

Abstract
Primary causes of heterosis are still unknown. Our goal was to investigate the extent and underlying genetic causes of heterosis for five biomass-related traits in Arabidopsis thaliana. We (i) investigated the relative contribution of dominance and epistatic effects to heterosis in the hybrid C24 × Col-0 by generation means analysis and estimates of variance components based on a triple testcross (TTC) design with recombinant inbred lines (RILs), (ii) estimated the average degree of dominance, and (iii) examined the importance of reciprocal and maternal effects in this cross. In total, 234 RILs were crossed to parental lines and their F1's. Midparent heterosis (MPH) was high for rosette diameter at 22 days after sowing (DAS) and 29 DAS, growth rate (GR), and biomass yield (BY). Using the F2-metric, directional dominance prevailed for the majority of traits studied but reciprocal and maternal effects were not significant. Additive and dominance variances were significant for all traits. Additive × additive and dominance × dominance variances were significant for all traits but GR. We conclude that dominance as well as digenic and possibly higher-order epistatic effects play an important role in heterosis for biomass-related traits. Our results encourage the use of Arabidopsis hybrid C24 × Col-0 for identification and description of quantitative trait loci (QTL) for heterosis for biomass-related traits and further genomic studies.
HYBRID vigor or heterosis is the superior performance of an F1 hybrid compared with the mean of its parental lines. In breeding of allogamous crops, the phenomenon of heterosis has been heavily exploited in all breeding categories except line breeding (Simmonds and Smartt 1999), mainly because of its large effect on grain and biomass yield (Becker 1993). However, the causes underlying this important phenomenon have remained unknown, even though many genetical, physiological, and molecular explanations have been suggested. Genetic hypotheses, such as dominance, overdominance, and epistasis were the first to be put forward and are still prevailing (cf. Lamkey and Edwards 1999).
Since there is strong evidence that heterosis is a polygenic phenomenon, its causes have been investigated primarily by quantitative genetics research, using either first- or second-degree statistics (cf. Hallauer and Miranda 1981). The former approach estimates the net contribution of dominance and epistatic effects of loci affecting heterosis of a trait by generation means or diallel analyses. In maize and other species, most studies demonstrated that dominance and/or overdominance are of major importance in heterosis for grain yield (Melchinger et al. 1986). Epistasis cannot be ruled out entirely, but its net effect on testcross generation means seems to be of secondary importance in most crosses of elite lines (Hinze and Lamkey 2003; Mihaljevic et al. 2005).
Alternatively, second-degree statistics estimate variance components attributable to additive, dominance, and epistatic effects from covariances of relatives. In particular, design III has played a prominent role in estimating the average degree of dominance over loci from the ratio of dominance to additive variance (Comstock and Robinson 1952). However, linkage between quantitative trait loci (QTL) in repulsion phase may mimic pseudooverdominance and, thus, result in biased estimates of the average squared degree of dominance. Moreover, the effects of epistasis are generally ignored in estimates of additive and dominance variance components obtained by design III. The triple testcross (TTC) design, proposed by Kearsey and Jinks (1968) as an extension of design III, provides a test for epistatic effects (A. E. Melchinger, H. F. Utz, H. P. Piepho and C. C. Schön, unpublished results).
Maize has long served as a primary model species for heterosis research due to the huge amount of heterosis displayed for grain yield. However, the completely sequenced genomes and most advanced genetic tools and resources available in Arabidopsis and rice recommend these two species as experimental model organisms in plant genetics and functional genomics. Up to now, several studies are available on heterosis in rice (Xiao et al. 1995; Yu et al. 1997; Li et al. 2001; Luo et al. 2001; Hua et al. 2002, 2003), whereas the information on heterosis in Arabidopsis is scarce. Kearsey et al. (2003) employed the TTC design to study the genetics of 22 quantitative traits related to plant size and development in the Arabidopsis hybrid Columbia (Col) × Landsberg (Ler). However, inferences on heterosis could not be made with the genetic material studied, because significant heterosis was observed only for one of the 22 traits. On the contrary, the extent of midparent heterosis (MPH) and best-parent heterosis (BPH) for biomass yield (MPH = 60.3%, BPH = 32.9%) and rosette diameter (MPH = 49.4%, BPH = 34.8%) averaged across five Arabidopsis hybrids derived from five ecotypes (Barth et al. 2003) was surprisingly high. The values of MPH and BPH of individual hybrids varied substantially, with even 140.1% MPH for biomass yield in one hybrid (C24 × Aa-0). Another systematic survey of 63 Arabidopsis accessions crossed with three reference lines (Meyer et al. 2004) also reported significant MPH for biomass yield, with hybrid Col-0 × C24 displaying significant hybrid vigor.
The goals of our study were to determine the extent of heterosis for biomass-related traits in Arabidopsis hybrid C24 × Col-0 and investigate its underlying genetic causes by quantitative genetic analyses. In particular, our objectives were to (i) investigate the relative contribution of dominance and epistatic effects to heterosis in this cross by generation means analysis and estimates of variance components calculated from a TTC design with recombinant inbred lines (RILs), (ii) estimate the average degree of dominance, and (iii) examine the importance of reciprocal and maternal effects.
MATERIALS AND METHODS
Plant materials:
Seeds from the Arabidopsis thaliana accessions C24 (provided by J. P. Hernalsteens, Vrije Universiteit Brussels, Belgium) and Col-0 (provided by G. Rédei; University of Missouri, Columbia, MO), subsequently referred to as parents P1 and P2, respectively, were used to establish the plant materials employed in this study. The F1 generation was produced in two reciprocal forms: P1 × P2 (F1-a) and P2 × P1 (F1-b), where the first parent refers to the seed parent. The F2 generation originating from F1-a is further referred to as F2-a and that originating from F1-b as F2-b. Two sets of RILs were derived as described by Törjék et al. (2006). RILs-a comprised 214 RILs derived from F1-a, and RILs-b comprised 209 RILs derived from F1-b. For this study, we used random subsets of 111 RILs-a and 123 RILs-b. The lines were propagated via single-seed descent to generation F7. Testcross progenies were produced using the TTC design (Kearsey and Jinks 1968). RILs were used as pollen parents and mated with P1 and P2. In addition, RILs-a were crossed with F1-a and RILs-b with F1-b. In all instances, one representative plant of each RIL was used to pollinate three plants of each tester (P1, P2, F1-a, or F1-b) and, apart from six siliques per mother plant, all others were removed to warrant a homogeneous seed size.
Experimental design:
The entire set of 234 RILs was subdivided into three experiments, each with 78 RILs and six checks. Experiments were arranged in a split-plot design with three replicates. Main plots were arranged in a 12 × 7 α-design. Each main plot comprised four entries: one RIL and its three testcross progenies produced by the TTC design. The main plots of checks also comprised four entries: parents P1 and P2, as well as the F1 and F2 generations either from F1-a or from F1-b. In all instances, the entries within each main plot were randomly assigned to the subplots. Each subplot consisted of a row of 10 plants per entry. For the subsequent statistical analyses with SAS PROC MIXED (SAS Institute 2004) described below, treatment effects were considered as fixed for checks as well as all other generations listed (Table 1) and considered as random for entries nested within generations (RILs and their TTC progenies). Apart from these treatment effects, the model contained fixed effects for experiments, replicates within experiments, and incomplete blocks and contained random effects for main plots and subplots.
TABLE 1.
Genetic expectations for the generation means of generations P1, P2, F1, and F2 in both reciprocal forms (a = P1 × P2; b = P2 × P1), corresponding RILs from F1-a and F1-b, as well as triple testcross progenies with testers P1, P2, and F1
| Genetic parametera
|
|||||||||
|---|---|---|---|---|---|---|---|---|---|
| Generation | Nb | μ | c | m | [a] | [d] | [aa] | [ad + da] | [dd] |
| P1 | 18 | 1 | 1 | 0 | −1 | −
|
1 |  |
 |
| P2 | 18 | 1 | 0 | 0 | 1 | −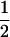
|
1 | −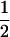
|
 |
| F1-a | 9 | 1 | 1 | 0 | 0 |  |
0 | 0 | 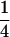 |
| F1-b | 9 | 1 | 0 | 0 | 0 |  |
0 | 0 | 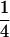 |
| F2-a | 9 | 1 | 1 | 1 | 0 | 0 | 0 | 0 | 0 |
| F2-b | 9 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 0 |
| RILs-a | 123 | 1 | 1 | 0 | 0 | −
|
0 | 0 |  |
| RILs-b | 111 | 1 | 0 | 0 | 0 | −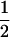
|
0 | 0 |  |
| P1 × RILs | 234 | 1 | 1 | 0 | −
|
0 | 1/4 | 0 | 0 |
| P2 × RILs | 234 | 1 | 0 | 0 | 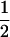 |
0 | 1/4 | 0 | 0 |
| F1-a × RILs-a | 123 | 1 | 1 | 1 | 0 | 0 | 0 | 0 | 0 |
| F1-b × RILs-b | 111 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 0 |
| General coefficient | 1 | u | v | x | z | x2 | xz | z2 | |
For a detailed explanation of the terminology and definition of genetic effects, see materials and methods.
Number of entries.
Plant cultivation:
Seeds were sown in petri dishes under sterile conditions on Murashige–Skoog medium. At the two-leaf stage, ∼10 days after sowing (DAS), each seedling was transferred to sterilized soil (Euflor GmbH: 90% peat, 7% perlite, and 3 vol% sand at pH of 5–6, salt content <1.5 g/liter, nitrogen availability <300 mg/liter N, phosphate availability <300 mg/liter P2O5, and calcium oxide availability <400 mg/liter K2O). All plants in a replicate were grown on a large bench of sterilized soil. Distance between plants inside the row as well as the distance between rows was 8 cm. Plants in soil were irrigated with tap water. Standard light and temperature regime under greenhouse conditions was 16 hr light (20,000 lux) at 21° and 8 hr dark at 18°.
Traits measured:
Rosette diameter (RD) (in millimeters) was recorded on a single-plant basis 22 days after sowing (RD22) and 29 days after sowing (RD29). The absolute growth rate (GR) per day (in millimeters per day) was determined as (RD29 − RD22)/7; this provides an approximation of the expansion of the rosette diameter under an exponential growth model. All plants of a subplot were harvested without the root system at 29 DAS and bulked into a plastic jar. Biomass yield (BY) (in milligrams) was recorded after drying in an oven to practically 0% moisture content. Dry matter content (DMC) (in percent) was calculated as the ratio between dry and fresh biomass, ×100.
Generation means:
Means and standard errors of all generations were calculated over the series of three split-plot experiments with SAS PROC MIXED (SAS Institute 2004). Midparent heterosis was calculated as MPH = 100 ×  , where
, where  = (F1-a + F1-b)/2 and
= (F1-a + F1-b)/2 and  . Means of generations P1, P2, F1-a, F1-b, F2-a, F2-b, RILs-a, RILs-b, P1 × RILs, P2 × RILs, F1-a × RILs-a, and F1-b × RILs-b, averaged over the three experiments, were used to estimate the genetic parameters in two genetic models fitted to the data. All parameters in the models were defined according to the F2-metric (Cockerham 1954; Yang 2004). Formulas for the generation means are given in Table 1.
. Means of generations P1, P2, F1-a, F1-b, F2-a, F2-b, RILs-a, RILs-b, P1 × RILs, P2 × RILs, F1-a × RILs-a, and F1-b × RILs-b, averaged over the three experiments, were used to estimate the genetic parameters in two genetic models fitted to the data. All parameters in the models were defined according to the F2-metric (Cockerham 1954; Yang 2004). Formulas for the generation means are given in Table 1.
Model 1 included cytoplasmic, maternal, additive, and dominance effects,
 |
where Y is the mean of the generation considered; μ is the mean of the F2 generation over the three experiments, in the absence of cytoplasmic and maternal effects; c is the cytoplasmic effect attributable to seed parent P1 vs. seed parent P2; m is the maternal effect attributable to a heterozygous vs. a homozygous seed parent; [a] = Σjθjaj, with θj = −1 if P1 carries the favorable allele at locus j and +1 otherwise, and aj is the additive effect of locus j; [d] = Σjdj, with dj the dominance effect of locus j; and u, v, x, and z are generation-dependent coefficients given in Table 1.
Besides the effects defined for model 1, model 2 included epistatic effects between unlinked pairs of loci,
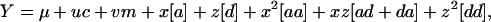 |
where [aa] = Σj<kθjθkaajk, with aajk the additive-by-additive (a × a) epistatic effect between loci j and k; [ad + da] = Σj<k{θjadjk + θkdajk}, with adjk the additive-by-dominance (a × d) epistatic effect between loci j and k and dajk the dominance-by-additive (d × a) epistatic effect between loci j and k; and [dd] = Σj<kddjk, with ddjk the dominance-by-dominance (d × d) epistatic effect between loci j and k.
In addition to the genetic effects in model 1 and model 2, we included in the generation means analyses a correction term τ (data not shown) to account for differences in the overall performance level of checks vs. RILs and TTC progenies, which were most likely attributable to differences in seed quality between the two groups of materials.
The genetic parameters for both models were estimated by weighted least squares:  = (X′WX)−1 (X′WY), where
= (X′WX)−1 (X′WY), where 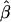 is the column vector of estimated genetic parameters, X is the design matrix with elements determined by model 1 or model 2, W is the matrix with the inverse of the variances of the generation means on the diagonal and zero on the off-diagonal, and Y is the column vector of the generation means.
is the column vector of estimated genetic parameters, X is the design matrix with elements determined by model 1 or model 2, W is the matrix with the inverse of the variances of the generation means on the diagonal and zero on the off-diagonal, and Y is the column vector of the generation means.
Standard errors of the genetic parameter estimates were calculated as the square root of the diagonal elements of matrix (X′WX)−1. Adequacy of each model was tested with a chi-square (χ2) test (Mather and Jinks 1982): χ2 =  (O − E)2 × w], where O is the observed generation mean, E is the expected generation mean based on the underlying model, and w designates the diagonal elements of matrix W, with summation over all generations considered in the model. The coefficient of determination (R2) was calculated to estimate the proportion of the variation among generation means accounted for by each model.
(O − E)2 × w], where O is the observed generation mean, E is the expected generation mean based on the underlying model, and w designates the diagonal elements of matrix W, with summation over all generations considered in the model. The coefficient of determination (R2) was calculated to estimate the proportion of the variation among generation means accounted for by each model.
Genetic variances and correlations:
Adjusted entry means of each RILn (n = 1, 2, . . ., 234) and its TTC progenies with testers P1, P2, and F1 (subsequently denoted H1n, H2n, and H3n) were used for estimating the genotypic variance (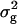 ) and associated standard errors. Since the difference between the means of populations RILs-a and RILs-b was not significant, we ignored subsets that referred to different reciprocal crosses and treated the entire set of RILs as one single population in the further analyses. The error variances for the main and subplots were calculated as combined estimates over the four types of progenies. Heritability (h2) on an entry-mean basis was estimated as h2 =
) and associated standard errors. Since the difference between the means of populations RILs-a and RILs-b was not significant, we ignored subsets that referred to different reciprocal crosses and treated the entire set of RILs as one single population in the further analyses. The error variances for the main and subplots were calculated as combined estimates over the four types of progenies. Heritability (h2) on an entry-mean basis was estimated as h2 =  /(
/( /r), where r corresponds to the number of replicates, and
/r), where r corresponds to the number of replicates, and  = main-plot error + subplot error. In the case of TTC progenies, we calculated only one estimate for h2, by averaging
= main-plot error + subplot error. In the case of TTC progenies, we calculated only one estimate for h2, by averaging 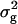 across all three types of testcross progenies.
across all three types of testcross progenies.
Additive, dominance, and epistatic variance components as defined for the F2-metric by Yang (2004) were estimated from the TTC progeny data by equating genotypic variances and covariances of H1, H2, and H3 to the corresponding linear combination of additive, dominance, and epistatic variance components given in Table 2. Six models were estimated directly with PROC MIXED, containing:  ,
, 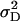 (model A);
(model A);  ,
,  ,
,  (model B);
(model B);  ,
,  ,
, 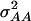 (model C);
(model C);  ,
,  ,
,  ,
,  (model D);
(model D); ,
, 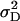 ,
, 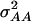 ,
, 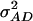 (model E); and
(model E); and 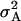 ,
,  ,
,  ,
,  ,
,  (model F), where
(model F), where 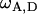 represents the covariance between additive and dominance effects. Other covariances among effects of different types as well as linkage were ignored in all models. The Bayesian information criterion (BIC) (Burnham and Anderson 2004) was applied to select among the six models. All necessary computations were performed with SAS PROC MIXED with the REML option (SAS Institute 2004).
represents the covariance between additive and dominance effects. Other covariances among effects of different types as well as linkage were ignored in all models. The Bayesian information criterion (BIC) (Burnham and Anderson 2004) was applied to select among the six models. All necessary computations were performed with SAS PROC MIXED with the REML option (SAS Institute 2004).
TABLE 2.
Variance and covariance matrix for triple testcross progenies H1, H2, and H3 under digenic epistasis, ignoring linkage and covariance terms involving epistatic effects
| Statistic |  |
 |
 |
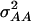 |
 |
 |
|---|---|---|---|---|---|---|
| Var(H1) |  |
1 |  |
 |
1 | 1 |
| Var(H2) |  |
1 | −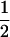
|
 |
1 | 1 |
| Var(H3) | 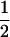 |
0 | 0 | 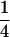 |
0 | 0 |
| Cov(H1, H2) |  |
−1 | 0 | −
|
0 | 1 |
| Cov(H1, H3) |  |
0 | 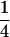 |
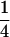 |
0 | 0 |
| Cov(H2, H3) | 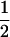 |
0 | −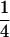
|
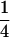 |
0 | 0 |
For description of triple testcross progenies and definition of genetic variance components see materials and methods.
The average degree of dominance was estimated as D = √ (2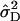 )/(
)/(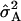 ) (Cockerham and Zeng 1996), and its standard error is according to Mood et al. (1974). Alternatively, we estimated the overall dominance ratio DR =
) (Cockerham and Zeng 1996), and its standard error is according to Mood et al. (1974). Alternatively, we estimated the overall dominance ratio DR =  from first-degree statistics as
from first-degree statistics as
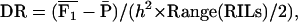 |
where Range(RILs) refers to the range across the entire set of n = 234 RILs, with regard to their per se performance. The denominator was chosen because in the absence of epistasis and linkage, the genetic expectation of Range(RILs) converges toward  for n → ∞.
for n → ∞.
Genotypic correlations among traits were estimated in RILs as well as their TTC progenies averaged over the three testers and between RILs and the mean of their TTC progenies. These computations were performed on the basis of adjusted entry means and with software PLABSTAT (Utz 2000).
RESULTS
Generation means:
Parental lines P1 and P2 differed significantly (P < 0.01) in RD29, GR, and DMC (Table 3). Significant (P < 0.01) differences were also observed between F1-a and F1-b in RD22, RD29, and GR. Differences between F2-a and F2-b, as well as between the means of RILs-a and RILs-b were not significant in any of the traits (data not shown). Generation F1 surpassed significantly (P < 0.01) both parents in RD22, RD29, GR, and BY. MPH was highest (49%) for BY and exceeded 23% in all traits but DMC.
TABLE 3.
Means and associated standard errors in generations P1, P2, F1, F2, RILs and triple testcross progenies with testers P1, P2, and F1 as well as range of the RILs and dominance ratio for five biomass-related traits in Arabidopsis hybrid C24 × Col-0
| Generations | Na | RD22 (mm) | RD29 (mm) | GR (mm/day) | DMC (%) | BY (mg) |
|---|---|---|---|---|---|---|
| Checks | ||||||
| P1 | 18 | 22.3 ± 0.75 | 59.1 ± 1.45 | 5.27 ± 0.15 | 7.66 ± 0.11 | 53.8 ± 4.09 |
| P2 | 18 | 24.0 ± 0.75 | 67.8 ± 1.45 | 6.26 ± 0.15 | 8.15 ± 0.11 | 62.1 ± 4.03 |
 b b
|
18 | 28.4 ± 0.89 | 79.9 ± 1.75 | 7.39 ± 0.18 | 7.76 ± 0.13 | 86.6 ± 4.75 |
 b b
|
18 | 25.3 ± 0.75 | 71.6 ± 1.45 | 6.65 ± 0.15 | 7.85 ± 0.11 | 69.0 ± 4.03 |
| MPHc | 22.7 ± 2.37 | 25.9 ± 1.21 | 28.2 ± 4.21 | −1.83 ± 3.07 | 49.4 ± 1.94 | |
| Entries | ||||||
| RILs | 234 | 25.8 ± 0.27 | 66.9 ± 0.53 | 5.94 ± 0.05 | 8.34 ± 0.04 | 76.6 ± 1.37 |
| P1 × RILs (H1) | 234 | 29.9 ± 0.45 | 76.0 ± 0.82 | 6.65 ± 0.06 | 7.51 ± 0.04 | 93.3 ± 2.30 |
| P2 × RILs (H2) | 234 | 30.5 ± 0.44 | 78.7 ± 0.74 | 6.92 ± 0.06 | 8.09 ± 0.04 | 103.0 ± 2.43 |
| F1 × RILs (H3) | 234 | 30.7 ± 0.46 | 78.5 ± 0.69 | 6.87 ± 0.05 | 7.83 ± 0.04 | 97.7 ± 2.31 |
| Ranged | ||||||
| RILs | 33.45 | 61.26 | 5.25 | 5.54 | 165.68 | |
| Dominance ratio | ||||||
| DRe | 0.51 | 0.60 | 0.72 | −0.04 | 0.99 | |
Number of entries.
F1 and F2 were calculated as means across both reciprocal forms.
Midparent heterosis (MPH) = 100 ×  , where
, where 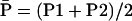 , with standard error calculated according to Mood et al. (1974).
, with standard error calculated according to Mood et al. (1974).
Range(RILs) = Max(RIL) − Min(RIL).
DR =  /(h2 × Range(RILs)/2).
/(h2 × Range(RILs)/2).
Testcross progenies outperformed (P < 0.01) the RILs in all traits except DMC (Table 3). The mean of P2 × RILs significantly surpassed (P < 0.01) that of P1 × RILs in all traits except RD22. Compared with F1 × RILs, P1 × RILs reached significantly (P < 0.01) lower values in all traits except RD22 and BY. Compared with F1 × RILs, P2 × RILs reached significantly (P < 0.01) higher values in DMC and P1 × RILs had significantly (P < 0.05) smaller values for GR and DMC. The F1 × RILs deviated significantly (P < 0.05) from the overall mean of P1 × RILs and P2 × RILs only for RD29 (data not shown). The range in the variation of the RILs exceeded by far the difference of the parents, indicating transgressive segregation in all traits. As expected, the dominance ratio DR was lowest (−0.04) for DMC, highest (0.99) for BY, and intermediate (between 0.51 and 0.72) for the other traits.
Model 1 accounted for 84–98% of the variation among generation means for all traits (Table 4). Nevertheless, the χ2-values for the goodness-of-fit of model 1 were highly significant (P < 0.01) for DMC. Neither cytoplasmic nor maternal effects were significant (P < 0.01) (data not shown). Inclusion of epistatic effects (model 2) considerably improved the fit, explaining from 97 to almost 100% of the variation among generation means for all traits. However, the χ2-value of model 2 was still significant (P < 0.05) for DMC.
TABLE 4.
Genetic effects and associated standard errors for five biomass-related traits estimated by generation means analysis in Arabidopsis hybrid C24 × Col-0
| Parameters | RD22 (mm) | RD29 (mm) | GR (mm/day) | DMC (%) | BY (mg) |
|---|---|---|---|---|---|
| Model 1 | |||||
| μ | 28.04 ± 0.43** | 74.85 ± 0.66** | 6.72 ± 0.05** | 7.78 ± 0.10** | 84.78 ± 1.78** |
| [a] | 0.75 ± 0.69 | 3.57 ± 1.06* | 0.36 ± 0.08** | 0.46 ± 0.15* | 6.47 ± 2.92 |
| [d] | 7.67 ± 1.02** | 19.99 ± 1.58** | 1.73 ± 0.11** | −0.80 ± 0.25* | 37.57 ± 4.19** |
 a a
|
11.52* | 8.03 | 4.82 | 40.92** | 7.19 |
| R2 (%) | 95.3 | 97.9 | 98.5 | 83.6 | 97.0 |
| Model 2 | |||||
| μ | 27.51 ± 0.46** | 74.32 ± 0.93** | 6.73 ± 0.06** | 7.92 ± 0.08** | 82.44 ± 0.87** |
| [a] | 0.59 ± 0.91 | 2.60 ± 1.71 | 0.27 ± 0.08 | 0.58 ± 0.11* | 9.75 ± 1.73* |
| [d] | 8.05 ± 0.89* | 20.37 ± 1.77** | 1.73 ± 0.10** | −0.84 ± 0.15* | 39.17 ± 1.65** |
| [aa] | 1.93 ± 1.04 | 1.92 ± 2.11 | −0.05 ± 0.13 | −0.43 ± 0.18 | 8.72 ± 1.97* |
| [ad + da] | −0.52 ± 2.39 | −3.49 ± 4.67 | −0.45 ± 0.27 | 0.67 ± 0.37 | 11.18 ± 4.55 |
 a a
|
4.22 | 4.81 | 1.92 | 7.35* | 0.53 |
| R2 (%) | 98.3 | 98.8 | 99.4 | 97.1 | 99.8 |
For description of traits see materials and methods. *Significant at P < 0.05 and **significant at P < 0.01.
Chi-square degrees of freedom are in parentheses.
Both models yielded similar estimates of [a] and [d] (Table 4). Dominance effects were significant (P < 0.01) and had a positive sign for all traits except for DMC. Additive effects were consistently smaller in magnitude and under model 2 significant (P < 0.05) only for DMC and BY. Estimates of [aa] under model 2 were significantly (P < 0.05) positive for BY. The estimates of [ad + da] were not significant for any trait and the same applied to estimates of [dd] (data not shown).
Variances and heritabilities:
Genotypic variances (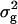 ) among RILs were highly significant (P < 0.01) for per se and testcross performance with all three testers for all biomass-related traits (Table 5). Surprisingly, estimates of
) among RILs were highly significant (P < 0.01) for per se and testcross performance with all three testers for all biomass-related traits (Table 5). Surprisingly, estimates of  in RILs were smaller than those in their TTC progenies for RD22 and BY, of similar size for RD29, but significantly (P < 0.01) larger for GR and DMC. Heritability (h2) estimates for the RILs and their TTC progenies were in most instances high and ranged between 76 and 84%, except for GR and DMC in the testcrosses.
in RILs were smaller than those in their TTC progenies for RD22 and BY, of similar size for RD29, but significantly (P < 0.01) larger for GR and DMC. Heritability (h2) estimates for the RILs and their TTC progenies were in most instances high and ranged between 76 and 84%, except for GR and DMC in the testcrosses.
TABLE 5.
Variance components ( , genotypic variance;
, genotypic variance;  , error variance) and heritability (h2) estimates with associated standard errors for five biomass-related traits in RILs and their triple testcross progenies with testers P1, P2, and F1 in Arabidopsis hybrid C24 × Col-0
, error variance) and heritability (h2) estimates with associated standard errors for five biomass-related traits in RILs and their triple testcross progenies with testers P1, P2, and F1 in Arabidopsis hybrid C24 × Col-0
| Parameters | RD22 | RD29 | GR | DMC | BY |
|---|---|---|---|---|---|
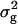 (RILs) (RILs) |
37.10 ± 4.50** | 145.53 ± 17.37** | 1.076 ± 0.140** | 1.004 ± 0.117** | 874 ± 112** |
 (H1)a (H1)a
|
49.51 ± 5.60** | 156.03 ± 18.14** | 0.682 ± 0.104** | 0.237 ± 0.047** | 1109 ± 133** |
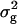 (H2)b (H2)b
|
50.76 ± 5.79** | 120.75 ± 15.31** | 0.535 ± 0.091** | 0.219 ± 0.051** | 1557 ± 176** |
 (H3)c (H3)c
|
44.32 ± 5.14** | 100.87 ± 13.11** | 0.421 ± 0.078** | 0.160 ± 0.039** | 1073 ± 131** |
 |
28.42 ± 0.89 | 98.77 ± 3.20 | 1.018 ± 0.033 | 0.663 ± 0.021 | 833 ± 26 |
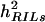 (%) (%) |
79.66 ± 2.13 | 81.55 ± 1.95 | 76.02 ± 2.56 | 81.96 ± 1.89 | 75.88 ± 2.55 |
 (%) (%) |
83.57 ± 1.28 | 79.27 ± 1.69 | 61.68 ± 3.06 | 48.21 ± 4.44 | 81.78 ± 1.41 |
* Significant at P < 0.05 and ** significant at P < 0.01.
H1 denotes the testcross progenies of RILs with tester P1.
H2 denotes the testcross progenies of RILs with tester P2.
H3 denotes the testcross progenies of RILs with tester F1.
Additive (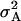 ) and dominance (
) and dominance ( ) variances were highly significant (P < 0.01) for all traits in all six models studied (data not shown). Regarding models without epistasis, model B (data not shown) provided a similar fit to the data as model A did, as reflected by the BIC values and estimates of
) variances were highly significant (P < 0.01) for all traits in all six models studied (data not shown). Regarding models without epistasis, model B (data not shown) provided a similar fit to the data as model A did, as reflected by the BIC values and estimates of  , which were not significant (P < 0.05) except for RD29. For models including epistasis, model D (including
, which were not significant (P < 0.05) except for RD29. For models including epistasis, model D (including  ,
,  ,
,  , and
, and 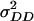 ) yielded the lowest BIC value for all traits. Hence, presentation of the results in Table 6 is restricted to models A and D.
) yielded the lowest BIC value for all traits. Hence, presentation of the results in Table 6 is restricted to models A and D.
TABLE 6.
Estimates of additive, dominance, and epistatic variance components with associated standard errors, as well as average degree of dominance for five biomass-related traits in Arabidopsis hybrid C24 × Col-0
| Parameters | RD22 | RD29 | GR | DMC | BY |
|---|---|---|---|---|---|
| Model A | |||||
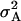 |
65.00 ± 7.29** | 179.99 ± 20.86** | 0.762 ± 0.112** | 0.393 ± 0.057** | 1479 ± 178** |
 |
11.32 ± 1.57** | 34.33 ± 4.69** | 0.211 ± 0.034** | 0.0832 ± 0.014** | 321 ± 45** |
| Da | 0.59 | 0.62 | 0.74 | 0.65 | 0.66 |
| BICb | 14924 | 16309 | 6512 | 5278 | 22223 |
| Model D | |||||
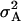 |
71.14 ± 8.79** | 169.61 ± 21.79** | 0.719 ± 0.113** | 0.361 ± 0.058** | 1656 ± 223** |
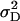 |
10.48 ± 1.58** | 34.90 ± 4.67** | 0.216 ± 0.033** | 0.080 ± 0.015** | 309 ± 45** |
 |
5.25 ± 0.23** | 3.89 ± 0.18** | 0.000 ± 0.000 | 0.020 ± 0.001** | 152 ± 7** |
 |
25.63 ± 3.37** | 39.68 ± 7.00** | 0.070 ± 0.037 | 0.048 ± 0.019** | 854 ± 107** |
| Da | 0.54 | 0.64 | 0.77 | 0.67 | 0.61 |
| BICb | 14742 | 16251 | 6519 | 5280 | 21986 |
* Significant at P < 0.05 and ** significant at P < 0.01.
D = √(2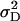 /
/ ).
).
The Bayesian information criterion.
In general, estimates of  were approximately four to six times larger than those of
were approximately four to six times larger than those of 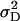 under both models (Table 6). Inclusion of epistatic variance components improved the fit of model D, as reflected by the reduced BIC values for all traits but GR and DMC. Estimates of
under both models (Table 6). Inclusion of epistatic variance components improved the fit of model D, as reflected by the reduced BIC values for all traits but GR and DMC. Estimates of 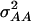 were highly significant (P < 0.01) and up to one-half of those for
were highly significant (P < 0.01) and up to one-half of those for  for all traits except GR. Estimates of
for all traits except GR. Estimates of  ranged between one-quarter and one-half of those for
ranged between one-quarter and one-half of those for 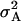 , except for GR and DMC, where
, except for GR and DMC, where  was only one-eighth of
was only one-eighth of  .
.
Estimates of the average degree of dominance D were similar for models A and D except for RD22 and BY, where the inclusion of  and
and  in the model resulted in reduced values (Table 6). D-values ranged between 0.54 and 0.77 and were of intermediate size for BY.
in the model resulted in reduced values (Table 6). D-values ranged between 0.54 and 0.77 and were of intermediate size for BY.
Correlations:
Genotypic correlations (rg) among traits were consistently higher in the RILs than in the TTC progenies averaged over the three testers (Table 7). Estimates of rg among RD22, RD29, GR, and BY were positive and high, ranging from 0.51 to 0.95, with a single exception. DMC was negatively correlated with all other biomass-related traits. Genotypic correlations between RILs and the mean of their TTC progenies ranged between 0.62 and 0.80 for the studied traits.
TABLE 7.
Genotypic correlations among five biomass-related traits in RILs (above diagonal), the mean of their triple testcross progenies averaged over the three testers (below diagonal), and that between both these measures (on the diagonal in italics) in Arabidopsis hybrid C24 × Col-0
| RD22 | RD29 | GR | DMC | BY | |
|---|---|---|---|---|---|
| RD22 | 0.66a | 0.94a | 0.73a | −0.53a | 0.95a |
| RD29 | 0.84a | 0.73a | 0.92a | −0.66a | 0.89a |
| GR | 0.51a | 0.90a | 0.77a | −0.67a | 0.67a |
| DMC | −0.28a | −0.53a | −0.63a | 0.80a | −0.39a |
| BY | 0.87a | 0.66a | 0.38a | −0.08 | 0.62a |
Genotypic correlation exceeded twice its standard error.
DISCUSSION
Choice of the experimental design:
Design III and the TTC design are most suitable for investigations of heterosis in the presence of epistasis because they provide estimates of augmented dominance effects in the terminology of A. E. Melchinger, H. F. Utz, H. P. Piepho and C. C. Schön (unpublished results). These effects capture the effects of dominance as well as the sum of a × a epistatic interactions of a QTL with the genetic background contributing to heterosis under the F2-metric. Instead of F2 plants or F3 lines used in design III studies with maize, we used RILs as parents for producing the testcross progenies following Kearsey et al. (2003). Use of homozygous parents maximizes the genetic variance among testcross progenies and leads to an increased power in F-tests as well as smaller standard errors of variance components estimates and D (A. E. Melchinger, H. F. Utz, H. P. Piepho and C. C. Schön, unpublished results). Use of RILs has the further advantage that the effects of linkage are reduced because linkage disequilibrium between tightly linked loci is almost halved compared with that in F2 plants or double-haploid lines.
Although the experimental design we applied possesses the above-mentioned advantages, we used a considerably larger sample of RILs and number of plants per RIL or TTC progenies than in previous studies to achieve a high level of precision in parameter estimates. For genetic variance components and h2, standard errors were generally small in comparison with the estimates, but for D they were of the same order.
Choice of the F2-metric:
Three different metrics have been proposed in the literature (Van der Veen 1959): (i) the F∞-metric (Hayman 1954); (ii) the F2-metric, which is a special case of Cockerham's (1954) model; and (iii) the mixed metric (Hayman and Mather 1955), which is a mixture of Cockerham's model and the F∞-metric. While the F2-metric yields orthogonal parameters with regard to an F2 population, the same applies to the F∞-metric with regard to homozygous lines. Following the definition and terminology of Melchinger (1987), the progenies of design III and the TTC design can be considered as gamete-orthogonal populations. Gene effects in a gamete-orthogonal population are commonly defined with regard to the respective gene-orthogonal population, which corresponds to the F2 population in the case of a TTC design.
For this study, we chose the F2-metric for three reasons. First, it has the advantage that each variance component is proportional to the sum of squares of the corresponding genetic effects (e.g., 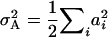 ,
, 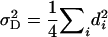 ,
,  , etc.; Yang 2004) and does not involve any other type of genetic effects that could obscure their interpretation. This would not apply if we had chosen the F∞-metric, as shown by Cheverud (2000). Second, the F2-metric underlies the genetic model used in testing for epistatic interactions among QTL by two-way ANOVAs for pairs of marker loci (Cockerham and Zeng 1996). Third, with digenic epistasis, MPH = [d] − [aa], i.e., besides dominance effects it involves only a × a effects, whereas under the F∞-metric MPH is additionally influenced by d × d effects (Van Der Veen 1959).
, etc.; Yang 2004) and does not involve any other type of genetic effects that could obscure their interpretation. This would not apply if we had chosen the F∞-metric, as shown by Cheverud (2000). Second, the F2-metric underlies the genetic model used in testing for epistatic interactions among QTL by two-way ANOVAs for pairs of marker loci (Cockerham and Zeng 1996). Third, with digenic epistasis, MPH = [d] − [aa], i.e., besides dominance effects it involves only a × a effects, whereas under the F∞-metric MPH is additionally influenced by d × d effects (Van Der Veen 1959).
Cytoplasmic and maternal effects:
While reciprocal differences in some traits were observed in the F1 generation, cytoplasmic effects were consistently not significant in the generation means analysis. One simple explanation, that both parents have the same cytoplasm, can be excluded because Forner et al. (2005) found differences in mtDNA between C24 and Col-0.
Maternal effects due to heterozygosity of seed parent were observed for grain and forage yield in maize (Melchinger et al. 1986). We found no such effect in Arabidopsis and, hence, excluded the respective parameter from the model. Possible explanations for this observation are that there is only marginal heterosis for seed weight in Arabidopsis and uniform seed quality in our study was achieved by restricting pollinations and selfing to six siliques per plant.
Interpretation of first- and second-degree statistics:
Our generation means analysis extends the ordinary generation means analysis based on the parents, F1, selfing, and backcross generations from biparental crosses to the populations of RILs and testcross progenies, underlying the test for epistasis in the TTC design (Kearsey and Jinks 1968). In both analyses, the parameters capture the net contribution of gene effects. However, the consequences of summation over loci are quite different for various parameters. If directional dominance (di > 0) prevails, as suggested by most QTL studies on traits related to plant growth, the sum in [d] involves mainly positive terms. In contrast, if the parents have similar performance levels, the sum in [a] involves both positive and negative terms that cancel each other to a substantial amount. Because the range in the RILs surpassed by far the absolute difference |P1 − P2| for all traits, we conclude that θi, the sign of the additive effect ai, was not uniform across the QTL involved in trait expression. Significance of epistatic effects [aa], [ad + da], or [dd] is indicative of their presence but does not provide any clue on their relative importance. Likewise, absence of significant estimates for [aa], [ad + da], or [dd] does not preclude the existence of these types of effects, because positive and negative effects may be canceled in the sum.
Under the F2-metric, this shortcoming is overcome by estimation of variance components because they represent sums of squared effects at individual loci except in the case of linkage among loci. Under directional dominance, linkage causes an inflation in  , whereas
, whereas  is inflated by the covariance of linked QTL in coupling phase and deflated by those in repulsion phase with corresponding carryover for estimates of D (Comstock and Robinson 1952).
is inflated by the covariance of linked QTL in coupling phase and deflated by those in repulsion phase with corresponding carryover for estimates of D (Comstock and Robinson 1952).
In the presence of epistasis, estimates of  and
and  obtained from design III are confounded by those of
obtained from design III are confounded by those of 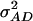 and
and  , respectively, and to a minor extent also influenced by other types of epistatic terms (A. E. Melchinger, H. F. Utz, H. P. Piepho and C. C. Schön, unpublished results). With the TTC design, it is possible to separate epistatic variance components from those of
, respectively, and to a minor extent also influenced by other types of epistatic terms (A. E. Melchinger, H. F. Utz, H. P. Piepho and C. C. Schön, unpublished results). With the TTC design, it is possible to separate epistatic variance components from those of  and
and  , but one cannot clearly discriminate between
, but one cannot clearly discriminate between  and
and 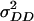 . This is because coefficients for these two variances are almost identical in the genetic expectation of the (co)variance of H1n, H2n, and H3n (Table 2). Hence, model D and model E yielded similar estimates of
. This is because coefficients for these two variances are almost identical in the genetic expectation of the (co)variance of H1n, H2n, and H3n (Table 2). Hence, model D and model E yielded similar estimates of  and
and 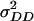 besides almost identical estimates of
besides almost identical estimates of  ,
,  , and
, and 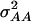 .
.
The comparison between RILs and their TTC progenies provides further evidence for the presence of epistasis. In the absence of epistasis, the genotypic correlation between the performance of RILs per se and the means of their TTC progenies is expected to be 1.0. Furthermore, 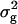 of RILs is expected to be twice as large as
of RILs is expected to be twice as large as  of testcross progenies with the F1 tester (A. E. Melchinger, H. F. Utz, H. P. Piepho and C. C. Schön, unpublished results). Estimates of these parameters were closest to these expectations for DMC and GR, but deviated substantially for all other biomass-related traits, most notably for BY.
of testcross progenies with the F1 tester (A. E. Melchinger, H. F. Utz, H. P. Piepho and C. C. Schön, unpublished results). Estimates of these parameters were closest to these expectations for DMC and GR, but deviated substantially for all other biomass-related traits, most notably for BY.
We estimated the average dominance ratio Di = di/ai across all QTL by D and DR. In both statistics, QTL with large ai and di effects receive a higher weight than those with smaller effects. Both statistics are inflated by linked QTL with additive effects in repulsion phase (θi ≠ θj). In contrast to D, which is always positive and provides a realistic estimate of the average dominance ratio only in the case of directional dominance (di ≥ 0), DR takes into account different signs in di. This explains the large discrepancy between DR and D for DMC and suggests that almost half of the QTL influencing this trait have negative sign of di. Conversely, the good agreement between D and DR for the other traits suggests a prevalence of directional dominance at the majority of QTL. The difference in size between D and DR for BY could be regarded as an indicator that our sample of 234 RILs was too small to represent adequately the extreme genotypes for this complex polygenic trait.
Level of heterosis in Arabidopsis hybrid C24 × Col-0:
In agreement with other studies on Arabidopsis (Barth et al. 2003; Meyer et al. 2004), we detected medium to high MPH for biomass-related traits with differences in the absolute values of MPH due to different light intensities and recording or harvesting times. MPH for BY was about twice as large as for RD. This result is in harmony with the hypothesis of Williams (1959) that multiplication effects are a major cause of heterosis in complex traits. In our study, RD reflects plant growth in just one dimension, namely leaf length, whereas BY depends in multiplicative fashion also on leaf width and leaf number as well as on other features of biomass accumulation, each of which presumably displays a small amount of heterosis.
Growth rate displayed a medium amount of MPH. However, because biomass accumulation follows an exponential curve during the early stages of plant development, a small amount of heterosis for GR translates into much bigger differences among the hybrid and the parental lines for BY.
No significant MPH was observed for DMC in our study. This finding is in contrast to negative MPH observed for DMC of grain in maize (Mihaljevic et al. 2005) due to earlier flowering and a more rapid vegetative and generative growth of hybrids. Although MPH for DMC has not been investigated in other studies with Arabidopsis, Barth et al. (2003) and Kearsey et al. (2003) also reported only a low magnitude of MPH for flowering date.
Genetic basis of heterosis for biomass-related traits in Arabidopsis hybrid C24 × Col-0:
We observed positive estimates of [aa] for all biomass-related traits except for GR and DMC. This indicates that pairs of genes at nonhomologous loci, which show positive epistatic interactions for biomass-related traits, have been accumulated in the parents C24 and Col-0 presumably as a result of natural selection. These gene effects reduce MPH, which is evident from the relationship MPH = [d] − [aa]. Therefore, they cannot be exploited in the hybrid. In the TTC progenies, the term 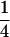 [aa] favors the average of testcross progenies with testers P1 and P2 over the mean of testcross progenies with tester F1, as detected by the test for epistasis in the TTC analysis devised by Kearsey and Jinks (1968).
[aa] favors the average of testcross progenies with testers P1 and P2 over the mean of testcross progenies with tester F1, as detected by the test for epistasis in the TTC analysis devised by Kearsey and Jinks (1968).
Significant estimates of  support the presence of epistasis of type a × a for all biomass-related traits except for GR (Table 6). In the case of RD22 and BY, these estimates amounted to half the size of those for
support the presence of epistasis of type a × a for all biomass-related traits except for GR (Table 6). In the case of RD22 and BY, these estimates amounted to half the size of those for  , suggesting that the epistasis of type a × a is a major factor influencing heterosis for biomass-related traits in hybrid C24 × Col-0. Estimates of
, suggesting that the epistasis of type a × a is a major factor influencing heterosis for biomass-related traits in hybrid C24 × Col-0. Estimates of 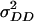 in model D were consistently of greater importance than estimates of
in model D were consistently of greater importance than estimates of  and of equal (RD29) or about twice the size (RD22, BY) of estimates of
and of equal (RD29) or about twice the size (RD22, BY) of estimates of  . Epistasis of types a × d and d × d does not contribute to MPH. However, it cannot be precluded that estimates of
. Epistasis of types a × d and d × d does not contribute to MPH. However, it cannot be precluded that estimates of  and
and  account also for higher-order types of epistatic effects that influence heterosis. Hence, digenic and higher-order epistatic effects seem to play an important role in heterosis for biomass-related traits. This is in agreement with the study of Cockerham and Zeng (1996), who reported significant epistasis among linked QTL besides dominance of favorable genes contributing to heterosis for grain yield and other traits in maize hybrid B73 × Mo17. In accordance with the studies on maize, Hua et al. (2002) and Li et al. (2001) detected substantial digenic epistasis in various populations of rice. Li et al. (2001) reported that most of the variance for BY was due to epistatic QTL.
account also for higher-order types of epistatic effects that influence heterosis. Hence, digenic and higher-order epistatic effects seem to play an important role in heterosis for biomass-related traits. This is in agreement with the study of Cockerham and Zeng (1996), who reported significant epistasis among linked QTL besides dominance of favorable genes contributing to heterosis for grain yield and other traits in maize hybrid B73 × Mo17. In accordance with the studies on maize, Hua et al. (2002) and Li et al. (2001) detected substantial digenic epistasis in various populations of rice. Li et al. (2001) reported that most of the variance for BY was due to epistatic QTL.
Estimates of D were of medium size for all biomass-related traits. This is in sharp contrast to most reports on design III studies with F2 populations in maize, where estimates of D generally exceeded 1.0 (Wolf et al. 2000). Possible explanations are that the dominance ratio Di at individual loci is much smaller in Arabidopsis than in maize and the same may generally hold true for autogamous vs. allogamous crops, as reflected by the different levels of MPH displayed by each group of species (Becker 1993).
The medium estimates of D and DR suggest that gene action at the majority of loci in the cross C24 × Col-0 is in the range of partial to complete dominance. However, because D averages over loci, it cannot be ruled out that gene action is purely additive at the majority of loci and only a minority of loci displays dominance or even (pseudo)overdominance. Discrimination among these various hypotheses on the genetic causes of heterosis in Arabidopsis is not possible by first- or second-degree statistics alone but requires mapping individual QTL involved in the expression of heterosis. This research will also provide information about the potential linkage bias in the estimates of D and DR.
In conclusion, Arabidopsis and particularly its hybrid C24 × Col-0 are an excellent plant resource to satisfy many requirements for analysis of the genetic causes of heterosis. The hybrid displays a substantial amount of biomass-related hybrid vigor. Furthermore, a high-density map has been established (Törjék et al. 2006) and also metabolic profile data have been collected (Meyer et al. 2007). Therefore, several follow-up studies with these materials are underway to gain a deeper understanding of the physiological, genetic, and molecular causes of heterosis by QTL mapping as well as analysis of transcript, protein, and metabolite profiles.
Acknowledgments
We gratefully acknowledge the expert technical assistance of B. Devezi-Savula, N. Friedl, M. Zeh, M. Teltow, and C. Marona. We are indebted to C. C. Schön for critical reading and valuable suggestions for improving the manuscript. This project was supported by the Deutsche Forschungsgemeinschaft (German Research Foundation) under the priority research program “Heterosis in Plants” (research grants ME931/4-1, ME931/4-2, AL387/6-1, and AL387/6-2).
References
- Barth, S., A. K. Busimi, H. F. Utz and A. E. Melchinger, 2003. Heterosis for biomass yield and related traits in five hybrids of Arabidopsis thaliana L. Heynh. Heredity 91: 36–42. [DOI] [PubMed] [Google Scholar]
- Becker, H. C., 1993. Pflanzenzüchtung. Eugen Ulmer Verlag, Stuttgart, Germany.
- Burnham, K. P., and D. R. Anderson, 2004. Multimodel inference: understanding AIC and BIC in model selection. Sociol. Meth. Res. 33: 261–304. [Google Scholar]
- Cheverud, J. M., 2000. Detecting epistasis among quantitative trait loci, pp. 58–81 in Epistasis and the Evolutionary Process, edited by J. B. Wolf, E. D. Brodiee and M. J. Wade. Oxford University Press, New York.
- Cockerham, C. C., 1954. An extension of the concept of partitioning hereditary variance for analysis of covariances among relatives when epistasis is present. Genetics 39: 859–882. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cockerham, C. C., and Z. B. Zeng, 1996. Design III with marker loci. Genetics 143: 1437–1456. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Comstock, R. E., and H. F. Robinson, 1952. Estimation of average dominance of genes, pp. 494–516 in Heterosis, edited by J. W. Growen. Iowa State College Press, Ames, IA.
- Forner, J., B. Weber, C. Wiethölter, R. Meyer and S. Binder, 2005. Distant sequences determine 5′ end formation of cox3 transcripts in Arabidopsis thaliana ecotype C24. Nucleic Acids Res. 33: 4673–4682. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hallauer, A. R., and J. B. Miranda, 1981. Quantitative Genetics in Maize Breeding. Iowa State University Press, Ames, IA.
- Hayman, B. I., 1954. A mathematical theory of gene action and interaction. Ph.D. Thesis, University of Birmingham, Birmingham, UK.
- Hayman, B. I., and K. Mather, 1955. The description of genetic interactions in continuous variation. Biometrics 11: 69–82. [Google Scholar]
- Hinze, L. L., and K. R. Lamkey, 2003. Absence of epistasis for grain yield in elite maize hybrids. Crop Sci. 43: 46–56. [Google Scholar]
- Hua, J., Y. Xing, W. Wu, C. Xu, X. Sun et al., 2002. Genetic dissection of an elite rice hybrid revealed that heterozygotes are not always advantageous for performance. Genetics 162: 1885–1895. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hua, J., Y. Xing, W. Wu, C. Xu, X. Sun et al., 2003. Single-locus heterotic effects and dominance by dominance interactions can adequately explain the genetic basis of heterosis in an elite rice hybrid. Proc. Natl. Acad. Sci. USA 100: 2574–2579. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kearsey, M. J., and J. L. Jinks, 1968. A general method of detecting additive, dominance and epistatic variation for metrical traits. I. Theory. Heredity 23: 403–409. [DOI] [PubMed] [Google Scholar]
- Kearsey, M. J., H. S. Pooni and N. H. Syed, 2003. Genetics of quantitative traits in Arabidopsis thaliana. Heredity 91: 456–464. [DOI] [PubMed] [Google Scholar]
- Lamkey, K. R., and J. W. Edwards, 1999. The quantitative genetics of heterosis, pp. 31–48 in The Genetics and Exploitation of Heterosis in Crops, edited by J. G. Coors and S. Pandey. CCSA/ASA/SSA, Madison, WI.
- Li, Z.-K, L. J. Luo, H. W. Mei, D. L. Wang, Q. Y. Shu et al., 2001. Overdominant epistatic loci are the primary genetic basis of inbreeding depression and heterosis in rice. I. Biomass and grain yield. Genetics 158: 1737–1753. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Luo, L. J., Z.-K Li, H. W. Mei, Q. Y. Shu, R. Tabien et al., 2001. Overdominant epistatic loci are the primary genetic basis of inbreeding depression and heterosis in rice. II. Grain yield components. Genetics 158: 1755–1771. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mather, K., and J. L. Jinks, 1982. Biometrical Genetics, Ed. 3. Chapman & Hall, London.
- Melchinger, A. E., 1987. Expectation of means and variances of test-crosses produced from F2 and backcross individuals and their selfed progenies. Heredity 59: 105–115. [Google Scholar]
- Melchinger, A. E., H. H. Geiger and F. W. Schnell, 1986. Epistasis in maize (Zea mays L.) 2. Genetic effects in crosses among early flint and dent inbred lines determined by three methods. Theor. Appl. Genet. 72: 231–239. [DOI] [PubMed] [Google Scholar]
- Meyer, R. C., O. Törjék, M. Becher and T. Altmann, 2004. Heterosis of biomass production in Arabidopsis. Establishment during early development. Plant Physiol. 134: 1813–1823. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meyer, R. C., M. Steinfath, J. Lisec, M. Becher, H. Witucka-Wall et al., 2007. Metabolic signature of Arabidopsis plants allows prediction of biomass. Proc. Natl. Acad. Sci. USA 104: 4759–4764. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mihaljevic, R., H. F. Utz and A. E. Melchinger, 2005. No evidence for epistasis in hybrid and per se performance of elite European flint maize inbreds from generation means and QTL analyses. Crop Sci. 45: 2605–2613. [Google Scholar]
- Mood, A. M., F. Graybill and D. C. Boes, 1974. Introduction to the Theory of Statistics, Ed. 3. McGraw-Hill, Boston.
- SAS Institute, 2004. SAS Version 9.1. SAS Institute, Cary, NC.
- Simmonds, N. W., and J. Smartt, 1999. Principles of Crop Improvement, Ed. 2. Blackwell Science, Oxford.
- Törjék, O., H. Witucka-Wall, R. C. Meyer, M. von Korff, B. Kusterer et al., 2006. Segregation distortion in Arabidopsis C24/Col-0 and Col-0/C24 recombinant inbred line populations is due to reduced fertility caused by epistatic interaction of two loci. Theor. Appl. Genet. 113: 1551–1561. [DOI] [PubMed] [Google Scholar]
- Utz, H. F., 2000. PLABSTAT: A Computer Program for Statistical Analysis of Plant Breeding Experiments. Institute of Plant Breeding, Seed Science and Population Genetics, University of Hohenheim, Stuttgart, Germany.
- Van Der Veen, J. H., 1959. Tests of non-allelic interaction and linkage for quantitative characters in generations derived from two diploid pure lines. Genetica 30: 201–232. [DOI] [PubMed] [Google Scholar]
- Williams, W., 1959. Heterosis and the genetics of complex characters. Nature 184: 527–530. [DOI] [PubMed] [Google Scholar]
- Wolf, D. P., L. A. Peternelli and A. R. Hallauer, 2000. Estimates of genetic variance in an F2 maize population. J. Hered. 91: 384–391. [DOI] [PubMed] [Google Scholar]
- Xiao, J., J. Li, L. Yuan and S. D. Tanksley, 1995. Dominance is the major genetic basis of heterosis in rice as revealed by QTL analysis using molecular markers. Genetics 140: 745–754. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang, R. C., 2004. Epistasis of quantitative trait loci under different gene action models. Genetics 167: 1493–1505. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yu, S. B., J. X. Li, C. G. Xu, Y. F. Tan, Y. J. Gao et al., 1997. Importance of epistasis as the genetic basis of heterosis in an elite rice hybrid. Proc. Natl. Acad. Sci. USA 94: 9226–9231. [DOI] [PMC free article] [PubMed] [Google Scholar]