

Abstract
We modeled the behavior of recessive mutations with deleterious effects to either the sporophyte or the gametophyte, or both, in polysomic tetraploid populations by allowing for varying levels of double reduction, mutation, and self-fertilization. Double reduction causes a decrease of the equilibrium frequencies of deleterious alleles, and it has much more influence on genes subjected to gametophytic selection than on genes solely under sporophytic selection. With gametophytic selection, low frequencies of double reduction are enough to reduce equilibrium frequencies severalfold. Double reduction occurs when sister alleles migrate to the same gamete during meiosis. It depends on the frequency at which a locus recombines with its centromere, and on the frequency of multivalent formation. Therefore, a greater accumulation of deleterious mutations should occur on polysomic chromosomes with a prevalence of bivalent pairing and in chromosomal regions between centromeres and proximal chiasmata. Proximal loci should have a greater impact in reducing the fitness of a polyploid population being inbred. This prediction can explain observations that homozygosities at different subchromosomal regions have distinct effects on inbreeding depression in polyploids. Furthermore, even mildly deleterious alleles can lead to large amounts of inbreeding depression because of their high equilibrium frequencies. Molecular studies correlating level of heterozygosity and degree of heterosis should take into account this nonuniform distribution of deleterious alleles in polyploid genomes. Preservation or enhancement of heterozygosity would be more critical at proximal regions than at other chromosome regions in polysomic polyploid species.
Polyploidy is widespread among angiosperms, corresponding to more than half of the total number of species and as much as 75% of the domesticated plant taxa (1, 2). Alfalfa, banana, canola, coffee, cotton, potato, soybean, strawberry, sugar cane, sweet potato, and wheat are all examples of polyploids of economic importance. Therefore, insights into the evolutionary consequences of polyploidy on genome organization would provide crucial information for designing sound breeding strategies for these crop species.
In nature, several mechanisms have been implicated in the origin of polyploid genomes (1). They can originate by spontaneous somatic chromosome doubling at apical chimeras or by occurrence of a zygote able to produce a polyploid individual. Another possible origin is through the mating of plants producing either 2n pollen grains or 2n eggs (1). Studies employing molecular techniques confirm that polyploidization is a dynamic as well as “reversible” process (3, 4). Both polyploidization of ancient diploid genomes and haploidization of ancestral polyploid genomes are now recognized as important phenomena in plant chromosome evolution (3–5).
Polyploids have been classified as either autopolyploids, originated from genetically similar genomes, or allopolyploids with chromosome combination derived from distinct (interspecific) genomes (3, 6). However, the terms disomic polyploids and polysomic polyploids better describe the nature of polyploids from the viewpoint of both genetics and breeding (7). More than two homologous chromosomes of polysomic polyploids can pair at meiosis, resulting in the formation of multivalents and polysomic inheritance (autopolyploids are often polysomic) (7). Disomic polyploids have bivalent chromosome pairing between strictly homologous chromosomes and a diploid-like genetic behavior (allopolyploids are generally disomic) (7). However, many polyploid taxa represent intermediate stages between these two extremes (3, 6).
Multivalent formation can result in double reduction. Double reduction arises from a combination of three major events during meiosis: crossing-over between non-sister chromatids, an appropriated pattern of disjunction, and the subsequent migration of the chromosomal segments carrying a pair of sister alleles to the same gamete (8, 9). Theoretically, the rate of double reduction, α (10), could assume maximum values of 0 (with pure random chromosome segregation), 1/7 (with pure random chromatid segregation), and 1/6 (with complete equational segregation) (11, 12). Experiments aimed at estimating α in tetraploids have yielded values ranging from 0% to almost 30% (13–17).
Several parameters are taken into account to determine the rate of double reduction at a locus (18, 19). The two most obvious are the proportion at which a chromosome enters in a multivalent configuration at meiosis and the crossing-over frequency between a locus and its centromere. Thus, double reduction is a position-dependent phenomenon. It may vary depending on which chromosome the locus resides on, because chromosomes may vary in their propensity to form multivalents, and where it resides on a chromosome, as the value of α will be greater toward the distal-proterminal regions and almost null at loci near the centromeres. These predictions have been proven experimentally (16).
The subchromosomal site of genetic loci is considered a possible factor in determining the extent and type of genetic change in polyploids (3). Wright (20) acknowledged that double reduction could affect the distribution of gene frequency in polyploid populations, but thought that this effect could be ignored. Fisher (13) also recognized that loci around the centromere are more protected against inbreeding, and Stebbins (21) hypothesized that recessive deleterious mutations should accumulate preferentially around the centromeres of ring-forming Onoethera chromosomes. In addition, the genetic nature of higher ploidy levels would allow for greater levels of heterozygosity as well as the persistence of deleterious alleles at a locus (4, 22). Under such circumstances, the occurrence of deleterious loci expressed postmeiotically and selected during the gametophytic stage of the life cycle cannot be ignored. A large number of genes is expressed in both sporophyte and gametophyte (23, 24) and it is likely that many of these genes are transcribed postmeiotically (24). However, to our knowledge, no model has been developed to estimate the impact of polysomic polyploidy on the accumulation of deleterious mutations and to assess the consequences of double reduction and gametophytic selection on the frequency and distribution of genetic loci along the chromosomes.
In the present study, we model the effects of double reduction and gametophytic selection on the equilibrium frequency of deleterious alleles in polysomic tetraploid populations. Polysomy has been considered a powerful mechanism for protection against inbreeding, especially when α = 0, such as in the neighborhood of the centromeres (13). For this reason, we also examined the consequences of selfing on the fitness of a polysomic tetraploid population in equilibrium at loci subjected to different levels of double reduction.
Mathematical Model
Establishing Generalized Recurrence Relations.
Consider a genetic locus with two alleles, A and a, subjected to a rate of double reduction α. The mutation rate, from A to a, is μ, and the reverse-mutation rate is ν. Fitnesses of sporophytes with genotypes AAAA, AAAa, AAaa, Aaaa, and aaaa are w0, w1, w2, w3, and w4, respectively. Fitnesses of female gametophytes with genotypes AA, Aa, and aa are ωf0, ωf1, and ωf2, respectively. Fitnesses of male gametophytes with genotypes AA, Aa, and aa are ωm0, ωm1, and ωm2, respectively. All fitnesses must be greater than or equal to 0. If the frequencies of AAAA, AAAa, AAaa, Aaaa, and aaaa formed at generation t are f0t, f1t, f2t, f3t, and f4t, respectively, then the frequencies of gamete-forming adults are F0t = w0f0t/w̄t, F1t = w1f1t/w̄t, F2t = w2f2t/w̄t, F3t = w3f3t/w̄t, and F4t = w4f4t/w̄t, respectively,
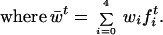 |
The frequencies of gametophytes of genotypes AA, Aa, and aa are, respectively,
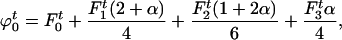 |
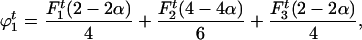 |
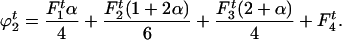 |
Mutation changes these values to
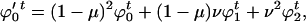 |
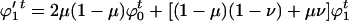 |
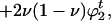 |
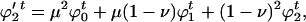 |
respectively.
The frequencies of female gametes of genotypes AA, Aa, and aa available for random mating are 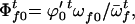
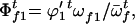 and
and 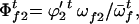 respectively,
respectively,
where
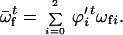 |
The frequencies of male gametes of genotypes AA, Aa, and aa available for random mating are 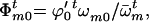
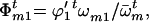 and
and 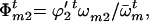 respectively,
respectively,
where
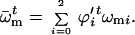 |
The frequencies of sporophytic genotypes AAAA, AAAa, AAaa, Aaaa, and aaaa formed by random mating in generation t + 1 are 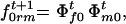
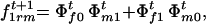
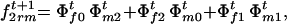
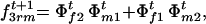 and
and 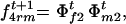 respectively.
respectively.
The random mating equations can be used to derive the frequencies of sporophytic genotypes formed by selfing: self-fertilizing a given parent (say AAaa) and random-mating a population of individuals with only that genotype are equivalent. Let fi,j be the frequency of sporophytes of genotype i produced by a parent of genotype j. Then fi,j = fi rm1 when using Fj0 = 1 and Fnot j0 = 0 to calculate fi rm1.
The frequencies of sporophytic genotypes AAAA, AAAa, AAaa, Aaaa, and aaaa formed by self-fertilization in generation t + 1 are of the form
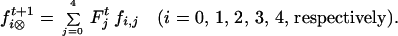 |
We normalized the frequencies before calculating the weighted average over parental genotypes. Posterior normalization may alternatively be used with no great differences in the final outcome. Whenever normalizing, proper care needs to be taken in the computations to avoid possible divisions by zero.
We finally use the rate of self-fertilization, σ, to calculate the frequencies of each sporophytic type formed to constitute generation t + 1:
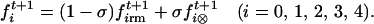 |
This procedure is iterated until equilibrium is reached. The equilibrium frequency of allele a is then
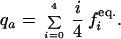 |
Closed form equations can be obtained by rearranging equations that provide equilibrium frequencies of gametophytes or sporophytes, as functions of α, σ, and qa (19, 25). In this case, the a allele has to be completely recessive and deleterious to either the sporophyte (w4 = 1 − s) or the gametophyte (ωf2 = ωm2 = 1 − g). We would then use the approximations
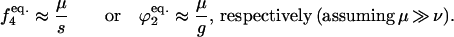 |
However, closed form equations deduced this way provide accurate answers only for a small subset of the parameter space (results not shown).
Modeling the Effect of Inbreeding.
Assume a population for which genotypic frequencies(fi0s), sporophytic and gametophytic fitnesses (wis, ωfis, and ωmis), mutation and double reduction rates (μ, ν, and α) are all defined for a given locus. We are interested in the change of fitness of that population for the first few generations of selfing. We assumed that each parent is able to contribute exactly one selfed progeny to form the next generation. This is a simplifying assumption, but it mimics how many inbreeding experiments are conducted. We calculated the fitness of generation t relative to generation 0 as
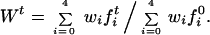 |
A similar approach is used to calculate change in gametophytic viability, but in this case allelic frequencies are allowed to change because of intraplant gametic selection. This again mimics how inbreeding experiments are done: the experimenter is seldom able to prevent gametophytic selection. The relative fertility of generation t relative to generation 0 is
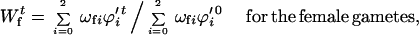 |
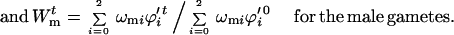 |
In the present model we will assume no reverse mutation and consider only completely recessive deleterious mutations that equally affect the male and female gametophytes. That is: ν = 0; [w0 = w1 = w2 = w3 = 1]; w4 = 1 − s, [ωf0 = ωf1 = ωm0 = ωm1 = 1], and [ωf2 = ωm2 = 1 − g].
For both the equilibrium frequencies and the inbreeding depression aspects of this analysis, we focused our attention on three values of α: α = 0 (absence of double reduction), α = 0.04 (corresponding to a locus where one would detect 1% of duplex genotype in the progeny of a simplex by nulliplex cross), and α = 1/7 (pure random chromatid segregation model). Computations were done in a Microsoft Excel97 workbook using Visual Basic for Applications to implement the recurrence equations and observe the effect of selfing.
Results
General Effects of Double Reduction.
A low rate of double reduction is enough to reduce greatly the equilibrium frequency of a deleterious allele at a locus (qa). In the absence of double reduction, qa is approximately 3.2% for a lethal allele generated at a mutation rate of 10−6. This value drops to 1.4% when α equals 0.04 (Fig. 1). The frequency of individuals carrying at least one copy of allele a is 12.1% in the first case, but declines to 5.3% if α = 0.04. If this allele causes a 10% reduction in fitness, then, at equilibrium, 21% of the individuals would carry at least one copy of allele a when α = 0, compared with 13% when α = 0.04. The impact of double reduction is even more pronounced when deleterious alleles at loci expressed in the gametophyte are considered. There, only minute levels of double reduction are needed to reduce equilibrium frequencies severalfold (Fig. 2).
Figure 1.

Equilibrium frequencies, qa, of the deleterious allele a in the absence of selfing or gametophytic selection. The frequency of double reduction is α, the mutation rate is μ, and the fitness of the aaaa genotype is 1 − s. For a diploid species under similar selection and mutation pressures the equilibrium allele frequencies are less than 0.35%.
Figure 2.

Equilibrium frequencies, qa, of a gametophytic lethal allele in the absence of selfing. The range of double reduction values, α, shown is only from 0 to 0.01, at which point most of the reduction in frequency has already taken place. The equilibrium frequencies for α = 0.17 are 5 × 10−5 and 5 × 10−6 when the mutation rates, μ, are 10−5 and 10−6, respectively.
From these observations, we expect that a slight pleiotropic effect of a sporophytic lethal allele on the viability of the gametophyte would greatly accentuate the difference in equilibrium frequency between loci subjected to different levels of double reduction. For example, with a pleiotropic reduction of gametic viability of only 1% [setting (μ, σ, 1 − s, 1 − g) = (10−6, 0, 0, 0.99)], the equilibrium frequency of a sporophytic lethal allele will decrease by half (from 1% to 0.5%) when α changes from 0 to 0.01.
There is a decrease in the equilibrium frequencies of deleterious alleles in the population if some selfing occurs, but the effect of double reduction remains significant for moderate amounts of selfing (Fig. 3 A and B). Furthermore, there are circumstances when some selfing can actually exacerbate the relative difference in frequencies between chromosomal regions with and without double reduction, even if the absolute frequencies of the deleterious alleles are reduced. This is the case, for example, when allele a is a sporophytic recessive lethal and approximately 5% of selfing occurs in the population (Fig. 3A).
Figure 3.

Ratio of equilibrium frequencies of the deleterious allele (qa) at a locus not subjected to double reduction compared with a locus subjected to a double reduction of 0.04 and of 0.17, upon different amounts of selfing. A applies in the absence of gametophytic selection (1 − g = 1), and B applies when the aa genotype is lethal to the gametophyte (1 − g = 0). For all cases we chose a mutation rate, μ, of 10−6 and lethality of the aaaa sporophytic genotype (1 − s = 0).
Impact of Selfing a Random-Mating Population in Equilibrium.
We considered how selfing would affect the fitness of an outcrossing polysomic tetraploid population in equilibrium. We found that inbreeding depression becomes more severe in polyploids than in diploids after the second generation of selfing (or third generation for higher values of α) when a sporophytic lethal gene is considered. Such depression is accentuated when loci reside in regions with no double reduction (Fig. 4). The effect of a gametophytic lethal on the viability of the gametic pool after selfing was also modeled. When α = 0, a substantial decline in viability is observed for gametes produced by plants resulting from one or two generations of selfing. However, a gradual recovery began by the third generation of selfing (Fig. 5). Small levels of double reduction were able to reduce greatly the equilibrium frequencies of gametophytic lethal alleles (Fig. 2). Therefore, it is not a surprise that little reduction of viability is observed when α = 0.04 (Fig. 5).
Figure 4.

Change of fitness during the first three generations of selfing diploid and tetraploid populations in equilibrium (assuming a mutation rate of 10−6, random mating, and a deleterious allele lethal to homozygous sporophytes and double reduction rates α = 0, 0.04, and 0.17). The inbreeding coefficient Fi is obtained from the recurrence relations Fi = (1 + Fi−1)/2 and Fi = (1 + 2α + (5 − 2α)Fi−1)/6 for the diploid and the tetraploid populations, respectively (where i represents the number of generations of selfing and α the is rate of double reduction) (51). At generation 0 we stipulate an inbreeding coefficient of 0 and a fitness of 1.
Figure 5.

Change of gametophytic viability under selfing of tetraploid populations in equilibrium (assuming a mutation rate of 10−6, random mating, and a deleterious allele lethal to homozygous sporophytes or gametophytes). The value of 1 is given to the gametophytic viability of generation 0.
Discussion
Effects of Double Reduction.
The genome of polysomic polyploid species is heterogeneous in its ability to eliminate deleterious alleles, even at low levels of double reduction. This phenomenon appears more pronounced for alleles of genes expressed in gametophytes. The rate of double reduction varies along the chromosome arm, being almost nil in the neighborhood of the centromere, and increasing toward the telomeric ends. Therefore, a corresponding decrease in the frequency of deleterious alleles should occur at distal loci in polysomic polyploid organisms. As double reduction is contingent on multivalent formation, this position effect should be less pronounced for chromosomes that form mostly bivalents. Furthermore, the polysomic chromosomes are likely to contribute more to inbreeding depression than do the disomic chromosomes in a species that presents a mixture of pairing types.
Effect of Gametophytic Selection.
The possibility of deleterious alleles at loci that influence the fitness of both the sporophyte and the gametophyte is often ignored because such alleles are extremely rare in diploid genomes. Deleterious alleles at such loci can accumulate at much greater frequencies in the case of polysomic polyploids. Furthermore, equilibrium frequencies at these genes are most affected by changes in α.
The effect of gametophytic selection may not be restricted to polysomic polyploids. Extensive genetic redundancy is known to occur in ancient polyploids such as maize (26). Such redundancy could be advantageous by allowing the evolution of novel functions at one locus of a duplicated gene. However, in the complete absence of selection pressure, a sporophytic gene in a finite population may be converted too rapidly into a pseudogene without a chance to evolve novel functions (27). Takahata and Maruyama (28) found that the time for silencing an unlinked duplicate locus (under a double recessive fitness model) increases with effective population size, and that partial dominance greatly increased the time to fix a null allele. Genes under gametophytic selection are under the same effective population size as genes solely under sporophytic selection. However, there cannot be alleles that are completely recessive “de facto” at these loci: gametophytic selection imparts a form of partial dominance to any allele. Thus, redundancy should be higher at genes under gametophytic selection, and these genes may have a greater chance to evolve new functions.
Inbreeding in Polysomic Tetraploids.
There is a linear relationship between mean performance and decrease in heterozygosity in diploids, unless there are significant effects from linkage or epistasis (29). This linear relationship, however, is not typical of polysomic polyploids (30–32). In fact, from a theoretical standpoint, there is no reason why there should be a linear relationship in polysomic polyploids (33, 34). The inbreeding coefficient (F) is the probability that two alleles taken at random from an individual are identical by descent. This is equivalent in diploids to the probability of fixation of either allele. However, in tetraploids the probabilities of loosing one, two, or three of the original alleles are confounded in the F coefficient:
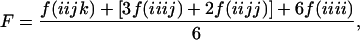 |
where f(iiii) is the frequency of zygotes with four alleles identical by descent.
When considering a population in selection–mutation balance, the probability of loss of three alleles [f(iiii)] is the main determinant of inbreeding depression because the deleterious allele is predominantly carried by AAAa individuals. Therefore, three A alleles need to be lost before the recessive phenotype is expressed.
Inbreeding depression is often greater in polysomic polyploids than in diploids at the same inbreeding coefficient (35). In the absence of double reduction, in a random mating n-ploid population, if we approximate the equilibrium frequency of a recessive allele deleterious to the sporophyte by
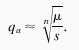 |
and we define the fitness observed upon inbreeding to complete homozygosity by
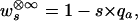 |
then the same ultimate amount of inbreeding depression in populations of different ploidy levels (say m- and n-ploid) will be caused by alleles whose deleterious effects (sm and sn) can be related by
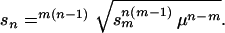 |
For example, with μ = 10−6, an allele that reduces fitness by only 1% percent (s = 0.01) in a tetraploid population will cause the same ultimate inbreeding depression as a lethal allele (s = 1) in a diploid population. Thus, alleles with only a very mild deleterious effect can be responsible for large amounts of inbreeding depression in polyploids because of their high equilibrium frequencies.
Our model predicts little inbreeding depression for the first two generations of selfing, contrary to what is empirically observed (35). Thus, linkage or allelic interactions must play a role. In polyploids, inbreeding depression is often analyzed in terms of the loss of higher-level allelic interactions at tetragenic loci (32, 33). Given the high genetic load expected, it seems that linkage should be given a more prominent role and “interacting alleles” could well be linkage blocks or “linkats” of loci with deleterious alleles in repulsion phase linkage (36, 37). This argument is supported by the fact that it has already been shown to be the case in diploids (38, 39), for which deleterious alleles are expected to accumulate at much smaller frequencies.
Implications for Plant Breeding and Molecular Genetics.
Proximal loci should have a much greater impact at reducing the fitness of a polyploid population upon inbreeding. Therefore, the preservation of proximal heterozygosity would be more critical than the preservation of heterozygosity at central and distal loci when breeding polysomic polyploids. Meiotic mutants in potato were used to identify genomic regions between centromeres and proximal chiasmata as the sites of major loci for tuber yield in potato (40–42). These regions may require higher levels of heterozygosity to offset the higher frequency of deleterious recessive alleles.
Pollen selection has been proposed for use in the breeding of diploid species (43). Moderately inbred polyploids may be an excellent system to demonstrate the effect of pollen selection on subsequent sporophytic generations. Selection pressure is usually less intense on the female than on the male gametophyte (44). Thus, reciprocal crosses between parents at different levels of inbreeding could lead to progenies with significantly different performances.
Molecular marker analyses of polysomic polyploid populations may also be affected. Analyses attempting to relate molecular marker heterozygosity to heterosis should detect subsets of loci that tend to give a greater correlation than the complete set, even when considering a range of unrelated germ plasm (45). Such observation would reflect the higher frequency of deleterious alleles present in proximal regions and probably not the presence of some genes of major effect (42). In quantitative trait loci (QTL) mapping experiments, dominant and overdominant QTL should be detected more often in genomic regions where a higher frequency of deleterious alleles is expected, irrespective of the trait being mapped. In fact, there seem to be extensive overlap of potato QTL (46–48). Thus, one should be cautious before making an inference about pleiotropy. Furthermore, variation at a much greater number of loci should affect quantitative traits in a polyploid species because of the relaxed selection pressure on genes expressed in the gametophyte. This added variability may reduce greatly the value of QTL mapping in diploid relatives and claiming orthology of QTL from different ploidy levels. When using mapping populations derived from crosses between wild diploid species and haploidized tetraploid parents, most of the dominant QTL (irrespective of the direction of their effect) should come from the wild diploid, those being functional dominant alleles that will compensate for recessive deleterious alleles present in the haploidized tetraploid parent (46, 47).
The model presented here is a very simple one. More realistic models could point to forces that counteract this heterogeneity or greatly reduce the impact of double reduction. The most obvious one is random genetic drift (49). What minimum population size is needed to maintain a meaningful difference in deleterious allele frequencies between genomic regions with and without double reduction? Multilocus models also need to be developed to assess the effect of linkage, and results should be generalized to any ploidy level. There is a great need for theoretical studies to account for some of these peculiarities of plant evolution (50), while also providing plant geneticists with some null hypotheses and models to test, especially at a time when there is a growing interest in the genetics and breeding of polyploids (3).
Acknowledgments
We are grateful to Drs. E. T. Bingham, J. G. Coors, J. F. Crow, R. P. Guries, J. L. Kermicle, D. M. Lascoux, T. C. Osborn, P. W. Simon, and S. J. Peloquin for their comments and/or critical review of the manuscript. We especially thank Dr. C. Denniston for his help in generalizing the mathematical model. This paper was written while D.V.B. received research assistantship support from the U.S. Department of Agriculture (in the form of Grant USDA-NRICGP#9701693 to J. G. Coors).
Abbreviation
- QTL
quantitative trait loci
Footnotes
Article published online before print: Proc. Natl. Acad. Sci. USA, 10.1073/pnas.100101097.
Article and publication date are at www.pnas.org/cgi/doi/10.1073/pnas.100101097
References
- 1.Ramsey J, Schemske D W. Annu Rev Ecol Syst. 1998;29:467–501. [Google Scholar]
- 2.Hilu K W. Am J Bot. 1993;80:1494–1499. [Google Scholar]
- 3.Leitch I J, Bennett M D. Trends Plant Sci. 1997;2:470–476. [Google Scholar]
- 4.Soltis D E, Soltis P S. Proc Natl Acad Sci USA. 1995;92:8089–8091. doi: 10.1073/pnas.92.18.8089. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Bretagnolle F, Felber F, Calame F G, Küpfer P. Bot Helv. 1998;108:5–37. [Google Scholar]
- 6.Jackson R C, Jackson J W. Am J Bot. 1996;83:673–678. [Google Scholar]
- 7.Peloquin S J. In: Plant Breeding II. Frey K J, editor. Ames: Iowa Sate Univ. Press; 1981. pp. 117–150. [Google Scholar]
- 8.Darlington C D. J Genet. 1929;21:207–218. [Google Scholar]
- 9.Haldane J B S. J Genet. 1930;22:357–372. [Google Scholar]
- 10.Fisher R A, Mather K. Ann Eugen. 1943;12:1–23. [Google Scholar]
- 11.Muller H J. Am Nat. 1914;48:508–512. [Google Scholar]
- 12.Mather K. J Genet. 1935;30:53–78. [Google Scholar]
- 13.Fisher R A. The Theory of Inbreeding. New York: Hafner; 1949. [Google Scholar]
- 14.Catcheside D G. Heredity. 1956;10:205–218. [Google Scholar]
- 15.Catcheside D G. Heredity. 1959;13:403–407. [Google Scholar]
- 16.Welch J E. Genetics. 1962;47:367–396. doi: 10.1093/genetics/47.4.367. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Haynes K G, Douches D S. Theor Appl Genet. 1993;85:857–862. doi: 10.1007/BF00225029. [DOI] [PubMed] [Google Scholar]
- 18.Mather K. J Genet. 1936;32:287–314. [Google Scholar]
- 19.Demarly Y. Ann Amélior Plantes. 1963;13:307–400. [Google Scholar]
- 20.Wright S. Proc Natl Acad Sci USA. 1938;24:372–377. doi: 10.1073/pnas.24.9.372. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Stebbins G L. Chromosomal Evolution in Higher Plants. Reading, MA: Addison-Wesley; 1971. [Google Scholar]
- 22.Soltis D, Rieseberg L H. Am J Bot. 1986;73:310–318. [Google Scholar]
- 23.Simon P W, Peloquin S J. J Hered. 1976;67:204–208. [Google Scholar]
- 24.Ottaviano E, Mulcahy D L. Adv Genet. 1989;26:1–64. [Google Scholar]
- 25.Bennett J H. Biometrics. 1968;24:485–500. [PubMed] [Google Scholar]
- 26.Gaut B S, Doebley J F. Proc Natl Acad Sci USA. 1997;94:6809–6814. doi: 10.1073/pnas.94.13.6809. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Walsh J B. Genetics. 1995;139:421–428. doi: 10.1093/genetics/139.1.421. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Takahata N, Maruyama T. Proc Natl Acad Sci USA. 1979;76:4521–4525. doi: 10.1073/pnas.76.9.4521. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Wright S. USDA Bull. 1922;1121:1–60. [Google Scholar]
- 30.Dewey D R. Crop Sci. 1966;6:144–147. [Google Scholar]
- 31.Dewey D R. Crop Sci. 1969;9:592–595. [Google Scholar]
- 32.Rice J S, Dudley J W. Crop Sci. 1974;14:390–393. [Google Scholar]
- 33.Bennett J H. Biometrics. 1976;32:449–452. [PubMed] [Google Scholar]
- 34.Busbice T H, Wilsie C P. Euphytica. 1966;15:52–67. [Google Scholar]
- 35.Jones J S, Bingham E T. Plant Breed Rev. 1995;13:209–233. [Google Scholar]
- 36.Demarly Y. In: Proceedings of the Conference on Broadening the Genetic Base of Crops. Zeven A C, van Harten A M, editors. Wageningen, The Netherlands: Center for Agricultural Publishing and Documentation; 1979. pp. 257–265. [Google Scholar]
- 37.Bingham E T, Groose R W, Woodfield D R, Kidwell K K. Crop Sci. 1994;34:823–829. [Google Scholar]
- 38.Moll R H, Lindsey M F, Robinson H F. Genetics. 1964;49:411–423. doi: 10.1093/genetics/49.3.411. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Gardner C O, Lonnquist J H. Agron J. 1959;51:524–528. [Google Scholar]
- 40.Buso J A. Ph.D. thesis. Madison: Univ. of Wisconsin; 1986. [Google Scholar]
- 41.Tai G C C, De Jong H. Theor Appl Genet. 1997;94:303–308. [Google Scholar]
- 42.Buso J A, Boiteux L S, Tai G C C, Peloquin S J. Proc Natl Acad Sci USA. 1999;96:1773–1778. doi: 10.1073/pnas.96.4.1773. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Sarli-Gorla M, Frova C. In: Pollen Biotechnology for Crop Production and Improvement. Stephenson A G, Kao T-h, editors. Rockville, MD: Am. Soc. Plant Physiol.; 1997. pp. 230–239. [Google Scholar]
- 44.Khush G S, Rick C M. Genetica. 1967;38:74–94. [Google Scholar]
- 45.Bonierbale M L, Plaisted R L, Tanksley S D. Theor Appl Genet. 1993;86:481–491. doi: 10.1007/BF00838564. [DOI] [PubMed] [Google Scholar]
- 46.Van den Berg J H, Ewing E E, Plaisted R L, McMurry S, Bonierbale M W. Theor Appl Genet. 1996;93:307–316. doi: 10.1007/BF00223170. [DOI] [PubMed] [Google Scholar]
- 47.Van den Berg J H, Ewing E E, Plaisted R L, McMurry S, Bonierbale M W. Theor Appl Genet. 1996;93:317–324. doi: 10.1007/BF00223171. [DOI] [PubMed] [Google Scholar]
- 48.Schäfer-Pregl R, Ritter E, Concilio L, Hesselbach J, Lovatti L, Walkemeier B, Thelen H, Salamini F, Gebhardt C. Theor Appl Genet. 1998;97:834–846. [Google Scholar]
- 49.Moody M E, Mueller L D, Soltis D E. Genetics. 1993;134:649–657. doi: 10.1093/genetics/134.2.649. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Bever J D, Felber F. Oxford Surv Evol Biol. 1992;8:185–217. [Google Scholar]
- 51.Kempthorne O. An Introduction to Genetic Statistics. New York: Wiley; 1957. [Google Scholar]