

Abstract
Identifying translocations of the MLL gene at chromosome band 11q23 is important for the characterization and treatment of leukemia. However, cytogenetic analysis does not always find the translocations and the many partner genes of MLL make molecular detection difficult. We developed cDNA panhandle PCR to identify der(11) transcripts regardless of the partner gene. By reverse transcribing first-strand cDNAs with oligonucleotides containing coding sequence from the 5′ MLL breakpoint cluster region at the 5′ ends and random hexamers at the 3′ ends, known MLL sequence was attached to the unknown partner sequence. This enabled the formation of stem-loop templates with the fusion point of the chimeric transcript in the loop and the use of MLL primers in two-sided PCR. The assay was validated by detection of the known fusion transcript and the transcript from the normal MLL allele in the cell line MV4–11. cDNA panhandle PCR then was used to identify the fusion transcripts in two cases of treatment-related acute myeloid leukemia where the karyotypes were normal and the partner genes unknown. cDNA panhandle PCR revealed a fusion of MLL with AF-10 in one case and a fusion of MLL with ELL in the other. Alternatively spliced transcripts and exon scrambling were detectable by the method. Leukemias with normal karyotypes may contain cryptic translocations of MLL with a variety of partner genes. cDNA panhandle PCR is useful for identifying MLL translocations and determining unknown partner sequences in the fusion transcripts.
Translocations of the MLL gene at chromosome band 11q23 occur in leukemias of infants (reviewed in ref. 1) and leukemias associated with DNA topoisomerase II inhibitors (reviewed in ref. 2). The ability to rapidly identify MLL translocations, whether by cytogenetic or molecular approaches, is relevant to diagnosis and prognosis and to treatment planning. MLL is an example of a gene involved in translocations with numerous different partner genes; many are still uncharacterized (1, 2). The specific partner gene with which MLL is fused may have an impact on the clinical response (3). Previously, we developed panhandle PCR approaches to identify MLL der(11) translocation breakpoints in genomic DNA (4–8). The salient features include attachment of known MLL sequence to the unknown partner gene, formation of a stem-loop template, and two-sided PCR. Because MLL sequences are at both ends of the template, all primers are derived from MLL. Panhandle PCR methods offer the advantage of amplifying MLL translocation breakpoints without primers from the partner genes.
Although panhandle PCR approaches are highly effective for genomic translocation breakpoint cloning (4–8), sometimes the genomic target sequence may be too large to amplify. In addition, if the amplicon contains intronic sequence only and not exonic sequence, panhandle PCR approaches may not reveal the partner gene (4, 8). MLL genomic translocation breakpoints occur within an 8.3-kb breakpoint cluster region (bcr) (9). Here, we targeted the corresponding 859-bp cDNA (9) for a panhandle PCR approach that would identify der(11) transcripts. We used cDNA panhandle PCR to investigate the nature of the MLL fusion transcripts in two cases of treatment-related acute myeloid leukemia (AML) with normal karyotypes but with MLL gene rearrangement detectable by Southern blot. Leukemias with normal karyotypes have been found to harbor MLL tandem duplications (6, 10–17). The results of this analysis show that leukemias with normal karyotypes may harbor MLL gene translocations. cDNA panhandle PCR can lead readily to discovery of partner genes.
Methods
The institutional review boards at Children's Hospital Los Angeles, Vanderbilt University Medical Center, and The Children's Hospital of Philadelphia approved this research. The AML cell line MV4–11 with known t(4;11) (18) and two cases of treatment-related AML (t-AML) (Table 1) were studied. Genomic DNA and total RNA were isolated from MV4–11 cells and cryopreserved leukemic marrow cells by using 5.7 M CsCl-4 M guanidine isothiocyanate gradients. MLL gene rearrangement was examined by using the B859 fragment of ALL-1 cDNA (19).
Table 1.
Cases of t-AML
| Patient t-36 | Patient t-44 | |
|---|---|---|
| Race/gender | Hispanic/male | White/male |
| Primary cancer | Alveolar rhabdomyosarcoma | Ewing's sarcoma |
| Age at primary cancer diagnosis | 1 yr, 4 months | 10 yrs, 8 months |
| Prior chemotherapy | VCR, VP16, IFOS | VCR, VP16, IFOS, ADR, CPM, AMD, CBDCA |
| Prior radiation | No | No |
| Interval to t-AML | 60 months | 34 months |
| FAB | M4 | M2 |
| Karyotype | 46,XY | 46,XY |
VCR indicates vincristine; VP16, etoposide; IFOS, ifosphamide; ADR, doxorubicin; CPM, cyclophosphamide; AMD, dactinomycin; CBDCA, carboplatin; FAB, French-American-British.
cDNA Panhandle PCR Analysis of Normal MLL and der(11) Transcripts.
Fig. 1 summarizes cDNA panhandle PCR. In step 1, a population of first-strand cDNAs is reverse-transcribed from total RNA with oligonucleotides containing known MLL coding sequence from the 5′ bcr at the 5′ ends and random hexamers at the 3′ ends. The random hexamers anneal to and prime from many complementary sequences in total RNA. This produces first-strand cDNAs of different sizes. Those derived from the normal MLL allele and from der(11) transcripts have a known MLL sequence and its inverse complement at the 5′ and 3′ ends, respectively (Fig. 1 Upper Left). The purpose of step 2 is to generate sense second strands by MLL primer 1 extension from the first-strand cDNAs (Fig. 1 Upper Center). In step 3, primer 2 is added and thermal cycling is begun. The first event during thermal cycling is formation of stem-loop “panhandle” templates from the second strands. Heat denaturation makes the double-stranded cDNAs single-stranded. Intrastrand annealing of complementary MLL sequences in the second strands and polymerase extension of the recessed 3′ ends complete formation of the handles. Either normal MLL sequence or the point of fusion in the chimeric transcripts and coding sequence of the partner gene are within the loops (Fig. 1 Upper Right). Step 4 continues with primer 1 extension from the 3′ end of the stem-loop structure during the thermal cycling. This generates the double-stranded template with MLL sequence at both ends to be amplified exponentially with MLL primers 1 and 2 (Fig. 1 Lower Left). Step 5 entails nested PCR with MLL primers 3 and 4 (Fig. 1 Lower Right). Positions of the MLL primers relative to the MLL sequence in the oligonucleotides used to generate the first-strand cDNAs are shown in Fig. 1.
Figure 1.
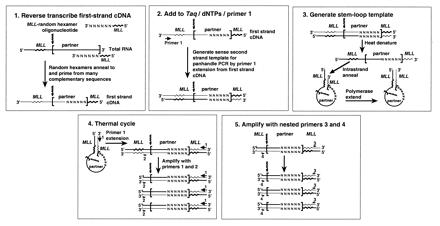
Steps in cDNA panhandle PCR. Corkscrew arrow indicates fusion point in MLL chimeric transcript. A population of first-strand cDNAs of various sizes is expected because of MLL-random hexamer oligonucleotide design. Transcripts from normal MLL allele also would be amplified.
The specific protocol follows. First-strand cDNAs were synthesized from 1 μg of total RNA by using the Superscript Preamplification System (Life Technologies, Grand Island, NY), except that MLL-random hexamer oligonucleotides were used in place of random hexamers. The oligonucleotide sequences were 5′-CCTGAATCCAAACAGGCCACCACTCCAGCTTC-NNNNNN-3′ (8); the MLL sequence in the oligonucleotides corresponded to bcr cDNA positions 92–123 in exon 5 (8, 9). Two microliters of first-strand cDNAs was added to a 45.5 μl mixture containing 1.75 units Taq/Pwo DNA polymerase, 385 μM each dNTP, 1.1× buffer (Expand Long Template System, Boehringer Mannheim), and 12.5 pmols of MLL primer 1 (8). The mixture was heated to 80°C for 5 min before the cDNAs were added. Primer 1 corresponded to MLL bcr cDNA positions 34–55 in exon 5 (5′-TCCTCCACGAAAGCCCGTCGAG-3′) (8, 9), upstream of the MLL sequence in the oligonucleotides used to synthesize the first-strand cDNAs. To achieve primer 1 extension, the mixture was denatured at 94°C for 1 min followed by 1 cycle at 94°C for 10 sec and 68°C for 7 min. The sample was heated again to 80°C for 5 min and 2.5 μl (12.5 pmols) of MLL primer 2 were added. Primer 2 corresponded to MLL bcr cDNA positions 136–158 in exon 5 (5′-TCAAGCAGGTCTCCCAGCCAGCAC-3′) (8, 9), downstream of the MLL sequence in the oligonucleotides used to synthesize the first-strand cDNAs. The final 50-μl PCRs contained 350 μM each dNTP and 1× buffer. PCR with primers 1 and 2, including the 7-min elongation and the increment in elongation times, was as described (4, 5, 8). One microliter of the products was used in nested PCR with primers 3 (5′-GGAAAAGAGTGAAGAAGGGAATGTCTCGG-3′) and 4 (5′-GTGGTCATCCCGCCTCAGCCAC-3′) corresponding to MLL bcr cDNA positions 55–83 and 159–179 in exon 5 (8, 9). Conditions were the same as above.
cDNA panhandle PCR products were subcloned by recombination PCR (7). pUC19 was linearized by HindIII digestion. MLL ends complementary to the ends of the cDNA panhandle PCR products that were to be inserted were added to the vector during PCR using primers 5′-ACATTCCCTTCTTCACTCTTTTCCTGGCGTAATCATGGTCATAGC-3′ and 5′-GTGGCTGAGGCGGGATGACCACCATGCCTGCAGGTCGACTC-3′ (8) and described conditions (7). The PCR-amplified pUC19 and cDNA panhandle PCR products were purified by using Geneclean III reagents and resuspended in 10 μl of elution buffer (Bio 101). PCR-amplified, purified pUC19 (2.5 μl) and 2.5 μl of purified cDNA panhandle PCR products were mixed and added to 50 μl of MAX efficiency DH5α cells (Life Technologies) to recombine in vivo. The transformation was per manufacturer's instructions (Life Technologies). Subclones containing cDNA panhandle PCR products were identified by PCR with primers 3 and 4 (8).
Validation of der(11) Transcripts.
The above-generated first-strand cDNAs were amplified with gene-specific primers. For the t-AML of patient 36, the sense primer 5′-CGCCCAAGTATCCCTGTAAA-3′ from MLL exon 6 or 5′-TGAAGCAGAAAATGTGTGGG-3′ from MLL exon 8 was used with the antisense primer 5′-GATTTCTTCCCTTTGCCCTC-3′ from AF-10. For the t-AML of patient 44, the sense primer 5′-AGTGAGCCCAAGAAAAAG-3′ from MLL exon 5 was used with the antisense primer 5′-GCCGATGTTGGAGAGGTAGA-3′ from ELL.
Results
cDNA Panhandle PCR Detects the Known der(11) Transcript in MV4–11 Cell Line.
To test cDNA panhandle PCR as a strategy for cloning der(11) transcripts, the cell line MV4–11 with a known transcript fusing MLL exon 6 to AF-4 exon 5 (GenBank accession nos. L04284 and NM_005933) (20, 21) was studied. cDNA panhandle PCR products of various sizes were obtained (Fig. 2A). Recombination PCR gave 50 subclones, 12 of which were sequenced (Fig. 2B). Two subclones with 1,589-bp inserts contained MLL exons 5–17, suggesting that they were from transcripts from the normal MLL allele. The expected in-frame fusion of MLL exon 6 to AF-4 exon 5 (20, 21) was present in eight of the 12 subclones. The insert sizes that contained this junction were from 381 to 2,904 bp; all but one were >1,565 bp and three were >2 kb. Because the MLL bcr cDNA sequence is only 859 bp (9), it was possible to identify the point of fusion in the 381-bp subclone. The sequences of two subclones were consistent with vector without insert.
Figure 2.
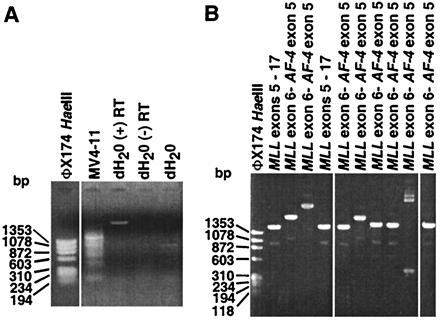
(A) cDNA panhandle PCR analysis of total RNA from cell line MV4–11. Smear indicates products of various sizes (lane MV4-11). dH2O control reactions with and without reverse transcriptase (RT) were included. (B) PCR screen of recombination PCR-generated subclones of cDNA panhandle PCR products from cell line MV4–11. MLL-containing subclones shown by sequencing are indicated above respective lanes. Two subclones contained normal MLL cDNA; eight showed in-frame fusion of MLL exon 6 to position 148 of 2,805-bp full-length AF-4 cDNA (GenBank accession no. L13773).
cDNA Panhandle PCR Reveals a Cryptic t(10,11) Translocation in t-AML with Normal Karyotype.
The karyotype was normal in the t-AML of patient 36 (Table 1) but 14.5- and 4.2-kb rearrangements on Southern blot analysis were consistent with a cryptic translocation involving MLL (Fig. 3). We used cDNA panhandle PCR to ascertain the partner gene. cDNA panhandle PCR products of various sizes were obtained (Fig. 4A). Recombination PCR gave 261 subclones (Fig. 4B); 25 were sequenced. Of the 25, 12 contained MLL sequence only, 12 revealed the unknown partner gene, and one was empty vector.
Figure 3.
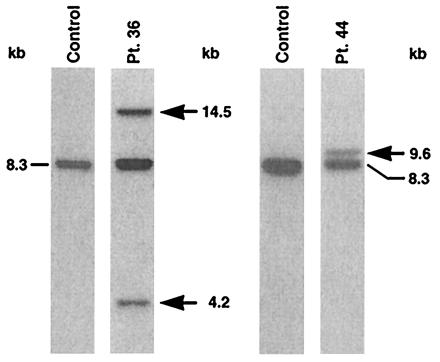
Identification of MLL rearrangements in cases of t-AML by Southern blot analysis. BamHI-digested marrow DNAs were hybridized with B859 fragment of ALL-1 cDNA (9). Peripheral blood lymphocyte DNA from normal subject was used as the control. Dash shows germ-line band; arrows show rearrangements. Patient numbers correspond to text and Table 1.
Figure 4.
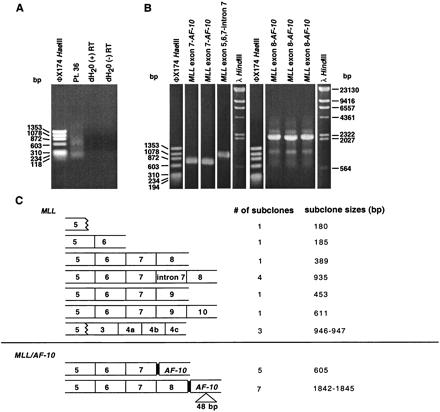
(A) cDNA panhandle PCR analysis of total RNA from t-AML of patient 36. Smear indicates products of heterogeneous sizes (lane Pt. 36). (B) PCR screen of representative recombination PCR-generated subclones of cDNA panhandle PCR products shown in A. MLL-containing subclones confirmed by sequencing are shown above respective lanes. (C) Summary of sequences in recombination PCR-generated subclones. Twelve subclones contained MLL sequence only (Upper). Incompletely processed and alternatively spliced transcripts and transcripts suggesting exon scrambling were detected. In three subclones, exon 5 sequence extended to position 3945 of MLL cDNA; the 3′ sequence was from exons 3, 4a, 4b, and 4c starting at position 2782 of MLL cDNA (GenBank accession nos. LO4284 and NM_005933). Positions 3945 and 2782 are the end and start of internal codons in MLL exons 5 and 3. Chimeric transcripts contained in-frame fusions of either MLL exon 7 or MLL exon 8 to position 979 of AF-10 cDNA (GenBank accession no. U13948) (Lower).
The subclones with MLL sequence only are summarized in Fig. 4C. The insert sizes were from 180 to 947 bp. These subclones were derived from fully processed transcripts, incompletely processed transcripts with intronic sequence, and alternatively spliced transcripts skipping exon 8. Transcripts fusing the end of an internal codon in exon 5 with the start of an internal codon in exon 3 suggested exon scrambling (22, 23).
Two in-frame chimeric transcripts were detected that fused either MLL exon 7 or MLL exon 8 to position 979 of the cDNA sequence of AF-10 (GenBank accession no. U13948) (Fig. 4C). Five subclones with 605-bp inserts contained the MLL exon 7-AF-10 junction. Seven subclones with 1,842- to 1,845-bp inserts contained the MLL exon 8-AF-10 junction. AF-10 is a known partner gene of MLL at chromosome band 10p12 (24, 25). In the subclones with the MLL exon 8-AF-10 junction, the 48 bases from positions 1883 to 1930 of the corresponding AF-10 cDNA, which are codons 567–582, were absent; the sequence then continued through AF-10 cDNA position 2379 in the largest subclone. A recent sequence entry (GenBank accession no. AL161799) containing intron-exon junctions for a portion of AF-10 suggests that the point of fusion in AF-10 is the first base of an exon. The same entry suggests that the 48 bases absent from the transcripts fusing MLL exon 8 to AF-10 may comprise an exon. These results indicate that both MLL and AF-10 were alternatively spliced.
Amplification of the same first-strand cDNA with MLL exon 6- and AF-10-specific primers and sequencing confirmed both fusion transcripts. Predicted 335-bp and 447-bp products contained the MLL exon 7-AF-10 and MLL exon 8-AF-10 junctions. Sequencing of a 268-bp product obtained with MLL exon 8- and AF-10-specific primers further confirmed the fusion of MLL exon 8 with AF-10.
cDNA Panhandle PCR Reveals a Cryptic t(11;19) Translocation in t-AML with Normal Karyotype.
The karyotype was also normal in the t-AML of patient 44 (Table 1), but Southern blot analysis of the MLL bcr revealed a single 9.6-kb rearrangement (Fig. 3). We used cDNA panhandle PCR to investigate the fusion transcript. The cDNA panhandle PCR products of various sizes are shown in Fig. 5A. Recombination PCR gave 130 subclones (Fig. 5B), 23 of which were sequenced.
Figure 5.
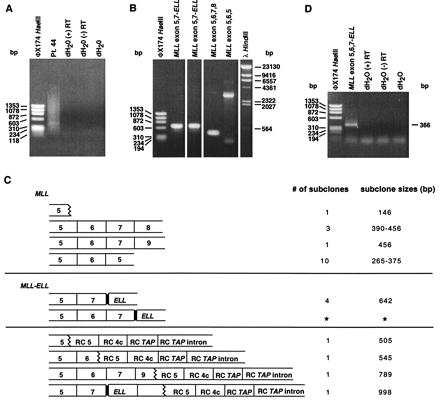
(A) cDNA panhandle PCR analysis of total RNA from t-AML of patient 44. Smear indicates products of heterogeneous sizes (lane Pt. 44). (B) PCR screen of representative recombination PCR-generated subclones of cDNA panhandle PCR products shown in A. MLL-containing subclones shown by sequencing are indicated above respective lanes. (C) Summary of sequences in recombination PCR-generated subclones. Fifteen subclones contained MLL sequence alone (Top). Alternatively spliced and scrambled transcripts were detected. Alternatively spliced chimeric transcript identified by cDNA panhandle PCR in which MLL exon 6 was absent had in-frame fusion of MLL exon 7 to position 148 of ELL cDNA (GenBank accession no. U139480) (Middle). * shows second MLL-ELL chimeric transcript in which MLL exon 6 was present, which was identified by amplification of same first-strand cDNA with gene-specific primers (please see second lane, D). Transcripts with MLL or MLL-ELL and TAP sequences are unconfirmed (Bottom). RC indicates reverse complement. (D) Detection of second MLL-ELL chimeric transcript by PCR with gene-specific primers. Sequence of 366-bp product showed fusion of MLL exon 7 to ELL in transcript in which MLL exons 5–7 all were present (lane 2) (please see * in C).
Fifteen of the 23 subclones contained MLL sequence only and were from 146 to 456 bp (Fig. 5 B and C). One subclone ended within exon 5. Three subclones contained MLL exons 5–8. One subclone derived from an alternatively spliced transcript contained MLL exons 5, 6, 7, and 9, skipping exon 8 (Fig. 5D). Ten subclones contained MLL exons 5 and 6 and then exon 5 joined accurately at splice junctions, suggesting scrambled exons (22, 23).
Four subclones with 642-bp inserts revealed the unknown partner gene. Each contained a fusion of MLL exon 7 to position 148 of the cDNA sequence of ELL (GenBank accession no. U16282), a previously described partner gene of MLL at chromosome band 19p13 (26, 27). Because MLL exon 5 was joined to exon 7, skipping exon 6, this fusion transcript was alternatively spliced. Amplification of a 366-bp product with MLL exon 5- and ELL-specific primers and sequencing revealed a second chimeric transcript with the same MLL exon 7-ELL point of fusion, in which MLL exon 6 was present (Fig. 5 C and D).
Three subclones contained MLL sequence as well as the reverse complement of MLL sequence and the reverse complement of TAP sequence (tip-associated protein) (GenBank accession nos. U80073 and AC015703), which is also from chromosome band 11q23 (Fig. 5D). One subclone contained the alternatively spliced MLL exon 7-ELL fusion and the reverse complement of MLL and TAP sequence. The significance of these unusual subclones is unclear because PCR with gene-specific primers did not verify the corresponding transcripts.
Discussion
The original panhandle PCR and panhandle variant PCR amplify genomic DNA with known 5′ and unknown 3′ sequences (28, 29). Because MLL has many unknown partner genes, we previously adapted these methods to isolate MLL translocation breakpoints from genomic DNA (4–8). Both attach known MLL DNA to the unknown partner sequence. A stem-loop template forms and is amplified in two-sided PCR with primers all from MLL. In the present work, we developed cDNA panhandle PCR for MLL der(11) transcripts. The strategy has some features of the original and panhandle variant PCR approaches.
New strategies to detect translocations are essential because a growing number of genes in leukemia-associated translocations, including MLL, form fusions with multiple and/or uncharacterized translocation partners. Southern blot analysis identifies MLL translocations regardless of the partner gene, but Southern blot analysis is not as sensitive as PCR. Alternative approaches for detecting MLL translocations regardless of the partner gene are karyotype analysis and fluorescence in situ hybridization analysis with an MLL-specific probe; both have become routine tests when treatment-related leukemia is suspected so that, in many cases, such information is available. However, karyotype analysis detects only two-thirds of translocations involving MLL (30, 31) and, as was true in this study, has a high incidence of falsely negative results. Reverse transcriptase–PCR with gene-specific primers will not recognize all of the translocations because, for many of the partner genes, no primers are available.
Because the partner gene may be relevant to outcome (3), cDNA panhandle PCR is a simple approach to screening for MLL translocations that could be useful clinically. All three panhandle PCR-based strategies are well suited to MLL translocations. However, cDNA panhandle PCR has fewer steps, does not require restriction enzyme cleavage, and does not require a ligation. Moreover, in cDNA panhandle PCR, smaller amplicons contain the informative exonic sequences and reveal the partner genes more readily. The steps entail reverse transcription of first-strand cDNAs from total RNA using MLL-random hexamer oligonucleotides, generation of second-strand cDNAs by MLL primer 1 extension, formation of stem-loop templates by intrastrand annealing of the second strands, and PCR with primers all from MLL. The random hexamers in the oligonucleotides used to generate the first-strand cDNAs prime from many complementary sequences in the total RNA; however, only second-strand cDNAs containing MLL sequence at both ends will form the stem-loop structure that is amplified in the PCR. This increases specificity.
cDNA panhandle PCR is distinct from 3′ rapid amplification of cDNA ends (RACE), which generates first-strand cDNA using an oligonucleotide with a sequence complementary to the cloning vector and an oligo(dT) sequence at its 5′ and 3′ ends (32–34). The oligo(dT) sequence anneals to and primes from poly(A) tails in heterogeneous full-length transcripts. cDNA panhandle PCR seems advantageous over 3′ RACE PCR where larger products are required to reach the point of fusion. Another method creates the first-strand cDNA by reverse-transcribing with oligonucleotides containing complementary sequence to the cloning vector at the 5′ ends and random hexamers at the 3′ ends (24); however, the obligate intrastrand loop formation in generation of cDNA panhandle PCR templates confers greater specificity.
We used cDNA panhandle PCR to identify the unknown partner genes in two cases of t-AML in which the karyotypes were normal. The results indicate that cDNA panhandle PCR will identify heterogeneous MLL-containing transcripts from the normal and translocated MLL alleles in the same reaction. Fully spliced transcripts, alternatively spliced transcripts, transcripts with exon scrambling, and incompletely processed transcripts were detectable. The transcripts with MLL exons accurately spliced but scrambled and in an order different from genomic DNA indicate complexity in the splicing process; the phenomenon has been described (22, 23). The scrambled transcripts in the leukemia of patient 36 joined codons in the middle of two exons. Similarly, Caldas et al. (23) reported exon scrambling that resulted from splicing at nonsplice consensus sites within exons of the MLL gene. Additional experiments using cDNA panhandle PCR may augment our understanding of the heterogeneity of MLL-containing transcripts.
In one case of t-AML with two MLL rearrangements at the level of the Southern blot, the partner gene was AF-10 and two alternatively spliced transcripts were detected. The AF-10 gene is a known partner gene of MLL at chromosome band 10p12 and encodes a putative transcription factor as its protein product (24, 25). In the transcripts with the MLL exon 8-AF-10 junction, 48 bases of the contiguous AF-10 cDNA were absent. Chaplin et al. (25) observed absence of the same 48 bases from another MLL-AF-10 cDNA. The reading frame is not disrupted in these transcripts, and a recent AF-10 sequence entry suggests that the 48 bases comprise an exon. Thus, both MLL and AF-10 were alternatively spliced. In the second case, Southern blot analysis revealed a single MLL rearrangement. Although single rearrangements frequently are MLL tandem duplications (6, 10–17), cDNA panhandle PCR identified an MLL-ELL chimeric transcript. ELL is one of three known partner genes of MLL at chromosome band 19p13 and encodes an arginine-rich basic protein, the C terminus of which has homology to poly(ADP) ribose polymerase (26). Leukemias with normal karyotypes may harbor cryptic MLL translocations with a variety of partner genes.
Acknowledgments
C.A.F. is supported by National Institutes of Health Grants CA66140, CA80175, CA77683, and CA85469 and a Leukemia and Lymphoma Society Scholar Award.
Abbreviations
- AML
acute myeloid leukemia
- bcr
breakpoint cluster region
- t-AML
treatment-related AML
- TAP
tip-associated protein
Footnotes
Data deposition: The sequences reported in this paper have been deposited in the GenBank database (accession nos. AF272370–AF272385).
Article published online before print: Proc. Natl. Acad. Sci. USA, 10.1073/pnas.150241797.
Article and publication date are at www.pnas.org/cgi/doi/10.1073/pnas.150241797
References
- 1.Felix C A, Lange B J. Oncologist. 1999;4:225–240. [PubMed] [Google Scholar]
- 2.Felix C A. Biochim Biophys Acta. 1998;1400:233–255. doi: 10.1016/s0167-4781(98)00139-0. [DOI] [PubMed] [Google Scholar]
- 3.Reaman G H, Sposto R, Sensel M G, Lange B J, Feusner J H, Heerema N A, Leonard M, Holmes E J, Sather H N, Pendergrass T W, et al. J Clin Oncol. 1999;17:445–455. doi: 10.1200/JCO.1999.17.2.445. [DOI] [PubMed] [Google Scholar]
- 4.Felix C A, Kim C S, Megonigal M D, Slater D J, Jones D H, Spinner N B, Stump T, Hosler M R, Nowell P C, Lange B J, Rappaport E F. Blood. 1997;90:4679–4686. [PubMed] [Google Scholar]
- 5.Felix C A, Jones D H. Leukemia. 1998;12:976–981. doi: 10.1038/sj.leu.2401026. [DOI] [PubMed] [Google Scholar]
- 6.Megonigal M D, Rappaport E F, Jones D H, Kim C S, Nowell P C, Lange B J, Felix C A. Proc Natl Acad Sci USA. 1997;94:11583–11588. doi: 10.1073/pnas.94.21.11583. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Megonigal M D, Rappaport E F, Jones D H, Williams T M, Lovett B D, Kelly K M, Lerou P H, Moulton T, Budarf M L, Felix C A. Proc Natl Acad Sci USA. 1998;95:6413–6418. doi: 10.1073/pnas.95.11.6413. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Megonigal M D, Cheung N-K V, Rappaport E F, Nowell P C, Wilson R B, Jones D H, Addya K, Leonard D G B, Kushner B H, Williams T M, et al. Proc Natl Acad Sci USA. 2000;97:2814–2819. doi: 10.1073/pnas.050397097. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Gu Y, Nakamura T, Alder H, Prasad R, Canaani O, Cimino G, Croce C M, Canaani E. Cell. 1992;71:701–708. doi: 10.1016/0092-8674(92)90603-a. [DOI] [PubMed] [Google Scholar]
- 10.Schichman S A, Caligiuri M A, Gu Y, Strout M P, Canaani E, Bloomfield C D, Croce C M. Proc Natl Acad Sci USA. 1994;91:6236–6239. doi: 10.1073/pnas.91.13.6236. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Schichman S A, Caligiuri M A, Strout M P, Carter S L, Gu Y, Canaani E, Bloomfield C D, Croce C M. Cancer Res. 1994;54:4277–4280. [PubMed] [Google Scholar]
- 12.Caligiuri M A, Strout M P, Schichman S A, Mrozek K, Arthur D C, Herzig G P, Baer M R, Schiffer C A, Heinonen K, Knuutila S, et al. Cancer Res. 1996;56:1418–1425. [PubMed] [Google Scholar]
- 13.Caligiuri M A, Strout M P, Oberkircher A R, Yu F, de la Chapelle A, Bloomfield C D. Proc Natl Acad Sci USA. 1997;94:3899–3902. doi: 10.1073/pnas.94.8.3899. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.So C W, Ma Z G, Price C M, Dong S, Chen S J, Gu L J, So C K C, Wiedemann L M, Chan L C. Cancer Res. 1997;57:117–122. [PubMed] [Google Scholar]
- 15.Yu M, Honoki K, Andersen J, Paietta E, Nam D K, Yunis J J. Leukemia. 1996;10:774–780. [PubMed] [Google Scholar]
- 16.Wiedemann L M, MacGregor A, Caldas C. Br J Haematol. 1999;105:256–264. [PubMed] [Google Scholar]
- 17.Schnittger S, Kinkelin U, Schoch C, Heinecke A, Haase D, Haferlach T, Buchner T, Wormann B, Hiddemann W, Griesinger F. Leukemia. 2000;14:796–804. doi: 10.1038/sj.leu.2401773. [DOI] [PubMed] [Google Scholar]
- 18.Lange B, Valtieri M, Santoli D, Caracciolo D, Mavillio F, Gemperlein I, Griffin C, Emanuel B, Finan J, Nowell P, Rovera G. Blood. 1987;70:192–199. [PubMed] [Google Scholar]
- 19.Felix C A, Hosler M R, Winick N J, Masterson M, Wilson A E, Lange B J. Blood. 1995;85:3250–3256. [PubMed] [Google Scholar]
- 20.Reichel M, Gillert E, Nilson I, Siegler G, Greil J, Fey G H, Marschalek R. Oncogene. 1998;17:3035–3044. doi: 10.1038/sj.onc.1202229. [DOI] [PubMed] [Google Scholar]
- 21.Felix C A, Hosler M R, Slater D J, Megonigal M D, Lovett B D, Williams T M, Nowell P C, Spinner N B, Owens N L, Hoxie J, et al. Mol Diagn. 1999;4:269–283. doi: 10.1016/s1084-8592(99)80002-2. [DOI] [PubMed] [Google Scholar]
- 22.Nigro J M, Cho K R, Fearon E R, Kern S E, Ruppert J M, Oliner J D, Kinzler K W, Vogelstein B. Cell. 1991;64:607–613. doi: 10.1016/0092-8674(91)90244-s. [DOI] [PubMed] [Google Scholar]
- 23.Caldas C, So C W, MacGregor A, Ford A M, McDonald B, Chan L C, Wiedemann L M. Gene. 1998;208:167–176. doi: 10.1016/s0378-1119(97)00640-9. [DOI] [PubMed] [Google Scholar]
- 24.Chaplin T, Bernard O, Beverloo H B, Saha V, Hagemeijer A, Berger R, Young B D. Blood. 1995;86:2073–2076. [PubMed] [Google Scholar]
- 25.Chaplin T, Ayton P, Bernard O A, Saha V, Valle V D, Hillion J, Gregorini A, Lillington D, Berger R, Young B D. Blood. 1995;85:1435–1441. [PubMed] [Google Scholar]
- 26.Thirman M J, Levitan D A, Kobayashi H, Simon M C, Rowley J D. Proc Natl Acad Sci USA. 1994;91:12110–12114. doi: 10.1073/pnas.91.25.12110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Shilatifard A, Lane W S, Jackson K W, Conaway R C, Conaway J W. Science. 1996;271:1873–1876. doi: 10.1126/science.271.5257.1873. [DOI] [PubMed] [Google Scholar]
- 28.Jones D H, Winistorfer S C. Nucleic Acids Res. 1992;20:595–600. doi: 10.1093/nar/20.3.595. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Jones D H, Winistorfer S C. BioTechniques. 1997;23:132–138. doi: 10.2144/97231rr01. [DOI] [PubMed] [Google Scholar]
- 30.Felix C A, Hosler M R, Slater D J, Parker R, Masterson M, Whitlock J A, Rebbeck T R, Nowell P C, Lange B J. J Pediatr Hematol Oncol. 1998;20:299–308. doi: 10.1097/00043426-199807000-00004. [DOI] [PubMed] [Google Scholar]
- 31.Martinez-Climent J A, Thirman M J, Espinosa R, Le Beau M M, Rowley J D. Leukemia. 1995;9:1299–1304. [PubMed] [Google Scholar]
- 32.Frohman M A, Dush M K, Martin G R. Proc Natl Acad Sci USA. 1988;85:8998–9002. doi: 10.1073/pnas.85.23.8998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Frohman M A. In: PCR Protocols: A Guide to Methods and Applications. Innis M A, Gelfand D H, Sninsky J J, editors. San Diego: Academic; 1990. pp. 28–38. [Google Scholar]
- 34.Frohman M A. Methods Enzymol. 1993;218:340–356. doi: 10.1016/0076-6879(93)18026-9. [DOI] [PubMed] [Google Scholar]