

Abstract
We have investigated the role of the ryanodine receptor in Drosophila development by using pharmacological and genetic approaches. We identified a P element insertion in the Drosophila ryanodine receptor gene, Ryanodine receptor 44F (Ryr), and used it to generate the hypomorphic allele Ryr16. An examination of hypodermal, visceral, and circulatory muscle showed that, in each case, muscle contraction was impaired in Ryr16 larvae. Treatment with the drug ryanodine, a highly specific modulator of ryanodine receptor channel activity, also inhibited muscle function, and, at high levels, completely blocked hypodermal muscle contraction. These results suggest that the ryanodine receptor is required for proper muscle function and may be essential for excitation-contraction coupling in larval body wall muscles. Nonmuscle roles of Ryr were also investigated. Ryanodine-sensitive Ca2+ stores had previously been implicated in phototransduction; to address this, we generated Ryr16 mutant clones in the adult eye and performed whole-cell, patch–clamp recordings on dissociated ommatidia. Our results do not support a role for Ryr in normal light responses.
The release of Ca2+ from intracellular stores is essential for the initiation and propagation of many Ca2+ signaling events (1). One of the channels that mediates this release from the endoplasmic reticulum is the ryanodine receptor (RyR), which, in vertebrates, is encoded by a family of three genes. The RyR channel is a ≈2-MDa homotetramer, and its activity is regulated by a diverse group of molecules and proteins, including Ca2+, cyclic ADP ribose, Ca2+-calmodulin, and FK506BP (2). All three RyR genes are expressed in brain, and one or more isoforms have been detected in a wide range of tissues (3), but mutations in these genes have largely reflected their roles in muscle contraction. RyR1 is essential for excitation-contraction coupling (ECC) in skeletal muscle (4), whereas RyR2 is important for cardiac muscle contraction (5). In contrast, RyR3-deficient mice showed few overt defects (6), although recent studies suggest a role for RyR3 in spatial learning and synaptic plasticity (7). Functional redundancies among the family members are possible, given their overlapping expression patterns.
The role of RyR in contraction appears to be conserved in invertebrates. The single Caenorhabditis elegans RyR gene is expressed and functions solely in muscles. The null mutant, unc-68, has diminished muscle function, resulting in decreased movement, slow growth, and low brood sizes (8). A single RyR gene, Ryanodine receptor 44F (Ryr), has been cloned in Drosophila melanogaster (this paper, and refs. 9 and 10), and the predicted protein shows ≈45% identity to mammalian family members. During embryogenesis, Ryr was shown to be expressed in developing body wall and head muscles (10), suggesting a role for Ryr in muscle contraction. In adults, Ryr transcript was found in muscles and the brain, and recently it was proposed that ryanodine-sensitive stores are essential for phototransduction (11).
Experimental Procedures
Isolation of Ryr16.
The inverse PCR method of Jay Rehm [Berkeley Drosophila Genome Project (BDGP), unpublished observations] was used to obtain genomic sequence flanking the insertion site of each P element that mapped to 44F (12). The resulting sequences were analyzed by using sequencher software. To generate deletions in Ryr, l(2)k04913, a P{lacW} that carries the miniwhite marker, was mobilized by introducing Ths.PRΔ2–3, a stable source of transposase on the third chromosome. l(2)k04913/CyO; Ths.PRΔ2–3 males were crossed to CyO/Adv females, and 150 white CyO males were selected from the progeny and used to establish individual lines. Genomic DNA from each lethal excision line was screened by PCR using primers upstream of the insertion site and within Ryr. Roughly 10% of the lines yielded PCR products shorter than wild type, and these products were sequenced to determine the deletion boundaries.
Molecular Biology.
Exons encoding the 5′ untranslated region (UTR) of Ryr mRNA were identified by screening the HR library (Berkeley Drosophila Genome Project, unpublished observations) and by 5′ rapid amplification of cDNA ends, which was performed by using the CLONTECH Marathon cDNA Amplification Kit. Exon designation and position within Ryr are as described (ref. 9; Ryr accession no. D17389). Two exons encoded the 154-bp 5′ UTR: P1 and a new exon, designated P0, which was mapped relative to P1 by using Roche Molecular Biochemicals' Long Template PCR. The 5′ boundary of exon P1 was reassigned to nucleotide 231, with the coding sequence starting at position 344.
RNA in Situ.
In situ hybridization was performed on embryos as described (13) with modifications. Probes corresponding to exons within 18,216–25,477 bp and to 4,672–6,400 bp of the Ryr gene were derived from a partial cDNA or made by PCR amplification of genomic sequence, respectively. Digoxygenin (DIG) labeling (and subsequent detection) was carried out by using Roche Molecular Chemicals reagents and protocols with 50 ng of template. DIG probes were purified by ethanol precipitation or with Qiagen (Chatsworth, CA) nucleotide removal kit. To detect probe, embryos were incubated with preadsorbed anti-DIG-AP Fab fragments.
Larval Collection, Visualization, and Ryanodine Treatment.
Assays were performed at room temperature (22°) on 0–2 h, i.e. newly hatched, larvae. For all experiments, w1118 larvae are referred to as “wild type.” Adults laid embryos for 3 h at 25° on molasses agar plates seeded with a dab of yeast paste. To collect larvae, plates were cleared at 20–21 h and incubated for an additional 2 h in the absence of yeast. Larvae were placed on apple juice agar plates to detect the marked balancer CyO-GFParm, in which green fluorescent protein (GFP) is expressed ubiquitously in embryos and larvae under the armadillo promoter (K.M.C.S. and M. Brodsky, unpublished results). GFP was detected with a Leica MZ8 stereomicroscope equipped with a fluorescence module and GFP-Plus filter set. The fluorescent images in Figs. 1 and 4 A and B were taken on a Leica confocal microscope, using an Argon 488-nm laser, GFP filter set, and the pseudocolor overglow, which gave the best contrast. Ethanol stocks of 1–10 mM ryanodine were diluted 1:100 in 5% yeast paste, which was then aliquotted onto glass microfibre filters (GF/C 2.4 cm, Whatman) placed on molasses agar plates. Larvae were allowed to ingest ryanodine for 30 min before performing muscle-contraction assays. Note that the effective concentration of ryanodine in larval muscles is unknown.
Figure 1.

Ryr16 is a deletion in Ryanodine receptor 44F. (A) The first three exons of Ryr (of 27) are represented as rectangles, the filled portions indicating coding and the open portions 5′untranslated region sequences. The inverted triangle marks the position of l(2)k04913, and the region deleted in Ryr16 is indicated by an open rectangle. (H, HindIII; X, XhoI). (B and C) The muscles of newly hatched Ryr16 and Ryr16/CyO larvae are visualized by expressing GFP under control of the Mef2 promoter with the UAS/GAL4 system. GFP fluorescence is shown in pseudocolor. (Scale bars, 10 μm.) (B) Ryr16 heterozygotes phenotypically resemble wild type (data not shown). (C) The muscles of Ryr16 larvae appear to have developed normally, but the mutant animals appear smaller and rounder than wild type, and the head is not properly extended.
Figure 4.

Dorsal vessel contraction is reduced in Ryr16 and ryanodine-treated larvae. (A) GFP is expressed in all muscles of a wild type, second instar larvae (see Fig. 2). (B) The cardial cells of the heart are shown at higher magnification with the dorsal vessel in the closed position. (Scale bars in A and B, 10 μm.) (C) Heart rate was measured by counting the number of beats per minute for a population of each larval genotype. There was no significant difference between the heart rate of wild-type (second instar) and Ryr16/CyO (first instar) larvae. Both Ryr16 larvae and second instar larvae treated with 100 μM ryanodine, however, showed a marked reduction in the rate of contraction. Heart rates were measured for each genotype in at least three independent experiments (n ≥ 15), and the error is the SEM.
Muscle-Contraction Assays.
Newly hatched larvae were placed on a molasses agar plate, and for those undergoing continuous, forward movement (80–100% of the sample), the number of body wall contractions (BWC) that occurred in a 30-s period was counted and converted to BWC/min. For ingestion assays, newly hatched larvae were placed directly on yeast paste dyed with bromophenol blue on a molasses agar plate. The dye is easily visible in the gut: the first section of the midgut appears as a slightly curving posterior extension from the pharynx, the hindgut as an elongated question mark or hook ending at the posterior. At each time point, larvae were removed from food and scored for dye uptake: those positive for ingestion had dark blue staining in the first section of the midgut, those negative had no or very light blue staining. To assay excretion, larvae were fed blue yeast for 4 h, then those with blue dye in the hindgut were transferred to undyed yeast. Larvae scored positive for excretion if the dye had been completely eliminated from their systems. To assay defecation specifically, larvae fed blue yeast as above were placed on a thin layer of undyed yeast for 30 min, and the number of blue spots, representing individual defecation events, was determined. Dorsal vessel (DV) contraction was detected by crossing GAL4mef2.PR (14) or Ryr16/CyO; GAL4mef2.PR females and GFPUAS or Ryr16/CyO; GFPUAS males. Newly hatched progeny were fed blue yeast for 4 h, to eliminate the yolk autofluorescence, enhance contrast between the DV and the underlying gut, and detect mutants. The Ryr16 larvae were identified by their rounded appearance, slow movement, and blue yeast in the pharynx. Contractions were counted by observing the dorsal vessel through the cuticle after rinsing the larvae in water to remove yeast paste.
Electrophysiological Recordings.
Large (>90% of the eye) Ryr16 clones were generated in eyFlp1.2; P{ry+ hs.neo FRT}42D, P{w+}, M53/P{ry+ hs.neo FRT}42D, Ryr16 animals (15). M53 is a minute mutation, and eyFlp1.2 is the flp recombinase expressed under control of the eyeless enhancer (M. Brodsky and G.M.R., unpublished results). Retinas were removed from newly eclosed animals, the white mutant tissue was dissected out and dissociated, and then whole-cell patch–clamp recordings were performed as described (16). The bath solution contained 124 mM NaCl, 4 mM KCl, 10 mM Hepes, 5 mM proline, 25 mM sucrose, 1.5 mM CaCl2, pH 7.15. The pipette solution contained 95 mM potassium gluconate, 40 mM KCl, 10 mM Hepes, 2 mM MgCl2, 0.2 mM EGTA, pH 7.15. Photoreceptors were clamped at a holding potential of −70 mV. Whole-cell capacitances were >35 pF, and seal resistances were >1 GW. Junction potentials were nulled just before seal formation and most (80%) series resistance errors were compensated during recording. In all experiments, light was filtered through a bandpass filter (l = 580 ± 10 nm), neutral density filters, and focused onto the photoreceptor cells by a 0.5 numerical aperture, 40× objective. Stimulation was by means of a 75-W Xenon source; unattenuated output at the stage was 10 mW for white light. Alternatively, heads of Ryr16 mosaic animals were cut coronally, and photoreceptor cells were penetrated with sharp microelectrodes (70–150 MW) filled with 2 M KCl. Bath solution and light stimulation were exactly as described above, except a 530-nm longpass filter was used. Responses obtained from five Ryr16 patches and five wild-type patches were not notably different, in agreement with the patch–clamp results.
Results and Discussion
The Drosophila genome contains a single RyR gene at cytological position 44F (ref. 9, and Berkeley Drosophila Genome Project, unpublished observations). Although an RyR cDNA had previously been partially sequenced and mapped to 76C (10), this localization could not be confirmed (9), and the cDNA's sequence was >99% identical to the corresponding exons at 44F. In this paper, we have subsumed both sets of published results under a single gene, renamed the Ryanodine receptor 44F (Ryr).
We obtained genomic sequence flanking each P element in the 44F region and determined that P{lacW}l(2)k04913 was inserted in an intron of Ryr, 255 bp upstream of the first coding exon (Fig. 1A). We excised l(2)k04913 and found that 80% of the independent excision lines were viable, indicating that the lethal lesion of l(2)k04913 corresponded to the P element insertion site and that there were no other lethal mutations on the chromosome. All but one of the lethal excisions failed to complement l(2)k04913 and Df(2R)Np3, a deficiency that uncovers Ryrk04913, confirming that these mutations were allelic to the P element. Using PCR, we identified four lethal excision lines that had deletions specifically in Ryr. The largest, Ryr16, removed the first coding exon and extended into the first and second introns (Fig. 1A).
To identify mutants, the Ryr16 chromosome was placed over the balancer CyO-GFParm, which specifically marked heterozygous animals with GFP. The development of Ryr16 embryos is normal as determined by direct observation and with myosin heavy chain (muscle) and fasciclin II antibodies, which stain muscles and a subset of embryonic central nervous system axons (data not shown). However, in contrast to Ryr16/CyO-GFParm larvae, which were indistinguishable from wild type, Ryr16 larvae appeared short and rounded (Fig. 1 B and C); furthermore, they did not grow and died over 4–7 days as first instar larvae. To control for developmental changes and for defects caused indirectly by malnutrition, all experiments were performed on newly hatched larvae, unless otherwise stated. It should be noted that Ryr16 larvae appeared unchanged for the first 12 h after hatching, as assayed by BWC, and not until 12–24 h was an increase in phenotypic severity observed (data not shown).
Lethal stage analysis revealed that approximately 60% of Ryr16/Df(2)Np3 embryos failed to hatch, whereas newly hatched Ryrk04913 larvae appeared normal and did not cease growth and die until the late first/second instar stages (data not shown). The relative allelic strength was tentatively assigned as Ryr16/Df(2R)Np3 > Ryr16 > Ryrk04913. These results indicate that Ryr16 is not a null allele, despite deletion of the first coding exon. Reverse transcription–PCR experiments confirmed the presence of Ryr mRNA in Ryr16 larvae (data not shown); thus, a functional, truncated ryanodine receptor may be synthesized. Ryr encodes a protein >5,000 amino acids in length that contains many methionine residues. The transmembrane, channel-forming domain is found at the extreme C terminus, and it has been shown for RyR1 that the C-terminal third of the protein is sufficient to make a functional channel (17).
Although Ryr transcript was reported to be exclusively localized to hypodermal and head muscles during late embryogenesis (10), we found Ryr to be more widely expressed. RNA in situs were performed on embryos using both genomic- and cDNA-derived probes. Ryr expression was first detected in the mesoderm around stage 9 and then increased starting at stage 13 (Fig. 2 A and B). The highest levels were seen in hypodermal muscles and in the visceral muscles surrounding the gut. The transcript was also detected at lower levels in other tissues, notably the central nervous system.
Figure 2.
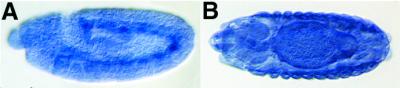
Ryr mRNA is detected in developing visceral and hypodermal muscles. In situ hybridization was performed on wild-type embryos with DIG-labeled probes made from a partial Ryr cDNA. (A) A stage-11 embryo is shown in a lateral view, with dorsal up and anterior to the left. Expression of Ryr mRNA is strongest in the mesoderm. (B) A stage-14 embryo is viewed from the dorsal perspective, anterior to the left. Ryr expression appears widespread with high levels in hypodermal and visceral muscles.
Based on the in situ results, we first examined the role of Ryr in hypodermal or body wall muscles, which are analogous in structure and function to vertebrate skeletal muscle (18). Forward movement is initiated by contraction of the posterior body wall muscles, moving the tail up, forward, and down. The contraction propagates as a constriction anteriorly, narrowing and lengthening the larval body, and finally terminates on extension of the mouth hooks up and forward. Initiation of a new BWC typically does not occur until termination of the previous one; however, initiation and propagation appear to be independent processes. For example, in calmodulin null mutants, initiation of BWCs was greatly decreased, whereas propagation occurred at normal speeds once initiated (19).
BWC was quantified by counting the number of end-to-end contractions a single larva underwent per minute. In Ryr16 larvae, the timing of BWC initiation appeared normal, but the contractions propagated more slowly than in wild-type and Ryr16/CyO-GFParm controls, such that the rate of BWC was reduced by 50% (Fig. 3A). Furthermore, contraction in Ryr16 animals was attenuated and often had associated tremors; this muscle weakness is probably responsible for the altered larval appearance. As expected, Ryr16/Df(2R)Np3 larvae had a more severe defect in BWC propagation, which was reduced by over 90% (Fig. 3A). The weakest allele, Ryrk04913, had a small but significant decrease in the rate of BWC propagation compared with wild-type and Ryrk04913/CyO-GFParm controls. By the second instar, Ryrk04913 larvae had visible contraction defects in hypodermal muscle, which included tilting and dragging of the mouth hooks (data not shown).
Figure 3.

Muscle function is impaired in ryanodine-treated and Ryr16 larvae. (A) The average BWC/min was determined for newly hatched larvae of the specified genotype. The wild type, Itp-r1, Ryrk04913/CyO-GFParm, and Ryr16/CyO-GFParm larvae performed identically in this assay. Ryrk04913 larvae had a slight, reproducible reduction, Ryr16 a ≈50% reduction, and Ryr16/Df(2R)Np3 hemizygotes a 90% decrease in BWC/min. The BWC/min for each genotype was measured at least three times on separate collections of larvae (n = 30), and the error is the SEM. (B) Yeast paste freshly doped with 0.01–100 μM ryanodine was fed to newly hatched Ryr16/CyO-GFParm, and the effect on BWC rates was determined after 30 min. Concentrations of ryanodine 1 μM or less and the ethanol control (not shown) had no significant effect on BWC rates, and concentrations ≥100 μM completely inhibited BWC. For each concentration of ryanodine, the BWC measurement was performed on at least two separate larval samples (n = 30), and the error is the SEM. (C) Larvae scored positive for ingestion if they had a concentrated level of dye in the first section of the midgut. Representative curves are shown for Ryr16/CyO-GFParm (○) and Itp-r1 (□). Ryrk04913 larvae (◊) showed only a slight lag in reaching 100% positive compared with controls, but the delay was reproducible in multiple trials. In contrast, the percentage of positive Ryr16 larvae (▵) typically ranged from 40% to 60% at the end of a 6-h time course. (D) To assay excretion, larvae positive for ingestion after feeding blue yeast for 4 h were transferred to unadulterated yeast paste and scored for the complete loss of dye. Ryr16/CyO-GFParm (○), Itp-r1 (□), and wild-type larvae (not shown) had nearly identical excretion time courses. The rate of excretion was significantly decreased in Ryr16 larvae (▵). For C and D, each larval genotype was assayed in at least three independent trials (n ≥ 30).
Recent reports suggest that larval growth defects do not necessarily impair feeding or locomotion (20, 21). To confirm this, BWC was examined in Itp-r1 larvae. Itp-r1 is a deletion in the InsP3 receptor gene, and these mutants do not progress beyond the second instar (22). Despite the larval growth defect, BWC rates in newly hatched Itp-r1 were completely normal (Fig. 3A).
The drug ryanodine is a highly specific modulator of the RyR channel. In vitro studies have demonstrated that low doses of ryanodine (<10 μM) activate the channel, medium doses (>10 μM) lock the channel into a subconductance state, and high concentrations (≥100 μM) completely inactivate it (23, 24). A range of ryanodine concentrations was fed to newly hatched larvae in yeast paste, and the effect on BWC rates was determined (Fig. 3B). Low concentrations of ryanodine (<5 μM) had no significant effect on BWC rates when compared to larvae fed yeast paste doped with solvent alone (data not shown). Higher ryanodine levels (5–100 μM) decreased the rate of BWC propagation, but not initiation, and at the highest doses (≥100 μM), BWC was completely inhibited. Ryanodine similarly inhibited BWC in second and third instar larvae (data not shown). Ryanodine concentrations ≥10 μM also caused larvae to round up, altering their appearance in a manner similar to that of Ryr16. These results demonstrate that Ryr16 is phenocopied by inhibitory concentrations of ryanodine, which is expected if the drug and the mutation both target Ryr. The complete inhibition of BWC by ryanodine provides further evidence that Ryr16 is not a null allele and suggests that the ryanodine receptor is essential for ECC in hypodermal muscles. In contrast, C. elegans can undergo muscle contraction even in the absence of RyR activity (8).
Visceral muscle function was analyzed in wild-type and Ryr16 larvae by assaying the ingestion and excretion of food. The dye bromophenol blue is nontoxic, readily ingested, and completely excreted by larvae, as well as easy to detect through the cuticle. In a typical ingestion assay (Fig. 3C), 96–100% of wild-type (not shown), Itp-r1, and Ryr16/CyO-GFParm larvae scored positive for ingestion within 30 min. A very mild, but reproducible, ingestion defect was seen in Ryrk04913 larvae. In sharp contrast, Ryr16 larvae ingested food much more slowly, and the percentage that scored positive failed to reach 100 even after 2 days (data not shown). Additionally, Ryr16 larvae accumulated food in the pharynx, which was never seen in controls, and those that failed to ingest food were indistinguishable in growth and movement from those that did (data not shown). Ryr16 mutants may have additional defects in nutrient absorption or metabolism; however, it may simply be that none of the Ryr16 larvae ingest sufficient food for growth. Our results demonstrate that Ryr16 mutants have a severe defect in the ingestion and passage of food into the gut, suggesting that the head and visceral muscles are impaired.
Pulse–chase and defecation assays were used to measure visceral muscle function specifically. Newly hatched larvae were fed blue yeast for 4 h, then transferred to undyed yeast and scored for complete loss of the dye. In a typical excretion time course, 100% of Ryr16/CyO-GFParm, Itp-r1, and wild-type larvae (Fig. 3D, and data not shown) had completely excreted the dye within 3 h. In contrast, Ryr16 larvae excreted the dye extremely slowly, such that roughly half still retained dye in the gut when assayed for up to 12 h. As a second test for visceral muscle function, we measured the rate of defecation. Wild-type and heterozygous larvae on average defecated once every 4 min, Ryrk04913 once every 7 min, and Ryr16 less than once per 60 min. Both assays demonstrate that Ryr16 larvae have a severe defect in excretion, consistent with impaired visceral muscle function.
Drosophila visceral muscles, although considered striated muscles, are distinct from those of the hypodermus: they are much smaller (1 μm in diameter) and mononucleate; the contractile elements are reduced in density and more disorganized; and the sarcoplasmic reticulum (SR) is less developed (25, 26). It has been proposed that the extent of SR in a given muscle type correlates with the requirement for intracellular Ca2+, and thus the RyR, in contraction (10). However, this hypothesis is inconsistent with the severe visceral defect seen in Ryr16 larvae. One possibility is that visceral muscle ECC requires Ca2+ release from internal stores, but not the rapid, widespread release enabled by extensively developed stores. Alternatively, intracellular Ca2+ stores may be essential for some other process equally vital for the passage of food through the gut, such as Ca2+ homeostasis or muscle relaxation.
The DV is the major pulsatile organ of the Drosophila circulatory system (27). The contractile region or heart is a tube-shaped chamber of cardial cells in the posterior segments that circulates the hemolymph by lateral constriction (Fig. 4 A and B). The aorta extends anteriorly and connects to the ring gland and lymph nodes. Little is known about the structure or physiology of the circulatory muscle, although L-type channels have been implicated in DV contraction (28), as is the case in vertebrate cardiac cells. DV function and development have been compared with that of the vertebrate heart, although it has recently been argued that visceral muscle development is the more analogous (14).
By expressing GFP in muscles, we were able to examine the larval heartbeat in vivo for all three larval stages and found that it remained relatively consistent (data not shown). The heartbeat frequency ranged from slow to fast to fibrillating, with the heart occasionally pausing for up to several seconds; more rarely, localized contractions or twitches occurred. There was no obvious correlation between larval behavior or movement and heart rate. Ryr16/CyO and wild-type heart rates and behavior were indistinguishable (Fig. 4C). In Ryr16 larvae, the heart rate was reduced by ≈75% relative to the heterozygotes. The decrease in heart rate was caused by loss of the fast and fibrillating contractions but not an increase in pausing.
The effect of ryanodine on the dorsal vessel was examined in wild-type first (data not shown) and second (Fig. 4C) instar larvae. Feeding larvae ≥25 μM ryanodine decreased heart rates (data not shown), and ≥100 μM ryanodine reduced it by ≈85% relative to wild type. It was not possible to completely inhibit circulatory muscle contraction with ryanodine as it was in the case of hypodermal muscles. This may reflect experimental limitations, because the larvae must be washed to observe the GFP signal. As detected by increasing BWC rates, larvae rapidly recovered when washed out of ryanodine. Alternatively, the ryanodine receptor may not be essential for ECC in circulatory muscles.
In previous studies (11), it was observed that depletion of Ca2+ stores in dissected photoreceptor cells by ryanodine inhibited subsequent light responses. This effect could be rescued by the addition of Ca2+-calmodulin, which inhibits Ca2+ release by the ryanodine receptor. Based on these results, it was proposed that Ca2+ release through the RyR is required for phototransduction. However, these experiments could not distinguish a pleiotropic effect of the pharmacological depletion of internal stores from an actual role for the RyR in light transduction. To test this, Ryr16 mutant eye clones were generated in heterozygous adults by using the Flp/FRT system (15). We generated clones representing >90% of the eye using eyFlp and a marker chromosome carrying the minute (M53) mutation, which slows cellular growth. The eyeless enhancer drives Flp expression specifically in the eye disc, and the resulting Ryr16 mutant cells outgrow both M53/Ryr16 and M53/M53 cells in mosaic tissue. Ryr16 clones appeared morphologically normal (data not shown); furthermore, whole-cell, voltage-clamp recordings on Ryr16 ommatidia showed responses indistinguishable from wild type (Fig. 5 A and B). Taken together, these results are inconsistent with the postulate that Ryr is required for phototransduction.
Figure 5.

Phototransduction is normal in Ryr16 eye clones. (A) Representative whole-cell voltage-clamp recordings of light-induced currents from a wild type (wt) and a Ryr16 photoreceptor. (Left) Cells were stimulated with 10-ms flashes of increasing light intensity given at the arrow. Numbers refer to the log order of light intensity associated with that light response (e.g., −4 is 10 times less light than −3). (Right) Cells were stimulated with a 400-ms pulse of light (log [I] = −1). In each case, Ryr16 responses are indistinguishable from wild type. (B) Quantitative analysis of responses to 10-ms flashes of light. The plot shows peak response amplitude vs. light intensity for wild type and Ryr16, and the bar graphs compare rise time (measured as the time between 10–90% rise of the peak current) and decay time (measured as the time between 90–10% decline of the current) vs. light intensity. Black bars correspond to wild type, white to Ryr16. Each data point represents the mean response ± SE for seven wt cells and 15 Ryr16 mutant cells sampled from 12 Ryr16 mutant patches.
To examine the role of Ryr in embryonic development, we generated females with germ-line clones homozygous for Ryr16 using the dominant female sterile technique (29). However, the progeny of these females, when crossed to Ryr16 males, were phenotypically identical to either wild type or Ryr16 (data not shown). Consistent with this, RNA interference (30) using double-stranded RNA transcribed from Ryr cDNA had no apparent effect on embryogenesis, although a majority of the injected embryos that hatched did have muscle contraction defects (data not shown). The overall behavior of Ryr16 larvae was indistinguishable from wild type (data not shown): they detected and migrated to food sources; had an enhanced rate of food ingestion after starvation; had normal salt avoidance and response to mechanical stimulation; and showed no spontaneous avoidance behavior. BrdUrd labeling of Ryr16 larval brains showed that cell cycle progression is normal in these mutants. Thus, at present our results provide no evidence that Ryr has any nonmuscle roles in Drosophila development.
Acknowledgments
We thank Laurie Lee Camp, Robert Williams, Tom Serano, and Amy Beaton for critical comments and helpful advice. The Corey S. Goodman lab generously supplied antibodies, and Eric Olsen and his lab provided GAL4mef2 flies. Tilmann Brotz and Holly Aaron provided help with confocal imaging. We are also indebted to Todd Laverty for cytological mapping, Susan Mullaney for embryo injections, Martha E. Holm for sequencing, and Amy H. Tang for help with larval manipulations. This work was funded by the Howard Hughes Medical Institute (to G.M.R.) and a Damon Runyon-Walter Winchell fellowship (to K.M.C.S.).
Abbreviations
- RyR
ryanodine receptor
- ECC
excitation-contraction coupling
- DIG
digoxygenin
- GFP
green fluorescent protein
- BWC
body cell contraction
- DV
dorsal vessel
Footnotes
Article published online before print: Proc. Natl. Acad. Sci. USA, 10.1073/pnas.110145997.
Article and publication date are at www.pnas.org/cgi/doi/10.1073/pnas.110145997
References
- 1.Berridge M J, Bootman M D, Lipp P. Nature (London) 1998;395:645–648. doi: 10.1038/27094. [DOI] [PubMed] [Google Scholar]
- 2.Sitsapesan R, McGarry S J, Williams A J. Trends Pharmacol Sci. 1995;16:386–391. doi: 10.1016/s0165-6147(00)89080-x. [DOI] [PubMed] [Google Scholar]
- 3.Sutko J L, Airey J A. Physiol Rev. 1996;76:1027–1071. doi: 10.1152/physrev.1996.76.4.1027. [DOI] [PubMed] [Google Scholar]
- 4.Takeshima H, Iino M, Takekura H, Nishi M, Kuno J, Minowa O, Takano H, Noda T. Nature (London) 1994;369:556–559. doi: 10.1038/369556a0. [DOI] [PubMed] [Google Scholar]
- 5.Takeshima H, Komazaki S, Hirose K, Nishi M, Noda T, Iino M. EMBO J. 1998;17:3309–3316. doi: 10.1093/emboj/17.12.3309. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Bertocchini F, Ovitt C E, Conti A, Barone V, Scholer H R, Bottinelli R, Reggiani C, Sorrentino V. EMBO J. 1997;16:6956–6963. doi: 10.1093/emboj/16.23.6956. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Balschun D, Wolfer D P, Bertocchini F, Barone V, Conti A, Zuschratter W, Missiaen L, Lipp H P, Frey J U, Sorrentino V. EMBO J. 1999;18:5264–5273. doi: 10.1093/emboj/18.19.5264. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Maryon E B, Saari B, Anderson P. J Cell Sci. 1998;111:2885–2895. doi: 10.1242/jcs.111.19.2885. [DOI] [PubMed] [Google Scholar]
- 9.Takeshima H, Nishi M, Iwabe N, Miyata T, Hosoya T, Masai I, Hotta Y. FEBS Lett. 1994;337:81–87. doi: 10.1016/0014-5793(94)80634-9. [DOI] [PubMed] [Google Scholar]
- 10.Hasan G, Rosbach M. Development (Cambridge, UK) 1992;116:967–975. doi: 10.1242/dev.116.4.967. [DOI] [PubMed] [Google Scholar]
- 11.Arnon A, Cook B, Montell C, Selinger Z, Minke B. Science. 1997;275:1119–1121. doi: 10.1126/science.275.5303.1119. [DOI] [PubMed] [Google Scholar]
- 12.Spradling A C, Stern D M, Kiss I, Roote J, Laverty T, Rubin G M. Proc Natl Acad Sci USA. 1995;92:10824–10830. doi: 10.1073/pnas.92.24.10824. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Tautz D, Pfeifle C. Chromosoma. 1989;98:81–85. doi: 10.1007/BF00291041. [DOI] [PubMed] [Google Scholar]
- 14.Ranganayakulu G, Elliott D A, Harvey R P, Olson E N. Development (Cambridge, UK) 1998;125:3037–3048. doi: 10.1242/dev.125.16.3037. [DOI] [PubMed] [Google Scholar]
- 15.Xu T, Rubin G M. Development (Cambridge, UK) 1993;117:1223–1237. doi: 10.1242/dev.117.4.1223. [DOI] [PubMed] [Google Scholar]
- 16.Ranganathan R, Harris G L, Stevens C F, Zuker C S. Nature (London) 1991;354:230–232. doi: 10.1038/354230a0. [DOI] [PubMed] [Google Scholar]
- 17.Bhat M B, Zhao J, Takeshima H, Ma J. Biophys J. 1997;73:1329–1336. doi: 10.1016/S0006-3495(97)78166-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Crossley A C. In: The Genetics and Biology of Drosophila. Ashburner M, Wright T R F, editors. London: Academic; 1980. pp. 499–560. [Google Scholar]
- 19.Heiman R G, Atkinson R C, Andruss B F, Bolduc C, Kovalick G E, Beckingham K. Proc Natl Acad Sci USA. 1996;93:2420–2425. doi: 10.1073/pnas.93.6.2420. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Migeon J C, Garfinkel M S, Edgar B A. Mol Biol Cell. 1999;10:1733–1744. doi: 10.1091/mbc.10.6.1733. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Galloni M, Edgar B A. Development (Cambridge, UK) 1999;126:2365–2375. doi: 10.1242/dev.126.11.2365. [DOI] [PubMed] [Google Scholar]
- 22.Acharya J, Jalink K, Hardy R W, Hartenstein V, Zuker C S. Neuron. 1997;18:881–887. doi: 10.1016/s0896-6273(00)80328-1. [DOI] [PubMed] [Google Scholar]
- 23.Lai F A, Misra M, Xu L, Smith A, Meissner G. J Biol Chem. 1989;264:16776–16785. [PubMed] [Google Scholar]
- 24.Meissner G. J Biol Chem. 1986;261:6300–6306. [PubMed] [Google Scholar]
- 25.Goldstein M A, Burdette W J. J Morphol. 1971;134:315–334. doi: 10.1002/jmor.1051340305. [DOI] [PubMed] [Google Scholar]
- 26.Sandborn E B, Duclos S, Messier P-E, Roberge J-J. J Ultrastruct Res. 1967;18:695–702. doi: 10.1016/s0022-5320(67)80214-4. [DOI] [PubMed] [Google Scholar]
- 27.Rizki T M. In: The Genetics and Biology of Drosophila. Ashburner M, Wright T R F, editors. London: Academic; 1980. pp. 397–452. [Google Scholar]
- 28.Gu G-G, Singh S. J Neurobiol. 1995;28:269–280. doi: 10.1002/neu.480280302. [DOI] [PubMed] [Google Scholar]
- 29.Chou T-B, Noll E, Perrimon N. Development (Cambridge, UK) 1993;119:1359–1369. doi: 10.1242/dev.119.4.1359. [DOI] [PubMed] [Google Scholar]
- 30.Misquitta L, Paterson B M. Proc Natl Acad Sci USA. 1999;96:1451–1456. doi: 10.1073/pnas.96.4.1451. [DOI] [PMC free article] [PubMed] [Google Scholar]