which has been found to be impaired in both dyslexic children (68) and adults (61). Third, orthographic information has been suggested to play an important role in programming saccadic eye movements during reading (69). The magnocellular pathway provides the main visual input to neural structures important for eye movement control (for review see ref. 38). Poor fixation stability or poor eye movement control therefore would be expected to degrade the orthographic information that is available in the parafovea immediately preceding a saccade.
It has been suggested that visual magnocellular processing also affects nonword reading skill, a sensitive measure of phonological ability (20, 39, 41–43). Cestnick and Coltheart (45) suggested that magnocellular visual processing would affect nonword reading more than exception words because the former might need to be read letter by letter because there is no top-down information about where the letters should be localized. Although we found that motion sensitivity did correlate with nonword reading to a similar degree as that found in previous studies (21, 45–47) (Table 2), we argue that the magnocellular contribution to exception word reading should be the stronger because these words cannot be read by phonological processes alone, i.e., extra visual information is needed to solve the orthographic irregularity presented by exception words. The relationships between deficient phonological skill and poor magnocellular visual processing in dyslexic adults that have been found (70, 71) therefore may result from the severity of their reading deficit.
Like the relationship between dynamic visual processing and orthographic skill, we found that auditory sensitivity to 2-Hz FM, which requires tracking dynamic changes in frequency, was a significant predictor of children's phonological decoding skills. However, this relationship did not hold as strongly for detection of 240-Hz FM, which is probably detected by a separate auditory mechanism. At 2-Hz FM, subjects can track the changes in the pitch of the sound in real time, and detection is likely to be mediated by temporally sensitive mechanisms, probably at a cortical level. However, the perception of 240-Hz FM probably depends on spectral cues because the side bands in the modulation spectrum fall outside the bandwidth of the peripheral auditory filters (72, 73). Subjects hear a tone at the pitch of the modulating frequency (a component not present in the physical spectrum).
Highly accurate processing of temporal change by the auditory system is therefore likely to be important for successful development of phonological skills. Segmentation of words into their constituent phonemes is required to match them with their written representations for grapheme-to-phoneme translation. Thus phonological analysis draws on auditory representations of the sounds that each letter stands for. These phonetic contrasts are signaled by changes in sound frequency and amplitude, which are processed by the auditory system. Because the 2-Hz and 240-Hz FM tests were administered identically, this finding suggests that detection of the slower modulations in speech are necessary for speech perception and therefore for the development of phonological skill. These results are consistent with other findings that showed that poor readers have a developmental delay in acquiring sensitivity to the rhythm in speech (74) and that the effects of speech perception on reading are likely to be mediated by phonological awareness (75).
Impaired processing of FM in sound has been demonstrated in some other neurological conditions associated with impairments of receptive language (76, 77) and in dyslexia (ref. 21, cf. ref. 78). Although there is no such clearly identifiable subsystem of auditory magnocells as that seen in the visual system, there are magno-like divisions of each of the auditory relay nuclei (79). These neurons are especially important for processing the frequency and amplitude changes that signal phonetic contrasts (80). Moreover, as for the visual system, we have shown that good readers have high auditory sensitivity, as indexed by FM (21). This correlation between auditory sensitivity and literacy skills across the whole range of reading abilities suggests that auditory processing could constrain the development of the phonological skills required for successful literacy development. Thus these data suggest that dynamic visual and auditory sensitivity are likely to play an important role in the development of the fine-grained orthographic and phonological representations necessary for successful reading. Although these correlations do not prove a causal relationship between these variables, together these results provide direct evidence for a link between basic temporal sensory processing and lexical decoding skills in normal readers
Conclusions
Our results can be summarized as follows. Measures of auditory and visual temporal sensitivity were well correlated with measures of phonological and orthographic skill, respectively, in our sample of unselected 10-year-old readers. These relationships were clarified by removing the effects of intelligence and reading; then auditory FM sensitivity at 2 Hz but not at 240 Hz strongly predicted phonological ability but not orthographic skill, whereas visual motion sensitivity predicted orthographic skill but not phonological ability. Although these correlations do not prove a causal relationship, it seems likely that low-level visual and auditory function play important and independent roles in determining children's ability to learn to read. These effects are likely to be indirect, mediated by constraining the phonological and orthographic processing engaged during lexical access.
Our results therefore provide support for Tallal's hypothesis (19, 20) that poor phonological processing skills can result from deficient neurological processing of rapidly presented or rapidly changing acoustic stimuli. However, selective deficits for lower FM rates in dyslexic readers (21) coupled with the current finding of substantial covariance between the phonological skill of normal readers and 2-Hz FM suggests that the temporal changes within a stimulus may be the crucial parameter.
Acknowledgments
We acknowledge the generous support of the students, teachers, and parents associated with the Meltham Primary School. Richard Olson and Helen Datta provided information about the pseudohomophone task. This work was supported by the Wellcome Trust, the Rodin Remediation Foundation, and the Medical Research Council/Interdisciplinary Research Centre for Cognitive Neuroscience.
Abbreviation
- BAS
British Abilities Scales
Footnotes
Article published online before print: Proc. Natl. Acad. Sci. USA, 10.1073/pnas.040546597.
Article and publication date are at www.pnas.org/cgi/doi/10.1073/pnas.040546597
References
- 1.Gough P, Walsh M. In: Phonological Processes in Literacy: A Tribute to Isabelle Y. Liberman. Brady S, Shankweiler D, editors. Hillsdale, NJ: Lawrence Erlbaum; 1991. pp. 199–209. [Google Scholar]
- 2.Stanovich K, West R, Cunningham A. In: Phonological Processes in Literacy: A Tribute to Isabelle Y. Liberman. Brady S, Shankweiler D, editors. Hillsdale, NJ: Lawrence Erlbaum; 1991. pp. 219–235. [Google Scholar]
- 3.Olson R, Forsberg H, Wise B, Rack J. In: Frames of Reference for the Assessment of Learning Disabilities: New Views on Measurement Issues. Lyon G R, editor. Baltimore: Paul H. Brookes; 1994. pp. 243–277. [Google Scholar]
- 4.Mason M. J Exp Psychol Gen. 1975;104:146–166. [Google Scholar]
- 5.Gibson E, Shurcliff A, Yonas A. In: Basic Studies of Reading. Levin H, Williams J, editors. New York: Basic Books; 1970. pp. 57–73. [Google Scholar]
- 6.Massaro D, Venezky R, Taylor G. J Exp Psychol Gen. 1979;108:107–124. [PubMed] [Google Scholar]
- 7.Goswami U. Q J Exp Psychol. 1988;40A:239–268. [Google Scholar]
- 8.Jacobs A, Grainger J. J Exp Psychol Hum Percep Perf. 1994;20:1311–1334. [Google Scholar]
- 9.Brady S, Shankweiler D. Phonological Processes in Literacy: A Tribute to Isabelle Y. Liberman. Hillsdale, NJ: Lawrence Erlbaum; 1991. [Google Scholar]
- 10.Mattingly I. In: Language by Ear and by Eye: The Relationships Between Speech and Reading. Kavanagh J, Mattingly I, editors. Cambridge, MA: MIT Press; 1972. pp. 133–148. [Google Scholar]
- 11.Bradley L, Bryant P. Nature (London) 1978;271:746–747. doi: 10.1038/271746a0. [DOI] [PubMed] [Google Scholar]
- 12.Olson R, Wise B, Conners F, Rack J, Fulker D. J Learn Disabil. 1989;22:339–348. doi: 10.1177/002221948902200604. [DOI] [PubMed] [Google Scholar]
- 13.Gallagher A, Frederickson N. Educ Child Psychol. 1995;12:53–67. [Google Scholar]
- 14.Pennington B, Van Orden G, Smith S, Green P, Haith M. Child Dev. 1990;61:1753–1778. [PubMed] [Google Scholar]
- 15.Stanovich K. J Learn Disabil. 1988;21:590–612. doi: 10.1177/002221948802101003. [DOI] [PubMed] [Google Scholar]
- 16.Bradley L, Bryant P. Nature (London) 1983;301:419–421. [Google Scholar]
- 17.Studdert-Kennedy M, Mody M. Psychon Bull Rev. 1995;2:508–514. doi: 10.3758/BF03210986. [DOI] [PubMed] [Google Scholar]
- 18.Mody M, Studdert-Kennedy M, Brady S. J Exp Child Psychol. 1997;64:199–231. doi: 10.1006/jecp.1996.2343. [DOI] [PubMed] [Google Scholar]
- 19.Tallal P. Brain Lang. 1980;9:182–198. doi: 10.1016/0093-934x(80)90139-x. [DOI] [PubMed] [Google Scholar]
- 20.Tallal P, Merzenich M, Miller S, Jenkins W. Exp Brain Res. 1998;123:210–219. doi: 10.1007/s002210050563. [DOI] [PubMed] [Google Scholar]
- 21.Witton C, Talcott J, Hansen P, Richardson A, Griffiths T, Rees A, Stein J, Green G. Curr Biol. 1998;8:791–797. doi: 10.1016/s0960-9822(98)70320-3. [DOI] [PubMed] [Google Scholar]
- 22.Adams M, Huggins A. Reading Res Q. 1985;20:262–281. [Google Scholar]
- 23.Frith U. In: Surface Dyslexia. Patterson K, Marshall J, Coltheart M, editors. Hillsdale, NJ: Lawrence Erlbaum; 1985. pp. 301–330. [Google Scholar]
- 24.Conners F, Olson R. In: Comprehension Processes in Reading. Balota D, Flores d'Arcais G, Rayner K, editors. Hillsdale, NJ: Lawrence Erlbaum; 1990. pp. 557–579. [Google Scholar]
- 25.Castles A, Datta H, Gayan J, Olson R. J Exp Child Psychol. 1999;72:73–94. doi: 10.1006/jecp.1998.2482. [DOI] [PubMed] [Google Scholar]
- 26.Castles A, Coltheart M. Cognition. 1993;47:149–180. doi: 10.1016/0010-0277(93)90003-e. [DOI] [PubMed] [Google Scholar]
- 27.Boder W. Dev Med Child Neurol. 1973;15:663–687. doi: 10.1111/j.1469-8749.1973.tb05180.x. [DOI] [PubMed] [Google Scholar]
- 28.Ehri L. In: Reading Acquisition. Gough P, Ehri L, Treiman R, editors. Hillsdale, NJ: Lawrence Erlbaum; 1992. pp. 107–143. [Google Scholar]
- 29.Van Orden G. Memory Cog. 1987;15:181–198. doi: 10.3758/bf03197716. [DOI] [PubMed] [Google Scholar]
- 30.Seidenberg M, McClelland J. Psychol Rev. 1989;96:523–568. doi: 10.1037/0033-295x.96.4.523. [DOI] [PubMed] [Google Scholar]
- 31.Goswami U. J Exp Child Psychol. 1986;42:72–83. [Google Scholar]
- 32.Tunmer W, Nesdale A. J Ed Psychol. 1985;77:417–427. [Google Scholar]
- 33.Juel C, Griffith P, Gough P. J Ed Psychol. 1986;78:243–255. [Google Scholar]
- 34.Ball E, Blachman B. Reading Res Q. 1991;26:49–66. [Google Scholar]
- 35.Tangel D, Blachman B. J Reading Behavior. 1995;27:153–185. [Google Scholar]
- 36.Hatcher P, Hulme C, Ellis A. Child Dev. 1994;65:41–57. [Google Scholar]
- 37.Stein J, Talcott J. Dyslexia. 1999;5:59–77. [Google Scholar]
- 38.Stein J, Walsh V. Trends Neurosci. 1997;20:147–152. doi: 10.1016/s0166-2236(96)01005-3. [DOI] [PubMed] [Google Scholar]
- 39.Lovegrove W, Martin F, Slaghuis W. Cognit Neuropsychol. 1986;3:225–267. [Google Scholar]
- 40.Farmer M, Klein R. Psychon Bull Rev. 1995;2:460–493. doi: 10.3758/BF03210983. [DOI] [PubMed] [Google Scholar]
- 41.Reed M. J Exp Child Psychol. 1989;48:270–292. doi: 10.1016/0022-0965(89)90006-4. [DOI] [PubMed] [Google Scholar]
- 42.Adlard A, Hazan V. Q J Exp Psychol A. 1998;51:153–177. doi: 10.1080/713755750. [DOI] [PubMed] [Google Scholar]
- 43.Cornelissen P, Hansen P, Hutton J, Evangelinou V, Stein J. Vision Res. 1998;38:471–482. doi: 10.1016/s0042-6989(97)00199-5. [DOI] [PubMed] [Google Scholar]
- 44.Cornelissen P, Hansen P, Gilchrist I, Cormack F, Essex J, Frankish C. Vision Res. 1998;38:2181–2191. doi: 10.1016/s0042-6989(98)00016-9. [DOI] [PubMed] [Google Scholar]
- 45.Cestnick L, Coltheart M. Cognition. 1999;71:231–255. doi: 10.1016/s0010-0277(99)00023-2. [DOI] [PubMed] [Google Scholar]
- 46.Slaghuis W, Twell A, Kingston K. Cortex. 1996;32:413–438. doi: 10.1016/s0010-9452(96)80002-5. [DOI] [PubMed] [Google Scholar]
- 47.Talcott J, Hansen P, Willis-Owen C, McKinnell I, Richardson A, Stein J. Neuro-ophthalmology. 1998;20:187–201. [Google Scholar]
- 48.Elliot C, Murray D, Pearson L. British Abilities Scales. Windsor, U.K.: National Foundation for Educational Research-Nelson; 1983. [Google Scholar]
- 49.Olson R, Forsberg H, Wise B. In: The Varieties of Orthographic Knowledge 1: Theoretical and Development Issues. Berninger V, editor. Dordrecht, The Netherlands: Kluwer; 1994. pp. 27–71. [Google Scholar]
- 50.Macmillan N, Creelman C. Detection Theory: A Users Guide. Cambridge, U.K.: Cambridge Univ. Press; 1991. [Google Scholar]
- 51.Wattam-Bell J. Vision Res. 1994;34:877–883. doi: 10.1016/0042-6989(94)90038-8. [DOI] [PubMed] [Google Scholar]
- 52.Kaernbach C. Percept Psychophys. 1991;49:227–229. doi: 10.3758/bf03214307. [DOI] [PubMed] [Google Scholar]
- 53.Hirsh I, Watson C. Annu Rev Psychol. 1996;47:461–484. doi: 10.1146/annurev.psych.47.1.461. [DOI] [PubMed] [Google Scholar]
- 54.Coltheart M, Leahy J. Aus J Psychol. 1996;48:136–140. [Google Scholar]
- 55.Morais J, Bertelson P, Cary L, Alegria J. Cognition. 1979;7:323–331. [Google Scholar]
- 56.Wagner R, Torgeson J, Rashotte C. Dev Psychol. 1994;30:73–87. [Google Scholar]
- 57.Shankweiler D, Liberman I, Mark L, Fowler C, Fischer F. J Exp Psychol Hum Learn Mem. 1979;5:531–545. [Google Scholar]
- 58.Newsome W, Paré E. J Neurosci. 1988;8:2201–2211. doi: 10.1523/JNEUROSCI.08-06-02201.1988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Britten K, Shadlen M, Newsome W, Movshon J. J Neurosci. 1992;12:4745–4765. doi: 10.1523/JNEUROSCI.12-12-04745.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Newsome W, Britten K, Movshon J. Nature (London) 1989;341:52–54. doi: 10.1038/341052a0. [DOI] [PubMed] [Google Scholar]
- 61.Vidyasagar T, Pammer K. NeuroReport. 1999;10:1283–1287. doi: 10.1097/00001756-199904260-00024. [DOI] [PubMed] [Google Scholar]
- 62.Eden G, Van Meter J, Rumsey J, Maisog J, Woods R, Zeffiro T. Nature (London) 1996;382:66–69. doi: 10.1038/382066a0. [DOI] [PubMed] [Google Scholar]
- 63.Demb J, Boynton G, Heeger D. Proc Natl Acad Sci USA. 1997;94:13363–13366. doi: 10.1073/pnas.94.24.13363. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Milner D, Goodale M. The Visual Brain in Action. Oxford, U.K.: Oxford Univ. Press; 1995. [Google Scholar]
- 65.Mishkin M, Ungerlieder L, Macko K. Trends Neurosci. 1983;6:414–417. [Google Scholar]
- 66.Steinman B, Steinman S, Lehmkuhle S. Vision Res. 1997;37:17–23. doi: 10.1016/s0042-6989(96)00151-4. [DOI] [PubMed] [Google Scholar]
- 67.Vidyasagar T. NeuroReport. 1998;9:1947–1952. doi: 10.1097/00001756-199806220-00006. [DOI] [PubMed] [Google Scholar]
- 68.Brannan J, Williams M. Percept Psychophys. 1987;41:23–28. doi: 10.3758/bf03208209. [DOI] [PubMed] [Google Scholar]
- 69.Beauvillain C, Doré K. Vision Res. 1998;38:115–123. doi: 10.1016/s0042-6989(97)00127-2. [DOI] [PubMed] [Google Scholar]
- 70.Borsting E, Ridder III W, Dudeck K, Kelly C, Matsui L, Motoyama J. Vision Res. 1996;36:1047–1053. doi: 10.1016/0042-6989(95)00199-9. [DOI] [PubMed] [Google Scholar]
- 71.Ridder III W, Borsting E, Cooper M, McNeel B, Huang E. Optom Vis Sci. 1997;74:99–104. doi: 10.1097/00006324-199702000-00021. [DOI] [PubMed] [Google Scholar]
- 72.Whitfield I, Evans E. J Neurophysiol. 1965;28:655–672. doi: 10.1152/jn.1965.28.4.655. [DOI] [PubMed] [Google Scholar]
- 73.Kay R. Physiol Rev. 1982;63:894–975. doi: 10.1152/physrev.1982.62.3.894. [DOI] [PubMed] [Google Scholar]
- 74.Wood C, Terrell C. Br J Dev Psychol. 1998;16:397–413. [Google Scholar]
- 75.McBride-Chang C. Child Dev. 1996;67:1836–1856. [PubMed] [Google Scholar]
- 76.Stefanatos G, Green G, Ratcliff G. Arch Neurol. 1989;46:871–875. doi: 10.1001/archneur.1989.00520440053021. [DOI] [PubMed] [Google Scholar]
- 77.Stefanotos G. Ann NY Acad Sci. 1993;682:412–414. doi: 10.1111/j.1749-6632.1993.tb23009.x. [DOI] [PubMed] [Google Scholar]
- 78.Adlard A, Hazan V. Q J Exp Psychol. 1998;51A:153–177. doi: 10.1080/713755750. [DOI] [PubMed] [Google Scholar]
- 79.Kaas J, Hackett T, Tramo M. Curr Opin Neurobiol. 1999;9:164–170. doi: 10.1016/s0959-4388(99)80022-1. [DOI] [PubMed] [Google Scholar]
- 80.Rauschecker J. Curr Opin Neurobiol. 1998;8:516–521. doi: 10.1016/s0959-4388(98)80040-8. [DOI] [PubMed] [Google Scholar]