
Figure 7. A model for the tissue-specific regulation of arg-1 expression by CeTwist.
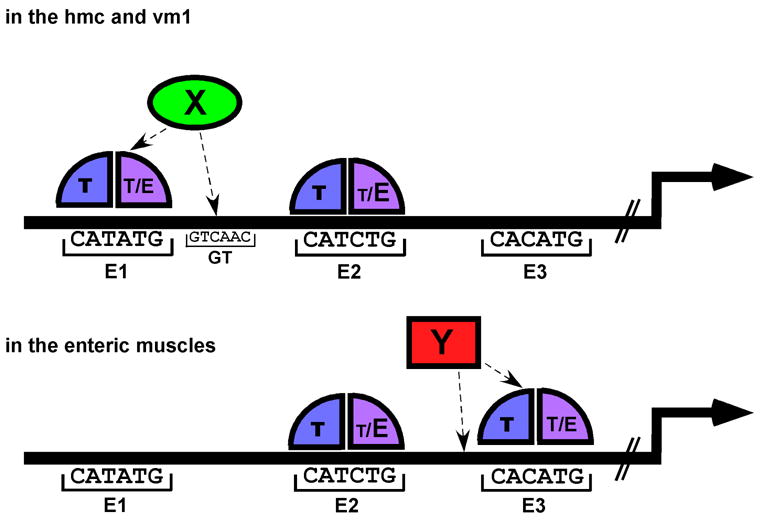
Each of the three E boxes in the 385 bp minimal promoter region could be bound by either CeTwist/CeE/DA heterodimers (T/E) or CeTwist homodimers (T/T). E2 is required for arg-1 expression in all of the tissues and is bound by T/E with high affinity in vitro (shown by the larger E vs. the smaller T bound at E2). In contrast, E1 and E3 are bound by T/E or T/T with lower affinity in vitro. The expression of arg-1 in the hmc and vm1 requires T/E or T/T to bind to E1 and to E2. It is proposed in this model that an ancillary factor (X) is present in the hmc and vm1 to facilitate the binding of CeTwist dimers to E1, allowing arg-1 expression to occur in these tissues. Similarly, in the enteric muscles, CeTwist dimers bind to both E2 and E3. A proposed ancillary factor (Y) in the enteric muscles may increase the binding affinity of CeTwist dimers to E3. Factors X and Y would be present in a subset of the tissues, thereby conferring tissue-specific regulation of arg-1 expression through the E boxes. An alternative possibility (not shown) could exist where T/E or T/T predominate in the individual tissues and have more affinity for either E1 or E3 whereas E2 can bind either dimer. In such a model, the presence of the specific CeTwist containing dimer would lead to tissue-specific regulation through the E boxes.