

Abstract
Heterochrony has become a central organizing concept relating development and evolution. Unfortunately, the standard definition of heterochrony—evolutionary change in the rate or timing of developmental processes—is so broad as to apply to any case of phenotypic evolution. Conversely, the standard classes of heterochrony only accurately describe a small subset of the possible ways that ontogeny can change. I demonstrate here that the nomenclature of heterochrony is meaningful only when there is a uniform change in the rate or timing of some ontogenetic process, with no change in the internal structure of that process. Given two ontogenetic trajectories, we can test for this restricted definition of heterochrony by asking if a uniform stretching or translation of one trajectory along the time axis superimposes it on the other trajectory. If so, then the trajectories are related by a uniform change in the rate or timing of development. If not, then there has been change within the ontogenetic process under study. I apply this technique to published data on fossil Echinoids and to the comparison of human and chimpanzee growth curves. For the Echinoids, some characters do show heterochrony (hypermorphosis), while others, which had previously been seen as examples of heterochrony, fail the test—implying that their evolution involved changes in the process of development, not just the rate at which it proceeded. Analysis of human and chimpanzee growth curves indicates a combination of neoteny and sequential hypermorphosis, two processes previously seen as alternate explanations for the differences between these species.
Keywords: evolution of development, morphology
A fundamental problem facing systematists and comparative biologists is that of deciding just how different two separate phenotypes are. While a number of morphometric techniques exist for quantifying the differences between phenotypes, these techniques in general do not address the issue of greatest concern: namely, what genetic or developmental changes must take place to derive a particular descendant phenotype from its ancestor, and how readily do these changes occur in the evolution of a lineage? Devising any general theory to address these questions will require a formalism for comparing developmental processes and describing the relations between them. One promising candidate for such a formalism has been the idea of heterochrony.
This idea holds the promise of allowing us to map differences in growth trajectories, which can be measured with relative ease for extant and, sometimes, even extinct organisms, to differences in developmental processes. Any such extrapolation from pattern to process, however, requires great care. I argue below that if the concept of heterochrony is to be up to the task, we must narrow and clarify its definition, lest different processes be lumped together because we erroneously define their resulting patterns as similar.
Heterochrony is generally defined as a change in the relative timing or rate of development of a character that persists from ancestor to descendant (1–3). Most authors now recognize six forms of “pure” heterochrony [progenesis, hypermorphosis, neoteny, acceleration, and pre- and post-displacement (4)], which are presumed to correspond to different kinds of evolutionary transformations in development. Although the term was originally applied at the level of the whole organism, it is now generally applied to any developmental process [often assuming it represents a causal sequence (5)], and different characters within an organism can exhibit different types of heterochrony. Using this expanded definition, heterochrony can be invoked even in the origin of morphological novelties through the rearrangement of inductive interactions (6).
This expanded definition, however, produces a problem: if heterochrony is defined as a change in the rate or timing of any developmental event, then it is hard to imagine an evolutionary change in phenotype that could not be said to result from some form of heterochrony. This problem has been noted by a number of authors, some of whom see it as a potential drawback, resulting from the fact that even changes that do not directly involve timing can have consequences that look like heterochrony (7, 8). Others, however, see it as implying that there is really no other way to change an existing structure (3).
In this paper, I argue that the traditional nomenclature of heterochrony, with its discrete categories, applies to only a narrow set of cases. Below, I define these cases and show how to identify them by comparing ontogenetic trajectories. Ultimately, narrowing the definition of heterochrony will allow us to better assess the biological processes behind both cases that do and cases that do not represent some type of heterochrony.
Ontogenetic Trajectories and the Kinds of Heterochrony
De Beer (9) based his discussion of heterochrony on the presence or absence of discrete characters in the juvenile and adult stages of descendants relative to their ancestors. This naturally leads to the identification of a finite number of distinct types of heterochrony. Gould (1) continued this tradition by sorting through De Beer’s list, discarding some types and clarifying others. He also changed the nature of the discussion by defining heterochrony in terms of continuous variables, namely size and shape.
Gould’s “clock” model defines the different types of heterochrony by specifying the values of size and shape at a standardized stage of development in a descendant relative to its ancestor. Alberch et al. (4) took the next step of allowing size and shape to change continuously by representing ontogeny as a path through phenotype-time space. This path, the ontogenetic trajectory, explicitly treats development as a continuous time dynamic process. These authors were principally concerned with demonstrating that the various types of heterochrony could be represented and studied in terms of ontogenetic trajectories. While this is clearly true, the oft-assumed corollary, that any change in an ontogenetic trajectory can be accurately described as some combination of six “pure” types of heterochrony, is not. I shall demonstrate below that the nomenclature of heterochrony applies only to a small (though potentially important) and well defined set of possible changes in an ontogenetic trajectory.
Alberch et al. (4) noted that the study of an ontogenetic trajectory could be simplified by linearizing the trajectory. Once linearized, a trajectory is defined completely by its endpoints, and the various types of heterochrony can be defined as different ways of shifting these endpoints. More recently, some authors (3) have dropped the convention of linearizing the trajectory and focused on changes in the origin and endpoints of whatever curve best represents the growth function being considered. The operative term here is the growth function. Because the various types of heterochrony are defined as different ways of shifting the endpoints of an ontogenetic trajectory, they cannot completely describe an evolutionary change in ontogeny unless the same type of curve connects those endpoints. I illustrate below what it means to say that two curves are of the “same type,” and discuss what restrictions this puts on our use of heterochronic terms.
I first focus on the simple kinds of growth functions considered by Alberch et al., those described by first order, autonomous differential equations. For such a system, one can indeed always find a transformation of the phenotype axis that linearizes any particular trajectory. The hitch is that this is not always the same transformation for any two trajectories.
As an example, consider a hypothetical organism (or organ) that grows according to the logistic equation
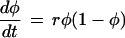 |
1 |
(Fig. 1; φ can represent any phenotypic character; I will use body size in the example). Here, growth rate drops off linearly as the overall size approaches some limit. The two curves in Fig. 1a result from different values of the growth rate parameter, r. Fig. 1b shows the results of applying the same linearizing transformation to the curves in Fig. 1a. Here, the difference between the two curves can be seen clearly as a difference in the slopes of the linearized functions. This illustrates that this particular change is described completely by a shift in one of the endpoints of the linearized ancestral trajectory.
Figure 1.

Ontogenetic trajectories of a hypothetical ancestor and its descendant. Both curves are defined by the logistic equation with different values of r (see text). (a) Untransformed trajectories. (b) Both trajectories after transforming the size data using the logistic linearizing transform (Eq. A6). The transformation separates the endpoints of the trajectories, which are so close as to look identical in the untransformed case.
In contrast, Fig.2 shows two growth trajectories that, although similar, cannot be linearized by the same transformation. This is illustrated in Fig. 2b, which shows that the transformation of the phenotype axis that linearizes the ancestral trajectory puts a distinct bend into the descendant trajectory. What this might mean biologically is suggested by a comparison of the functions that produced the two curves. The ancestor in Fig. 2 is the standard logistic curve (Eq. 1). The descendant trajectory is modified so that the feedback term is nonlinear:
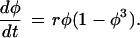 |
2 |
The difference between descendant and ancestor therefore involves not just a change in the rate of growth (though that is a consequence) but also a change in the rules that translate overall size into growth rate.
Figure 2.

(a) An ancestral trajectory (solid line) showing logistic growth (Eq. 1) and a descendant (dashed line) resulting from a modified growth equation (Eq. 2). (b) The same two trajectories transformed according to Eq. A6. Note that the transformed trajectory of the descendant is not a straight line.
The example in Fig. 1 represents a change that can be described completely using the language of heterochrony, this case being an example of acceleration. Fig. 2, however, shows a change in ontogeny that involves more than just a change in the endpoints of the ancestral trajectory. In fact, as we see from Eq. 2, it involves a change in the very structure of the function generating the trajectory. That there is a concomitant change in rate is a consequence of the fact that any change in the outcome of a dynamical system must involve a change in the rate at which something happens. Defining a change such as that in Fig. 2 as heterochrony is tantamount to making that term synonymous with the term morphological evolution. In contrast, I argue that the change shown in Fig. 1 is an example of a biologically meaningful subset of phenotypic changes that is deserving of a name and is accurately described by the nomenclature of heterochrony.
Defining Heterochrony
So, what exactly are the conditions under which the nomenclature of heterochrony is useful? As indicated in the examples above, and by the fact that the terms are derived from the comparison of linear trajectories, one sufficient condition is that the two ontogenetic trajectories are linearized by the same transformation of the phenotype axis. The Appendix shows that this is tantamount to saying that the time derivative of one function is a constant multiple of the time derivative of the other. Heterochrony is thus a uniform change in the rate or timing of some ontogenetic process, with no change in the nature of the biological interactions going on within that process.
Biologically, this definition means that we are considering only changes that speed up, slow down, or shift some developmental process as a unit. Mathematically, it means that given two trajectories, φ1(t) and φ2(t), there is a constant, C, such that for every value of the character φ* that is visited by both trajectories,
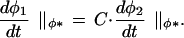 |
3 |
Though this discussion focuses on one-dimensional systems, the main point—the difference between a uniform scale change and a change in the form of the growth function—is just as relevant to multivariate comparisons (8), especially if results are grouped into the traditional classes of heterochrony (10).
This definition is much narrower than simply “any change in timing,” but this narrowness is what makes it useful. Using this definition, to say that a particular character transformation is an example of “acceleration” or “neoteny” is to say something about how the entire growth process of that character in the descendant is related to that of the ancestor, not just how the endpoints changed.
Of course, we could apply these terms in only a small portion of cases. Rather than being a drawback, though, this leads to another advantage of the narrow definition: saying that something is a case of heterochrony under this definition naturally leads one to look for particular kinds of biological processes, namely those that alter the ontogeny of the character as a unit. Alternately, failure to satisfy this narrow definition of heterochrony suggests that there has been change within the ontogenetic process that we are studying.
Just saying that something is a special case is not the same as labeling it unimportant, and there is good evidence that developmental processes exist that can yield the sorts of changes that fit this definition of heterochrony. These include the effects of growth hormone-like compounds (11), the lin- mutants in Caenorhabditis elegans (12), and studies of transgenic mice (13, 14).
To see how narrowing the definition of heterochrony leads us to interpret data differently, consider the allometric trajectories for Eocene Echinoids shown in Fig. 3 (redrawn from ref. 15). Here, two shape parameters are plotted against a measure of body size. The solid lines are the best fit polynomials through the data (not plotted), and the dashed lines indicate the rough outlines of the clouds of data points. Note that we cannot strictly ascertain heterochrony in these examples because shape is plotted with respect to body size rather than to time. We can still apply the same reasoning, though, so long as we are careful to specify that we are studying the relationship between two characters rather than one character and time.
Figure 3.

Allometric trajectories for three species of Eocene Echinoids. Redrawn from McKinney (15), who plots the data points and calculates the best fit polynomial curves through them. Dashed lines show the rough outlines of the actual clouds of data points. Periproct-margin distance is the distance between the anus and the margin of the test. The peristome comprises the mouth and surrounding structures.
In Fig. 3a, the trajectories of Oligopygus phelani and Oligopygus wetherbyi definitely seem to lie along the same line, which is nearly linear over values of the phenotype for which they overlap with Oligopygus haldemani. The trajectory for O. haldemani, though, is clearly not linear over that region. Using classical heterochronic nomenclature, we would interpret this as illustrating that, with respect to this character, O. wetherbyi is hypermorphic with respect to O. phelani and O. haldemani is neotenic and post-displaced with respect to the others (15).
Using the definition of heterochrony presented here, we still say that O. wetherbyi is hypermorphic to O. phelani since the same growth function that produces the latter can produce the former if it is simply extended. O. haldemani, however, shows a different kind of growth function altogether. Calling this neoteny would imply that the growth process is uniformly slowed down, when in fact there must have been some change in the way that periproct-margin distance changed with respect to body size, not just a change in the rate at which it did so. One transition could be achieved by a change that influenced the shape character and body size as a unit; the other must have influenced the two characters differentially.
Fig. 3b shows trajectories for a different character in the same three species. Once again, one of the species follows a trajectory that cannot be linearized along with that of the other two. In this case, though, it is O. phelani that exhibits the different pattern of growth. Though neither of these trajectories is completely linear, we can tell that they could not both be linearized by the fact that over the range in which they overlap, one is convex and the other is concave. Not all trajectories are so easily compared, though, so we seek an easier way to identify cases that meet this definition of heterochrony.
Comparing Trajectories
Linearizing trajectories is difficult, and is not even possible for nonmonotonic curves. Fortunately, there is an easier and more general test: If two trajectories, φ1(t) and φ2(t), differ by a uniform rate change, as defined by Eq. 3, then there exists some constant, z, such that for each value of φ visited by both trajectories, either φ2(t) = φ1(t + z) or φ2(t) = φ1(zt). This is tantamount to saying that we can superimpose one trajectory on the other simply by shifting it along the time axis or by multiplying each value on the time axis by a constant so as to linearly stretch it (Fig. 4 a–c). Note that trajectories that represent hypermorphosis or progenesis are already superimposed (though hypermorphosis is a problematic concept when dealing with nonlinear trajectories since it is not clear where a trajectory would go if allowed to continue). If we can simultaneously linearize two trajectories, then they necessarily also meet this more general criterion.
Figure 4.

Comparing trajectories. The examples in a, b, and c correspond to the six traditional types of heterochrony. (a) Translation along the time axis yields post- and pre-displacement. (b) Stretching the time axis yields neoteny and acceleration. (c) Hypermorphosis and progenesis. (d) Both the time and the phenotype axes are stretched (to the same degree). This would result if the trajectory were made up of many small linear segments, all of which underwent progenesis or hypermorphosis.
Defining heterochrony in this way leads us to pay extra attention to the level of organization at which characters are studied. For example, consider a character, φ, composed of two parts, ϕ1 and ϕ2 (for example, a forelimb composed of a humerus and a radius/ulna complex). For simplicity, let each component grow exponentially with different growth rates (r1 and r2). Then
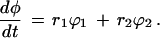 |
4 |
If either r1 or r2 changes by itself, or if they change to different degrees, then the overall change in φ will not meet the criteria discussed above, and the change would not be called heterochrony at the level of φ. Carrying out the same analysis with the two parts, however, would identify one (or both) of them as undergoing some form of heterochrony.
Clearly, there are kinds of transformations that are not adequately described with the language of heterochrony alone. This does not mean, however, that clarifying the meaning of the traditional terms removes these other kinds of evolutionary changes from consideration. In fact, it invites us to study them more carefully.
Fig. 4d shows a kind of transformation that is not included in the traditional nomenclature of heterochrony. It is clearly related to the other kinds of changes, but it does not represent a single uniform change in rate or timing. For a trajectory composed of many small linear segments, the transformation in Fig. 4d corresponds to a simultaneous extension of the growth period, or hypermorphosis, of each segment. It thus formalizes what McNamara (16) called “sequential hypermorphosis.” Observing such a pattern would suggest that we are actually looking at a number of distinct developmental processes, arranged in sequence.
As a last example, consider one of the classic cases in which the nomenclature of heterochrony is applied. McKinney and McNamara (3) argue that the relation of human growth to that of chimpanzees is an example of hypermorphosis rather than neoteny, as is usually stated. Shea (17) has argued a similar case. In Fig. 5, I compare human and chimpanzee growth trajectories (18, 19) using the analysis presented above. The two trajectories can in fact be nearly overlapped, but to do so requires two different transformations: one corresponding to sequential hypermorphosis and the other to neoteny. This illustrates the value of actually carrying out the transformations. It also helps to explain why there has been disagreement as to which “kind” of heterochrony this represents.
Figure 5.

Growth trajectories for male humans (18) and chimpanzees (Pan) (19). The “Transformed Pan” curve, which closely overlaps that for humans, is derived by applying the transformations shown in Fig. 4 b and d to the chimpanzee trajectory.
Finally, if a particular case fails this test for heterochrony, how it fails can suggest where we should look next. Imagine a structure whose growth is retarded such that it comes into contact with some other structure later than it would have in the ancestor. If this new contact sets off some new set of interactions, altering the process of development in our character, then we almost certainly could not superimpose the ancestor and descendant trajectories. Fig. 6 shows such a case and applies the transformation test discussed above to it. While this comparison leads us to reject this case as an example of pure heterochrony (at the level of the entire trajectory), it also directs our attention to the particular point where the new interaction becomes important.
Figure 6.

(a) The descendant (dashed) trajectory is a neotenic version of the ancestral (solid) one up to a point, then follows a new growth function. Note that this is not revealed by simply looking at the curves. (b) The dotted line is the image of the ancestor stretched along the time axis as in Fig. 4b. Comparing this with the dashed curve reveals the point at which the descendant is no longer just a slowed down version of its ancestor.
Anyone with a healthy fear of excessive nomenclature may suspect that I am about to propose a handful of new heterochronic terms. Not so. Defining heterochrony as I propose shifts attention from the naming of categories to the comparison of trajectories. The analysis of the human–chimpanzee example above could omit all references to hypermorphosis or neoteny without losing any descriptive power.
The restricted definition of heterochrony proposed here is not at odds with those of Gould (1) and Alberch et al. (4). In fact, it is derived by specifying the conditions under which the traditional terms really apply to the study of continuous developmental processes. The concept of an ontogenetic trajectory was originally devised to facilitate the study of heterochrony. Because it does so so well, it is often not recognized that this concept actually allows for an even broader approach to the evolution of development. Much can be derived from a full study of the shape of ontogenetic trajectories, not just the endpoints. The traditional categories of heterochrony probably do represent an important subset of the ways that ontogeny can evolve, but this importance cannot be fully understood unless we carefully specify the boundaries of this set. We should apply the lexicon of heterochrony when it is appropriate, but we should not let the traditional categories get in the way of recognizing that there are far more than six ways to change size and shape in ontogeny and phylogeny.
Acknowledgments
I thank John Bonner, Leo Buss, Rich Strauss, and David Wake for valuable comments on the manuscript and the ideas. This work was supported by the Miller Institute for Basic Research in Science at the University of California, Berkeley, CA, and by a Natural Environment Research Council (London) grant to Prof. J. McGlade at the University of Warwick.
Appendix
To see what is implied when two trajectories can be simultaneously linearized, consider an ontogenetic trajectory describing phenotype (φ) as a function of time (t), and satisfying the differential equation
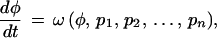 |
A1 |
where the pi are constant parameters that will hereafter be ignored. We seek a function, L(φ), such that L(φ) is a straight line when plotted against time. This function must therefore satisfy
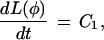 |
A2 |
where C1 is a constant. Expanding this last equation gives
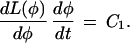 |
A3 |
Substituting Eq. A1 into Eq. A3 and rearranging, we get
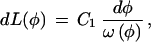 |
A4 |
which, upon integration, yields
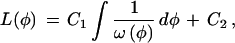 |
A5 |
where C 1 and C2 are constants that, for the purposes of comparing two different ontogenies, are arbitrary.
Thus, two different trajectories have the same linearizing transform only if their derivatives [the ω(φ)] differ by, at most, a constant multiple, which would be absorbed into the C1 value in Eq. A5. Substituting ω(φ) = rφ into Eq. A5, and setting C1 = r and C2 = 0, yields L(φ) = ln(φ), the standard transformation for an exponential growth process. Substituting ω(φ) = rφ(1 − φ) into Eq. A5, with the same constants gives the linearizing function for the logistic equation
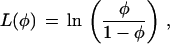 |
A6 |
References
- 1.Gould S J. Ontogeny and Phylogeny. Cambridge, MA: Belknap Harvard; 1977. [Google Scholar]
- 2.Raff R A, Kaufman T C. Embryos, Genes, and Evolution. New York: Macmillan; 1983. [Google Scholar]
- 3.McKinney M L, McNamara K J. Heterochrony: The Evolution of Ontogeny. New York: Plenum; 1991. [Google Scholar]
- 4.Alberch P, Gould S J, Oster G F, Wake D B. Paleobiology. 1979;5:296–317. [Google Scholar]
- 5.Alberch P. Syst Zool. 1985;34:46–58. [Google Scholar]
- 6.Müller G B, Wagner G P. Annu Rev Ecol Syst. 1991;22:229–256. [Google Scholar]
- 7.Raff R A, Parr B A, Parks A L, Wray G A. In: Evolutionary Innovations. Nitecki M H, editor. Chicago: Univ. Chicago Press; 1990. pp. 71–98. [Google Scholar]
- 8.Zelditch M L, Fink W L. Paleobiology. 1996;22:241–254. [Google Scholar]
- 9.De Beer G. Embryos and Ancestors. 3rd Ed. London: Oxford Univ. Press; 1958. [Google Scholar]
- 10.Godfrey L R, Sutherland M R. Am J Phys Anthropol. 1996;99:17–26. doi: 10.1002/ajpa.1330990102. [DOI] [PubMed] [Google Scholar]
- 11.Palmiter R D, Brinster R L, Hamm R E, Trumbauer M E, Rosenfeld M G, Birnberg N C, Evan R M. Nature (London) 1982;300:611–615. doi: 10.1038/300611a0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Ambros V. In: Heterochrony in evolution. McKinney M L, editor. New York: Plenum; 1988. [Google Scholar]
- 13.Shea B T, Hammer R E, Brinster R L. Endocrinology. 1987;121:1924–1930. doi: 10.1210/endo-121-6-1924. [DOI] [PubMed] [Google Scholar]
- 14.Shea B T, Hammer R E, Brinster R L, Ravosa M R. Genet Res. 1990;56:21–34. doi: 10.1017/s0016672300028846. [DOI] [PubMed] [Google Scholar]
- 15.McKinney M L. Paleobiology. 1984;10:407–419. [Google Scholar]
- 16.McNamara, K. J. (1983) in Trilobites and Other Early Arthropoda: Papers in Honour of Professor H. B. Whittington, FRS, Special Papers in Palaeontology 30, eds. Briggs, D. E. G. & Lane, P. D. (Palaeontological Association, London).
- 17.Shea B T. In: Heterochrony in Evolution. McKinney M L, editor. New York: Plenum; 1988. [Google Scholar]
- 18.Tanner J M. Foetus into Man. Cambridge, MA: Harvard Univ. Press; 1978. [Google Scholar]
- 19.Grether W F, Yerkes R H. Am J Phys Anthropol. 1940;27:181–197. [Google Scholar]