

Abstract
Understanding the nature and relative importance of endogenous (density-dependent) and exogenous (density-independent) effects on population dynamics remains a central problem in ecology. Evaluation of these forces has been constrained by the lack of long time series of population densities and largely limited to populations chosen for their unique dynamics (e.g., outbreak insects). Especially in herbivore populations, the relative contributions of bottom-up and top-down effects (resources and natural enemies, respectively) have been difficult to compare because population data have rarely been combined with resource measurements. The feeding scars of a wood-mining herbivorous insect (Phytobia betulae Kangas; Diptera: Agromyzidae) of birch trees (Betula pendula and Betula pubescens) provided long time series data (47 and 65 years) of absolute abundance (larvae/tree) in replicated trees within replicated stands. Measurements of tree annual rings provided matching time series of host age and physiological status. Analyses showed a powerful exogenous effect of stand age on temporal variation in insect abundance (58 and 32% of the variance in two populations, respectively). With the additional effects of variation among trees, 77 and 64% of the total variance in abundance was attributable to exogenous bottom-up effects of host plants. Potential endogenous effects were evident as immediate linear density dependence, but only accounted for ≈10% of the total variance. Abundance of Phytobia is primarily a function of disturbance history, which produces a mosaic of different aged birch stands that harbor Phytobia populations of different sizes. Density-dependence tends to regulate local populations around levels determined by host suitability.
Animal population dynamics have fundamentally different characteristics depending on the strength and form of exogenous (density-independent) vs. endogenous (density-dependent) forces (1). It is now generally agreed that some degree of negative density dependence is required for population persistence (1–3). But exogenous factors, defined as those that exert their effects independent of population density, can play important roles in determining the level at which regulation occurs (2). Many factors that affect population abundance, e.g., competition, natural enemies, and resources, can act in either a density-dependent or density-independent fashion (1, 2), but the relative contribution of exogenous and endogenous effects remains an open question for nearly all biological populations.
One technique for exploring population dynamics is the analysis of time series data from natural populations (1). However, there are limitations to what can be learned from time series data. Ideally, time series should include >30 time steps, but such data are very rare for natural populations (1), and, where they exist, they usually come from species with atypical population dynamics, such as pest insects (4–6). Birch trees (Betula spp.) provided a unique record of annual abundance history in an otherwise inconspicuous mining insect, Phytobia betulae (Diptera: Agromyzidae). Replicated time series of 47 and 65 years at two sites allowed us to partition, quantify, and characterize the forces that influence its population abundance in time and space.
Another central problem in contemporary population ecology is understanding the relative contributions of bottom-up (resources) and top-down (natural enemies) forces in the population dynamics of organisms in tritrophic systems (7–9). Normally, time series data for herbivorous insects are measures of relative abundance (e.g., based on light traps, pheromone traps, or aerial defoliation photographs) that cannot be easily related to resource quality or resource availability (7, 10, 11). Our data represent the longest time series yet analyzed that is based on measures of absolute abundance per host plant and also includes measures of resource status (tree age and annual growth).
Study System and Data Collection
P. betulae is a common univoltine herbivore of birch in Fennoscandia. Like the vast majority of insect herbivores, it persists through space and time at relatively low densities. Adults emerge in June, and females lay their eggs into growing shoots within the canopies of birch trees of all ages (12). The larvae that hatch from eggs bore toward the base of the stem for several meters (up to 15 m) while feeding on differentiating xylem close to cambium (12, 13). In August, fully developed larvae bore out through the bark at the stem-base or in the roots to pupate and overwinter in the soil. Larval tunnels become filled with brown parenchyma tissue and remain permanently visible in the annual rings, even in fossil trees (13–15).
We selected two even-aged birch stands, Valtatie (47 years) and Peikkola (65 years), that were situated 700 m apart in southeastern Finland (61°48′ N, 29°18′ E). The Valtatie stand was naturally regenerated, and the Peikkola stand was planted. We determined the age of the Valtatie stand by counting the annual rings of 10 sample trees at the ground level and for the Peikkola stand from planting records. Both stands contained a mixture of two birch species, Betula pendula and Betula pubescens.
We felled 25 (15 B. pendula and 10 B. pubescens; mean height 25 m) and 30 (16 B. pendula and 14 B. pubescens; mean height 21 m) trees at Peikkola and Valtatie, respectively. We took stem disks at the height of 1 meter, where each Phytobia larva leaves only a single scar in the growth ring (13). We censused annual Phytobia population sizes on single tree individuals by counting a total of 31,569 larval tunnels in cross-sections with a stereomicroscope. This yielded 55 time series of Phytobia abundance in individual host trees. We measured the annual growth rate of host trees as the width of the annual rings in the sample disks.
Data Analyses
We analyzed the two sets of time series data (25 and 30 trees at Peikkola and Valtatie, respectively) by testing increasingly complex general linear models in a stepwise approach. Raw data were log transformed, loge(N + 1), which converted to units of proportional change in population size and corrected for heteroscedasticity. Analyses included years 9–61 (1937–1989) at Peikkola and years 7–47 (1953–1993) at Valtatie (we excluded some years early and late in stand development when Phytobia abundance was very low). The statistical model incorporated stand age as a continuous variable, then tree species, and trees nested within species as class variables. With each step in model development, we conducted an F-test to evaluate whether or not the new model provided a significant reduction in variance compared with the simpler model (16). F-tests are approximate because annual abundance of Phytobia within a tree may not be independent among years. This did not interfere with the chief purpose of the model, which was to partition the variation in Phytobia abundance that was attributable to different factors (7).
We studied the relationship between tree growth and Phytobia abundance by correlating the 55 time series of Phytobia abundance (untransformed) and annual ring widths in their individual host trees for both populations separately. We calculated Spearman rank correlations both across trees within each year and across years within each tree.
We evaluated endogenous effects on population dynamics by testing for relationships between population growth rate and previous population density. For each site, population growth rates were calculated as rt = ln (Nt/Nt−1), where Nt and Nt−1 = the average abundance of Phytobia at time t and t − 1 (adjusted for the exogenous effects of stand age and tree species). We plotted partial rate correlation functions (PRCF) of rt to identify the time lags at which negative feedback was operating (2). The PRCF at lag 1 (at time t − 1) was calculated as the correlation coefficient between rt and ln(Nt−1), and PRCF for higher lags as the square root of coefficients of partial determination, the sign of which is determined by the slope of corresponding regression (2, 16). The significance of PRCF was tested with Bartlett’s band ±2/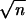 , where n is the length of time series (17). We fitted linear models of population growth rates as a function of lagged population densities Nt−1 and Nt−2. We also tested for more complex, nonlinear endogenous dynamics with a response surface algorithm that searched among alternative first and second order models with and without interactions (18–20) (full model: rt = a0 + a1X + a2Y + a3X2 + a4Y2 + a5XY + ɛt, where X = Nt−1, Y = Nt−2, and X, Y, X2, Y2, and XY could each be transformed by Box-Cox transformations with any of the exponents −1, −0.5, 0, 0.5, or 1) (19, 21). This set of models can describe a broad range of linear and nonlinear response surfaces in population dynamics. The dynamic behavior of the endogenous component is characterized by the dominant Lyapunov exponent. Negative Lyapunov exponents indicate stable dynamics (18, 19).
, where n is the length of time series (17). We fitted linear models of population growth rates as a function of lagged population densities Nt−1 and Nt−2. We also tested for more complex, nonlinear endogenous dynamics with a response surface algorithm that searched among alternative first and second order models with and without interactions (18–20) (full model: rt = a0 + a1X + a2Y + a3X2 + a4Y2 + a5XY + ɛt, where X = Nt−1, Y = Nt−2, and X, Y, X2, Y2, and XY could each be transformed by Box-Cox transformations with any of the exponents −1, −0.5, 0, 0.5, or 1) (19, 21). This set of models can describe a broad range of linear and nonlinear response surfaces in population dynamics. The dynamic behavior of the endogenous component is characterized by the dominant Lyapunov exponent. Negative Lyapunov exponents indicate stable dynamics (18, 19).
We tested for synchronizing effects of climatic variation on Phytobia population dynamics by correlating population growth rates at the two sites for the 37 years of overlap in the data set (1953–1989) (22, 23). Effects of stand age and endogenous dynamics were first removed by using residuals from the models that incorporated both exogenous and endogenous variation.
Results
Both Phytobia populations showed a very similar temporal pattern that followed the aging of host trees (Fig. 1 and 2). The effects of stand age in the Valtatie population were adequately described with a second-order polynomial whereas the Peikkola population, with its longer time series, required a third-order polynomial to describe the slow, gradual decrease in population densities as stand age exceeded 50 years (Table 1, Fig. 2). In the two populations, stand age accounted for 58 and 32% of the total variation in Phytobia abundance (Table 1). Host plant characteristics (age, tree species, host individual) collectively explained 77 and 64% of the variation in Phytobia abundance (Table 1). Tree species explained only a fraction (<2%) of the variation in Phytobia abundance. However, tree individuals within species explained 5 and 23% of the variation (Table 1). At Valtatie and Peikkola, numbers of Phytobia varied from 271 to 1,809 and from 144 to 1,008 per tree during the host lifespans of 47 and 65 years, respectively. Resistant and susceptible trees were intermixed within the stands.
Figure 1.

The time series of Phytobia abundance on two host species, B. pendula and B. pubescens, in two host stands: Peikkola population, years 1929–1993, B. pendula 15 trees, B. pubescens 10 trees; Valtatie population, years 1947–1993, B. pendula 16 trees, B. pubescens 14 trees. The points indicate the annual average numbers of Phytobia per tree.
Figure 2.

The log-transformed time series. The points indicate annual mean abundance in each host species, and the lines show predicted functions (solid line for B. pendula and broken line for B. pubescens). General linear models are described in Table 1.
Table 1.
Analysis of exogenous and endogenous components in Phytobia population dynamics
| Component | Dependent variable | Host stand | Step | Independent variables added | Fmodel | dfmodel | dferror | SS | MSE | r2 | Fimprovement |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Exogenous | Abundance | Peikkola | 1 | StndAge, StndAge2, StndAge3 | 610.93*** | 3 | 1321 | 1302.45 | 0.711 | 0.58 | 610.93*** |
| 2 | Species | 495.59*** | 4 | 1320 | 1345.36 | 0.679 | 0.60 | 63.22*** | |||
| 3 | Tree (Species) | 90.45*** | 27 | 1297 | 1463.78 | 0.599 | 0.65 | 8.59** | |||
| 4 | Species × StndAge, Species × StndAge2 | 88.45*** | 29 | 1295 | 1489.31 | 0.581 | 0.66 | 21.98*** | |||
| 5 | Tree × StndAge, Tree × StndAge2 | 56.37*** | 75 | 1249 | 1730.08 | 0.409 | 0.77 | 12.79*** | |||
| Abundance | Valtatie | 1 | StndAge, StndAge2 | 287.19*** | 2 | 1227 | 550.84 | 0.959 | 0.32 | 287.19*** | |
| 2 | Species | 192.31*** | 3 | 1226 | 552.82 | 0.958 | 0.32 | 2.06 | |||
| 3 | Tree (Species) | 47.15*** | 31 | 1198 | 949.40 | 0.650 | 0.55 | 21.81*** | |||
| 4 | Species × StndAge, Species × StndAge2 | 45.94*** | 33 | 1196 | 965.68 | 0.637 | 0.56 | 12.77*** | |||
| 5 | Tree × StndAge, Tree × StndAge2 | 22.44*** | 89 | 1140 | 1099.77 | 0.551 | 0.64 | 4.35* | |||
| Endogenous | Growth rate | Peikkola | 6 | Nt−1 | 31.22*** | 1 | 49 | 0.2374 | 0.00760 | 0.39 | 31.22*** |
| 7 | Nt−2 | 19.40*** | 2 | 48 | 0.2727 | 0.00703 | 0.45 | 5.02* | |||
| Growth rate | Valtatie | 6 | Nt−1 | 12.51** | 1 | 37 | 0.0427 | 0.00342 | 0.25 | 12.51** | |
| 7 | Nt−2 | 6.82** | 2 | 36 | 0.0465 | 0.00341 | 0.28 | 1.10 |
Exogenous component: development of general linear model quantifying the exogenous effects of host plant factors on the abundance of Phytobia [population density = ln (Phytobia + 1)]. The explanatory factors are: stand age (StndAge), birch species (Species), and tree individuals within birch species (Tree). With each step in the model development, we conducted an F-test (Fimprovement) to evaluate whether the new model allowed a significant reduction in variance compared to the model that included all other factors (Fmodel = F statistic for full model at each step). Endogenous component: stepwise development of linear response surface models testing for effects of population density in year t − 1 (Nt−1) and t − 2 (Nt−2) on population growth rate [rt = ln (Nt/Nt−1)] calculated from residuals of the general linear model steps (1–5). F-tests are approximate and were used primarily as a guide for model development (7, 24). Significance levels are indicated by asterisks: ∗∗∗, P < 0.001; ∗∗, P < 0.01; and ∗, P < 0.05.
There were positive correlations across trees between the annual ring widths and Phytobia abundance, indicating that, within years, fast growing trees tended to support larger populations of Phytobia (Fig. 3). Similarly, there were strong positive correlations across years between annual diameter growth and Phytobia abundance: 70 and 96% of the correlations at Valtatie and Peikkola populations were positive and significant (compare with 6% expected by chance alone) (25).
Figure 3.

Spearman rank correlations, rS, between annual radial growth of the host tree and Phytobia abundance during each year of stand development. Broken lines indicate positive and negative critical values (α = 0.05). Peikkola, B. pendula n = 8–14; Peikkola, B. pubescens n = 5–10; Valtatie, B. pendula n = 5–16; and Valtatie, B. pubescens n = 12–14. Compare 68 significant correlations to 10 that would be expected by chance (205 tests × 0.05) (25). Overall, 94% of the correlations were positive (significantly more positive than negative correlations: Peikkola population χ2 = 41.4, df = 1, P < 0.0001; and Valtatie χ2 = 38.7, df = 1, P < 0.0001). We did not test correlations for a few years when all trees did not include any Phytobia.
Time series analyses revealed endogenous density dependence in Phytobia populations. At both Peikkola and Valtatie, there was a significant negative relationship between population size in the previous year and population growth rate (Table 1, Fig. 4). The PRCF values were significant at both populations: Peikkola PRCF(1) = −0.63 (Bartlett’s band = −0.28) and Valtatie PRCF(1) = −0.54 (Bartlett’s band = −0.32). At Peikkola, there was an additional contribution of population size two years previously (Table 1 and Fig. 4). The PRCF(2) were −0.31 and −0.22 in Peikkola and Valtatie, respectively, but only the value for Peikkola was significant. These endogenous dynamics explained 45 and 25%, respectively, of the interannual variation in detrended population growth rates at Peikkola and Valtatie (or 9–10% of the total variation in abundance) (Table 1). Thus, the endogenous dynamics of Phytobia, detrended for the effects of host attributes, are characterized by stabilizing, linear density dependence. None of the alternative, more complex endogenous models provided a better fit. In simulations, the addition of an effect from Nt−2 at Peikkola (Table 1 and Fig. 4) caused some initial overshoot then undershoot of population size relative to eventual equilibrium, but, regardless of the initial population size, deterministic projections of population size converged to within 1% of the equilibrium within 10 generations. The Lyapunov exponents, −0.816 for the Peikkola model and −1.139 for the Valtatie model, similarly indicated rapid convergence of population size toward a stable equilibrium (18–20).
Figure 4.

Density dependence in Phytobia populations. Phase plots of per capita rate of increase, rt = ln (Nt/Nt−1), of Phytobia populations at Peikkola and Valtatie plotted against lagged population densities (Nt−1 and Nt−2) after detrending the time series with the general linear model described in Table 1 and Fig. 2. The best linear response surface models (Table 1) were a second-order time series model (rt = 1.076–0.198 Nt−1 − 0.086 Nt−2, r2 = 0.45) for Peikkola population, and a first-order time series model (rt = 0.576–0.069 Nt−1, r2 = 0.25) for Valtatie population.
There was no evidence for synchronizing effect of climatic variation on Phytobia abundance; i.e., there was no positive cross-correlation between the two sites (r = −0.15, P = 0.38 adjusted for exogenous effects in Table 1; or r = −0.25, P = 0.13 after further adjusting for endogenous effects in Fig. 4).
Discussion
Our results showed an overwhelming role of host plants in the long-term population dynamics of Phytobia. These exogenous bottom-up effects explained 77 and 64% of the overall variation in Phytobia abundance in Peikkola and Valtatie populations, respectively. Endogenous effects explained only 10 and 9% of the overall variation in both populations. Only 13 and 27% of the variation in Phytobia abundances remained unexplained in Peikkola and Valtatie populations, respectively.
Apparently, Phytobia abundance is positively associated in space and time with the growth and development of host trees. As trees age, they grow too tall for Phytobia larvae to tunnel to the roots before autumn, causing larval mortality (13). The positive correlations between Phytobia abundance and annual radial growth showed that those years when radial growth is greater yield larger populations of Phytobia. Furthermore, Phytobia abundance each year is greater in the individual trees that have been growing most rapidly.
Annual radial growth may have a direct positive effect on the larvae by providing more food and/or space within the differentiating xylem. In addition, annual growth correlates with other factors in the host plant such as the length of new long shoots and the proportion of long shoots in the canopy (26, 27). Phytobia females prefer to lay eggs on the longest of long shoots (T.Y., S. Hinkkanen, H.R., and M.R., unpublished result). As trees age, these shoots become rarer and are present only in the top most part of the canopy (28). Therefore, the availability of suitable oviposition sites may partly determine the maximum density of eggs and larvae in individual trees (29).
Apparently, Phytobia abundance across the landscape of boreal forests is largely driven by disturbance history, which determines the location, extent, and age of birch forests. Presently, clear-cutting silviculture is the dominant disturbance type within Scandinavian forests, but, historically, forest fires were important (10, 30). Such disturbances result in the establishment of a new generation of even-aged birch trees, which creates a habitat patch that will support a population of Phytobia that increases and then decreases in a very predictable pattern over the next 50–60 years. In the absence of additional disturbance, local Phytobia populations eventually go extinct after the senescence of birch trees and their replacement by other tree species.
Phytobia populations separated by <1 km have dynamics that are quite independent. The lack of any detectable synchrony between nearby populations during 37 years of overlap argues against the importance of climatic variation (e.g., temperature and snow cover) or any other spatially autocorrelated exogenous factors (e.g., abundance of natural enemies) for Phytobia population dynamics. This reinforces the importance of the host plant in the long-term population dynamics of Phytobia.
Density dependence tends to regulate local Phytobia populations around an equilibrium that varies with host-suitability. Although time series analyses by themselves cannot identify the specific processes that affect population abundance (1, 7, 31–33), they do provide clues that can be evaluated with other information and subjected to experimental tests. In general, intraspecific competition is a leading candidate to produce rapid density dependence [with a lag of one year in our analyses (1, 2)], but this seems unlikely in the case of Phytobia. The differentiating xylem on which Phytobia feed is in great supply and is unlikely to be a limiting resource. Phytobia adults lay only one egg per shoot and use only a small fraction of the available shoots (T.Y., S. Hinkkanen, H.R., and M.R., unpublished result). Territorial interactions among adults could still produce density dependence (34–36), but we know of no evidence for territoriality in Phytobia. General natural enemies with functional responses or rapid numeric responses are another possibility. However, Phytobia larvae and eggs are largely protected from natural enemies within birch tree, and the predation of adults and pupae is not known. Rapid, induced responses of the trees could be a plausible mechanism for producing density dependence because the mortality of feeding larvae within the stems can be remarkably high (13). If birch responded to larvae with some chemical or developmental mechanism, then response to one larva could negatively impact neighboring larvae. This could produce the rapid density dependence evident in both populations of Phytobia.
If Phytobia abundance in one year impacts tree suitability in the next year, it could produce the delayed density dependence that was evident in the Peikkola population (2). Hymenoptera parasitoids are the only known specialized natural enemies that could explain the delayed density dependence of Phytobia, but the primary infection rates for Phytobia are not known (37–41). The delayed effect in Peikkola population also could be an artifact because the rate of detecting statistical density dependence increases as the length of time series increases (42). Furthermore, an exogenous weather variable could influence Phytobia either directly or indirectly through the effects of the growth of the host plant, in which case it would cause a delayed effect (31–33). Identification of the process that produces density dependence in Phytobia will be a priority in our future work because this is apparently the factor that maintains Phytobia populations way below the numbers that could otherwise be supported by the food resource.
Understanding the dynamics of natural populations is improving as longer time series data become available and methods for extracting endogenous dynamics from noisy time series are developed (3, 43). Exogenous effects on population dynamics are generally difficult to identify because the relevant variables are usually not measured (7, 10, 11). Perhaps because of this, prevailing models tend to treat exogenous effects as “stochastic variation” and instead emphasize the role of endogenous factors. We detected endogenous effects in Phytobia population that might otherwise have been considered unregulated (3). However, the endogenous dynamics by themselves offer a rather unsatisfying description of Phytobia population dynamics. Abundance of Phytobia is primarily a function of forest disturbance history, which produces a mosaic of different aged birch stands that harbor Phytobia populations of different sizes. Our results show that exogenous influences on insect population dynamics can be both strong and highly predictable.
Acknowledgments
We thank A. A. Berryman, I. Hanski, W. J. Mattson, E. Ranta, J. Tahvanainen, and the students of Advanced Population Ecology at Dartmouth College for valuable comments and discussion on the earlier versions of the manuscript. We are grateful to all those involved with data collection, particularly Marko Asikainen, Haijia Li, Kalervo Tervonen, and Olli-Pekka Tikkanen. This work was supported by the Ministry of Agriculture and Forestry, The Foundation for Research of Natural Resources in Finland, Graduate School of Forest Ecology at Faculty of Forestry, University of Joensuu, and Academy of Finland.
ABBREVIATION
- PRCF
partial rate correlation function
References
- 1.Royama T. Analytical Population Dynamics. London: Chapman & Hall; 1992. [Google Scholar]
- 2.Berryman A A, Stenseth N C, Isaev A S. Oecologia. 1987;71:174–184. doi: 10.1007/BF00377282. [DOI] [PubMed] [Google Scholar]
- 3.Turchin P. In: Population Dynamics: New Approaches and Synthesis. Cappuccino N, Price P W, editors. San Diego: Academic; 1995. pp. 17–40. [Google Scholar]
- 4.Turchin P. Nature (London) 1990;344:660–663. [Google Scholar]
- 5.Woiwod I P, Hanski I. J Anim Ecol. 1992;61:619–629. [Google Scholar]
- 6.Hanski I, Woiwod I. Nature (London) 1991;350:28. [Google Scholar]
- 7.Hunter M D, Varley G C, Gradwell G R. Proc Natl Acad Sci USA. 1997;94:9176–9181. doi: 10.1073/pnas.94.17.9176. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Reeve J R, Ayres M P, Lorio P., Jr . In: Population Dynamics: New Approaches and Synthesis. Cappuccino N, Price P W, editors. San Diego: Academic; 1995. pp. 339–357. [Google Scholar]
- 9.Hunter M D, Price P W. In: Effects of Resource Distribution on Animal–Plant Interactions. Hunter M D, Ohgushi T, Price P W, editors. San Diego: Academic; 1992. pp. 1–12. [Google Scholar]
- 10.Dempster P, Pollard E. Oecologia. 1981;50:412–416. doi: 10.1007/BF00344984. [DOI] [PubMed] [Google Scholar]
- 11.Price P W. In: Population Dynamics of Forest Insects. Watt A D, Leather S R, Hunter M D, Kidd N A C, editors. Andover, U.K.: Intercept; 1990. pp. 221–232. [Google Scholar]
- 12.Kangas E. Commun Inst For Fenn. 1935;22:1–31. [Google Scholar]
- 13.Ylioja T, Saranpää P, Roininen H, Rousi M. J Econ Entomol. 1998;91:175–181. [Google Scholar]
- 14.Süss H. Feddes Repertorium. 1979;90:165–172. [Google Scholar]
- 15.Süss H, Müller-Stoll W R. Wiss Z Humboldt-Univ Berlin Math-Naturwiss Reihe. 1975;24:515–519. [Google Scholar]
- 16.Neter J, Wasserman W, Nachtsheim C J, Kutner M H. Applied Linear Statistical Models. Homewood, IL: Irwin; 1996. [Google Scholar]
- 17.Box G E P, Jenkins G M. Time Series Analysis: Forecasting and Control. Oakland, CA: Holden Day; 1976. [Google Scholar]
- 18.Turchin P, Taylor A D. Ecology. 1992;73:289–305. [Google Scholar]
- 19.Turchin P, Millstein J A. EcoDyn/RSM: Response Surface Modeling of Nonlinear Ecological Dynamics. Setauket, NY: Applied Biomathematics; 1993. [Google Scholar]
- 20.Turchin P. Oikos. 1993;68:167–172. [Google Scholar]
- 21.Box G E P, Cox D R. J R Stat Soc B. 1964;26:211–252. [Google Scholar]
- 22.Moran P A P. Aust J Zool. 1953;1:291–298. [Google Scholar]
- 23.Williams D W, Liebhold A M. Environ Entomol. 1995;24:987–995. [Google Scholar]
- 24.Hunter M D. Ecol Entomol. 1998;23:168–173. [Google Scholar]
- 25.Turchin P, Lorio P L, Jr, Taylor A D, Billings R F. Environ Entomol. 1991;20:401–409. [Google Scholar]
- 26.Ilvessalo Y, Ilvessalo M. Acta For Fenn. 1975;144:1–97. [Google Scholar]
- 27.Kramer P J, Kozlowski T T. Physiology of Woody Plants. New York: Academic; 1979. [Google Scholar]
- 28.Kozlowski T T. Growth and Development of Trees. Volume I. Seed Germination, Ontogeny, and Shoot Growth. New York: Academic; 1971. [Google Scholar]
- 29.Mopper S, Simberloff D. Ecology. 1995;76:1233–1241. [Google Scholar]
- 30.Zackrisson O. Oikos. 1977;29:22–32. [Google Scholar]
- 31.Williams D W, Liebhold A M. Ecology. 1995;76:1005–1008. [Google Scholar]
- 32.Williams D W, Liebhold A M. Ecology. 1997;78:320–322. [Google Scholar]
- 33.Hunter M D, Price P W. Ecol Entomol. 1998;23:216–222. [Google Scholar]
- 34.Whitham T G. Ecology. 1978;59:1164–1176. [Google Scholar]
- 35.Whitham T G. Nature (London) 1979;279:324–325. [Google Scholar]
- 36.Whitham T G. Am Nat. 1980;115:449–466. [Google Scholar]
- 37.Martinez M, Gumez J-L, Munnier P. Phytoma. 1985;374:51–53. [Google Scholar]
- 38.Barnes H F. Ann Appl Biol. 1933;20:498–519. [Google Scholar]
- 39.Greene C T. J Agric Res. 1914;1:471–474. [Google Scholar]
- 40.Moraal L G. Entomol Berichten Amst. 1987;47:5–8. [Google Scholar]
- 41.Hanson J B, Benjamin D M. J Econ Entomol. 1967;60:1351–1355. [Google Scholar]
- 42.Godfray H C J, Hassell M P. Nature (London) 1992;359:673–674. [Google Scholar]
- 43.Ellner S, Turchin P. Am Nat. 1995;145:343–375. [Google Scholar]