

Abstract
Fisher’s runaway process of sexual selection is potentially an important force generating character divergence between closely related populations. We investigated the evolution of multiple female preferences by Fisher’s runaway process. There are two outcomes of runaway. The first is the evolution of mate preference to a stable equilibrium. This evolution occurs if the benefits of mate choice are sufficiently large relative to the cost of choice. Alternatively, mate preferences evolve cyclically. The rate and pattern of cyclic evolution depends primarily on the individual cost of choice and epistasis in the joint cost of choice. If there are small differences in natural selection (e.g., predation risk) between populations, cyclic evolution quickly leads to divergence in mate preferences and sexual ornaments and so to sexual isolation.
Keywords: limit cycle, speciation, sexual traits, mate choice
Secondary sexual traits and courtship displays show a much greater degree of diversity than do nonsexual morphological traits over the same taxonomic range (1, 2). There are some notorious cases of diversity in sexual traits: the cichlid species flocks of the African Great Lakes, the highly species-rich Hawaiian Drosophila, male insect genitalia, and the birds of paradise (3–6). Sexual diversity also is seen between subpopulations of a species. A particularly good example is the isolated Gunnison population of American sage grouse (7). Despite little apparent ecological or morphological change in Gunnison sage grouse, male birds differ markedly in their secondary sexual characteristics. Gunnison males have whiter tail feather plumes, a higher rate of air sac inflation, and a lower rate of strutting. In addition, two novel elements have been added to their display: males wag their tails from side to side and, more dramatically, toss their neck plumes over their heads.
In a previous paper, we showed how Fisher’s runaway process can generate continual change in a sexual ornament (8). Two conditions must be met for runaway to cause perpetual cyclic coevolution of female preference for a male sexual ornament. First, the survival cost of the male trait must be small around the natural selection optimum but must increase dramatically once the male trait becomes extreme. Second, female mate preference must be accompanied by a cost. These are general and plausible assumptions. Another possibility in finite populations is that drift could cause cyclic change even in the absence of a cost to female mate preference.
In this paper, we generalize our finding. The previous model considered a single female preference for a single male trait. For a better understanding of how diversity evolves, we need to consider the simultaneous evolution of multiple sexual ornaments. In many species, it is clear that females use multiple male traits when choosing a sexual partner (9, 10). This development will enable us to address a number of questions about sexual traits. In particular, do all male sexually selected traits show cyclic evolution, what forces determine the degree of variability in male traits, and how quickly can sexual selection generate divergence between allopatric populations?
Model and Results.
We modeled the evolution of multiple female mate preferences for multiple male traits (ornaments) by Fisher’s runaway process by using a standard quantitative genetic model (11–13). In this model, sexual selection on male ornaments is generated by female mate preferences. A runaway process then ensues because of the genetic coupling of female preference with the male ornament (14). Both female preferences and male ornaments can become exaggerated in a negative (smaller than natural selection optimum) or positive (larger than natural selection optimum) directions.
Model.
For simplicity, consider the evolution of two female preferences, p1 and p2, for two male sexual ornaments, t1 and t2, respectively. This treatment easily can be extended to include further preferences and ornaments. The per generation changes in the mean value of the preferences and ornaments are (12)
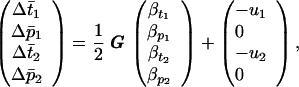 |
1 |
where
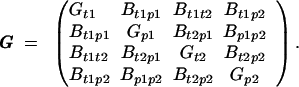 |
The factor ½ denotes sex-limited expression. Both female and male traits are assumed to have a quantitative genetic basis, and G is a matrix of the additive genetic parameters. The effect of selection is given by selection gradients for each trait
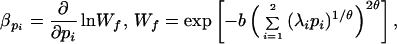 |
2 |
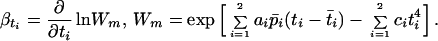 |
2a |
Female fitness (Wf) is determined by the strength of preference, p1 and p2. We assume that there is some slight cost for females in choosing a mate. This cost might be caused by predation risks, time constraints, or some other factor associated with mating behavior. Female costs are set to a minimum when females do not discriminate between potential mates (p1 = p2 = 0), and costs increase symmetrically at a rate b. The choice costs for each preference can differ according to the coefficients λ1 and λ2.
Epistatic fitness interactions between the two preferences are measured by θ, the joint cost of choice (12, 13). If θ = 0.5, then there is no interaction and the cost of each preference combines multiplicatively. If θ ≈ 0, the overall cost is no more than the maximum when either preference is considered alone. If θ ≈ 1, costs combine in a super-multiplicative manner and females using more than one preference face severe fitness loss. For instance, if females already visit a lek and spend time examining male wing plumage, then consideration of bill color at the same time will not significantly alter choice costs (i.e., θ → 0). Conversely, if females are already time-limited, then the additional time required to select males using a second ornament will increase costs significantly (i.e., θ → 1). We consider a range of θ values because we have shown already that epistasis in the joint cost of choice has a major influence on the evolutionary outcome (12, 13).
Male fitness (Wm) is the product of mating success and survivorship. Mating success is determined by female mate preference (first term of Eq. 2). The two ornaments are assumed to have independent effects on mate choice. Female preference for one male character does not extend to the other. Mean female preference can be either for males with larger (p̄1 > 0, p̄2 > 0) or smaller (p̄1 < 0, p̄2 < 0) than average ornaments (15). We assume that males can mate many times and that females have open-ended preferences (16). The coefficients a1 and a2 define the effectiveness of the two ornaments in causing increased mating success.
Male survivorship is maximized when the sizes of the male ornaments are at their respective natural selection optima, set at t1 = 0 and t2 = 0 (second term of Eq. 2). Survival chances decline symmetrically on either side of this value at a rate c1 and c2. One of the interesting features of our model is that male survival is assumed to decrease with the fourth power rather than with the quadratic as in previous models (8). This assumption means that the cost of the ornaments is very small around the natural selection optimum but increases very quickly beyond a certain limit.
We assume that there is no epistatic selection on male ornaments. Each ornament has an independent effect on mate choice and an independent effect on survival chances. This assumption is adopted to simplify the analysis (see Discussion). The only fitness interaction is in the cost of female choice.
The effect of selection depends on the additive genetic variances and covariances. The genetic variances (Gp1, Gp2, Gt1, and Gt2) are assumed to be constants. The six genetic covariances are allowed to evolve to their equilibrium values (12–14). Three are created directly through selection, either mate choice (Bt1p1 and Bt2p2) or epistasis in the cost of female preferences (Bp1p2). The others (Bt1p2, Bt2p1, and Bt1t2) arise indirectly (e.g., Bt1p2 > 0 if Bt1p1, Bp1p2 > 0). The final genetic terms in Eq. 1 are u1 and u2, which measure any mutation bias acting on the male ornaments (16).
Independent Preferences.
If the joint cost of choice θ = 0.5, then there is no interaction between the two preferences, which therefore evolve in an independent fashion. In this case, we only need to consider the evolution of a single preference and so drop the suffixes 1 and 2 as each trait pair evolves autonomously. There are two outcomes (8). The first possibility is that the population evolves to a stable equilibrium (Fig. 1a),
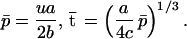 |
3 |
At equilibrium, females benefit from choosing males with exaggerated ornaments. The benefit is proportional to the mutation bias u multiplied by the ratio of the effectiveness of the male ornament in attracting females a and the cost of choice b. Males with larger ornaments have higher total fitness, causing selection in favor of greater exaggeration. This is balanced exactly by mutation bias (u) acting against further exaggeration.
Figure 1.

Evolution of the mean male ornament size (t̄) through time (mean female preference p̄ follows a similar path). The population is plotted every 40 generations. Two conditions varying in the degree of mutation bias are shown leading to a stable equilibrium (u = 0.0001) (a) and cyclic evolution (u = 0) (b). Other parameter values are a = 0.4, b = 0.001, c = 0.05, Gt = 0.5, and Gp = 0.5.
The second possibility is cyclic evolution (Fig. 1b). This occurs when the benefits of choice (i.e., mutation bias) fall below a critical threshold,
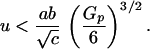 |
4 |
If this condition holds, the equilibrium defined in Eq. 3 becomes unstable. Cycles are more likely if the cost of choice b and the additive genetic variance in female preference Gp are large, relative to the degree of mutation bias u. If the equilibrium is unstable, the population undergoes a series of rapid runaway events interspersed with periods of semi-stable exaggeration (8). Semi-stability occurs when natural selection on the male ornament becomes severe and balances the mating advantage of further exaggeration. But change does not stop completely. Female preference slowly declines because of the cost of choice. Decline continues until the system crosses a critical value triggering a new runaway. In turn, this runaway slows down once natural selection on the male ornament becomes severe and a period of relative stability ensues. Again, the strength of female preference declines until another runaway is triggered in the opposite direction. No equilibrium is ever reached.
Cyclic evolution has not been recognized as a regular feature of evolution by sexual selection even though the possibility of cycles was mentioned in previous analyses (17). For cycles to appear, the equilibrium near the origin must be unstable and runaway must be bounded. In our model, this behavior is created by assuming that selection on the male ornament is a cost function with the fourth power. This makes the cost of the male ornament very small around the natural selection optimum but very large beyond a certain range. Cyclic evolution does not occur with a quadratic selection function; the only possibility is a single equilibrium given that female choice is costly (16–17).
Fisher’s runaway process occurs in such a short time as to be instantaneous on an evolutionary time scale. So, the rate of cyclic evolution is determined by the interval between successive runaways. The interval between runaways is a function of how quickly female preference declines when there is an approximate balance between sexual and natural selection on the male ornament (i.e., when βt = 0). Under this condition, the evolution of female preference is Δp̄ = Gp(−2bp̄ + ua)/2, and the approximate time between successive runaways τ ∝ 1/bGp (assuming mutation bias is very small, u ≈ 0). So, the rate of cyclic evolution is determined by the cost of choice b and genetic variation in female preference Gp (8). If the cost of choice is high, then the population does not stay in a semi-stable condition for very long before another runaway is triggered. Greater genetic variance in female preference has a similar effect because it increases the speed of response to selection against female preference when in a semi-stable condition. The other selection and genetic parameters (a, c, and Gt) do not alter the rate of cycling; they only influence the degree of exaggeration (8).
Multiple Preferences.
The evolutionary dynamics are considerably more complex when the cost of each preference interacts. Further investigation was made by simulation. To make understanding easier, we concentrated on describing the general phenomena observed. As with independent preferences, each preference and ornament can either reach a stable equilibrium or cycle.
In the extreme case, neither ornament is subject to any mutation bias (u1 = u2 = 0; Fig. 2) and both preference and ornament pairs show cyclic evolution. This is seen most simply if we equalize natural selection on the two male ornaments (a1 = a2, c1 = c2) but assume that the first preference p1 carries a greater cost than the second preference p2 (e.g., λ1 = 1, λ2 = 0.6; see Eq. 2a).
Figure 2.

Coevolution of two preferences for separate male ornaments as a function of epistasis in the joint cost of choice θ. Evolution is represented in two ways. First as a phase-space of the mean male ornaments (t̄1 and t̄2) every 20 generations; second as t̄1 (black) and t̄2 (gray) values through time. The mean female preferences are not shown because they follow similar evolutionary trajectories. The cost of choosing t1 is set to be larger than the cost of choosing t2 (λ1 = 1.0 and λ2 = 0.6). Four values of the joint cost of choice are shown: (a) θ = 0.05, t̄1 and t̄2 show equal rates of change and entrained cycles; (b) θ = 0.2, t̄1 changes three times more quickly than t̄2 and cycles are entrained; (c) θ = 0.8, t̄1 changes more quickly than t̄2 and cycles are not entrained, the phase-space shows the first 8,000 generations; and (d) θ = 1.2, females only show preference for t̄2 and progressively ignore t̄1. Parameter values are a1 = a2 = 0.4, b = 0.001, c1 = c2 = 0.05, and all G = 0.5.
The dynamics depend heavily on θ, the epistasis in the joint cost of female choice. When the joint cost of choice (θ) is very small, the population evolves in a highly ordered fashion (θ = 0.05; Fig. 2a). Following the mean values of the two male ornaments (t̄1 and t̄2) and starting in the, t̄1−, t̄2+ sector (i.e., t̄1 < 0, t̄2 > 0), there is a slow decay in both ornaments that eventually gives way to runaway from t̄1− to positive exaggeration t̄1+. Runaway is followed by slow decay and then runaway of the second ornament from t̄2+ to negative exaggeration t̄2−. This pattern of decay and runaway is repeated in a regular sequence: runaway to t̄1+, runaway to t̄2−, runaway to t̄1−, and runaway to t̄2+, returning the population to its initial state.
This pattern is adopted quickly irrespective of starting conditions (although the sequence of change can be reversed). Once runaway evolution comes to a stop, female preference always slowly decays because of the cost of choice. Eventually, female preference falls sufficiently that it crosses a critical value and triggers a new runaway. So the evolution of the mean female preferences (p̄1 and p̄2) shows the same timing of the switch from slow decline to runaway as seen in the male ornaments (t̄1 and t̄2).
At very low values of epistasis in the joint cost of choice, both ornaments cycle at the same rate (θ = 0.05; Fig. 2a). This at first seems surprising because the individual choice costs for each preference differ (λ1 = 1, λ2 = 0.6). But when θ is very small, the cost of choice is determined almost entirely by whichever preference is more exaggerated, as can be seen by setting θ ≈ 0 in Eq. 2a. To be exact, the cost of choice for both preferences is determined by max[λ1p̄1, λ2p̄2] (in the example here, max[p̄1, 0.6p̄2]). So if one preference is already exaggerated, the addition of a second preference hardly alters the overall cost of choice. This explains why there is a stepped trajectory (Fig. 2 a–c). The stepped period of slow decline in p̄1 (and hence t̄1) occurs when 0.6p̄2 > p̄1 , during which selection on p̄1 is greatly reduced. A similar effect on p̄2 (and hence t̄2) is seen when p̄1 > 0.6p̄2.
As the value of the joint cost of choice (θ) increases, the two preference and ornament pairs cycle at different rates (Fig. 3a). When θ = 0.2, the more costly preference p1 cycles three times more quickly than the cheaper preference p2 (Fig. 2b). However, the cycle times remain closely coupled and show entrainment onto regular cycle periods (Fig. 3b). This is a common phenomena in coupling of nonlinear oscillators (18–19). For example, for θ ≤ 0.1, the ratio of cycle times is 1:1, for 0.1 < θ < 0.17, the ratio is 1:2, and for 0.17 < θ < 0.39, the ratio is 1:3 (Fig. 3b). Entrainment also causes an asymmetry in the order in which ornaments are used at very low values of θ. For example, at θ = 0.05, the population spends more time in the t̄1−, t̄2+ and t̄1+, t̄2− conditions than in either t̄1+, t̄2+ or t̄1−, t̄2− (Fig. 2a). This relationship can be reversed by using different initial conditions. Entrainment breaks down with higher values of the joint cost of choice (e.g., θ = 0.8; Fig. 2c). There is no clear phase-locking of the two cycles any longer, and the periodicity of the whole system is chaotic (Fig. 3b).
Figure 3.

Length of an evolutionary cycle plotted against epistasis in the joint cost of choice (θ) when the individual cost of choice is higher for p1 than for p2 (λ1 = 1, λ2 = 0.6). Plots are given for the number of generations per cycle (p1 diamonds, p2 crosses) (a) and the ratio of p1 cycle time to p2 cycle time (b). Other parameter values as in Fig. 2.
As the joint cost of choice (θ) rises, cycles times become even shorter (Fig. 3a). This is because the overall selection on each preference increases with θ (see Eq. 2a), and this increase shortens the interval between runaway events. Eventually, when θ becomes very large, females stop using both male ornaments and concentrate on a single ornament (e.g., θ = 1.2; Fig. 2d). The two preferences cycle together for some time, but one eventually falls to 0. Either preference can be lost.
Multiple Preferences with Mutation Bias.
Significant mutation bias (u1, u2 > 0) introduces the possibility that stable equilibria exist for one or both male ornaments. If both ornaments have high mutation bias (u1 = u2 = 0.001, other parameters as in Fig. 2), then there is a single, globally stable equilibrium. At this equilibrium, females prefer males with large t1 and large t2. Stability also depends on a number of other parameters, similar to those that influence stability when preferences are independent (see Eq. 4). In contrast, if both ornaments have small mutation bias (u1 = u2 = 0.00001), then neither reaches a stable equilibrium. Both ornaments cycle as when u = 0. If there is high mutation bias in only one ornament, it alone is capable of reaching a stable equilibrium (e.g., u1 = 0.001, u2 = 0). The other ornament continues to cycle. A small degree of fluctuation is caused in the first ornament as the second goes through each runaway, but this fluctuation is insufficient to cause destabilization.
If both ornaments show different levels of mutation bias, the result is more complex. Consider the case in which mutation bias is sufficient to lead to stability in p1 but not in p2 (u1 = 0.001, u2 = 0.00001). When the joint cost of choice is small, θ < 0.5, the stable preference “captures” the cyclic preference and both preferences become stably exaggerated (Table 1a). It is simple to see why this happens. The exaggeration already present in the first preference (p̄1 > 0) reduces the cost of choice for the second cycling preference p2 when the joint cost of choice is small (θ < 0.5). So even weak mutation bias is sufficient to allow for stable preference for the second ornament.
Table 1.
Stability of two preferences dependent on the joint cost of choice θ
| Joint cost of choice θ | p1 | p2 | |
|---|---|---|---|
| (a) | 0.1 | stable | stable |
| 0.2 | stable | stable | |
| 0.3 | stable | stable | |
| 0.4 | stable | stable | |
| 0.5 | stable | cycles | |
| 0.6 | stable | cycles | |
| 0.7 | stable | cycles | |
| (b) | 0.1 | stable | cycles |
| 0.2 | stable | cycles | |
| 0.3 | stable | cycles | |
| 0.4 | stable | cycles | |
| 0.5 | stable | cycles | |
| 0.6 | cycles | cycles | |
| 0.7 | cycles | cycles |
(a) One male ornament t1 is subject to strong mutation bias u1 = 0.001 and the other t2 to weak small mutation bias u2 = 0.00001. The cost of choice b = 0.001. (b) Mutation bias is just sufficient for stable evolution of p1 but causes cyclic evolution of p2 (u1 = 0.0001, u2 = 0.00001). The cost of choice was b = 0.0023, which just satisfies Eq. 4. Other parameter values were the same in both female and male traits Gt = Gp = 0.5, a = 0.4, b = 0.001, c = 0.04, λ = 1. Simulations were started with p1, p2, t1, and t2 at high values and ran until stability or obvious cyclic behavior was observed (≈10,000 generations).
The alternative case to consider is that in which mutation bias places p1 on the borders of stability while p2 cycles (u1 = 0.0001, u2 = 0.00001). If there is no interaction between the preferences or if the joint cost is small (θ ≤ 0.5), p2 cycles do not affect p1, which remains stable (Table 1b). But if the joint choice cost is high (θ > 0.5), the cycles in p2 break down stability in p1, which ends up cycling as well. Again, it is simple to see why this happens. When the joint cost is high (θ > 0.5), p2 cycles cause periodic large increases in the cost of female choice for p1. This increase causes p1 to be pulled sufficiently far away from equilibrium to initiate runaway evolution. So, the first preference escapes from stability and cycles.
Allopatric Divergence.
Imagine that a population has been split into two distinct, noninterbreeding demes. Each deme will be subject to different selection pressures arising from different environmental conditions. For example, one environment may have fewer predators, reducing the strength of natural selection on exaggerated male ornaments (smaller c) or on female preference (smaller b or λ), habitat differences may make male displays easier to detect thereby increasing their effectiveness at attracting females (larger a), or male density may be greater, allowing females to utilize several male ornaments without severely increasing the joint cost of choice (lower θ).
If female preference has evolved to a stable equilibrium, these selective differences will alter the equilibrium strength of female preferences and the size of the male ornaments. From Eq. 3, we can see that an increase in the cost of choice (b) and a decrease in the effectiveness of the ornament (a) will cause a reduction in exaggeration in both preference and ornament, whereas an increase in the cost of the ornament (c) will only decrease the size of the male ornament. Under equilibrium conditions, small environmental differences predict modest divergence in male sexual ornaments.
Environmental differences have a much greater impact if the preference and ornament evolve cyclically. First, change in the selective conditions will cause the evolutionary dynamics of the two populations to be out of phase. The most important variables are the cost of choice (b or λ) and epistasis in the joint cost of choice (θ). These three variables alter the amount of time between runaway events (whereas to first approximation a and c only alter the degree of exaggeration). So differences in b, λ, or θ cause runaway to be triggered at different times in the two populations, resulting in preference and ornament divergence (Fig. 4).
Figure 4.

Male ornament divergence in two allopatric populations subject to different selection pressures. Selection on female preference is set to be 20% weaker in population 2, both for the individual cost of choice and epistasis in the joint cost of choice (population 1 b = 1.0, θ = 0.6; population 2 b = 0.8, θ = 0.48). Otherwise selection in the two populations is identical. There are two male ornaments, so there are four possible character states: gray, t̄1 < 0, t̄2 < 0; stipple, t̄1 > 0, t̄2 < 0; blank, t̄1 > 0, t̄2 > 0; and stripe, t̄1 < 0, t̄2 > 0. The sequence in which these character states evolve is shown through time. The two populations start with the same strength of preferences and ornament sizes. Other parameter values as in Fig. 2.
In addition, changes to the joint cost of choice (θ) will alter the relative stability of preferences in the two populations and change the sequence of character state evolution. An example is shown (Fig. 4) in which there are two preferences and four possible semi-stable states (t̄1 < 0, t̄2 < 0; t̄1 > 0, t̄2 < 0; t̄1 > 0, t̄2 > 0; and t̄1 < 0, t̄2 > 0). In population 1, the joint cost of choice (θ) is lower than in population 2. The higher joint cost of choice (θ) makes the preference with the higher individual cost (p1) evolve relatively more quickly (Fig. 3b) and so alters the order in which the ornaments evolve (Fig. 4). The female preferences and male sexual display in the two populations quickly become distinct.
Finally, different selective conditions can result in different preferences being stable in each population. Let us assume, for example, that visual predators are common in one environment, causing the cost of choice to be more severe (larger b). This severity can destabilize preferences in this environment. So, whereas visual ornaments remain constant through time in the predator-scarce environment, they undergo rapid change in the predator-common environment. Again, the result is divergence of the sexual displays of the two populations.
DISCUSSION
Sexual traits used in courtship appear to be exceedingly diverse across species (1, 2). In this paper, we have considered how Fisher’s runaway process can create this diversity. Our model shows that there are two general outcomes of the runaway process. The first is the evolution of a stable equilibrium (see Eq. 3). At equilibrium, the degree of exaggeration in female preference is determined by the costs of choice (searching for a mate, predation risks, etc.) and benefits of choice (mating with an attractive male). The associated degree of exaggeration in the male ornament is proportional to the strength of female preference divided by the cost of male display.
The second outcome is that this equilibrium is unstable, leading to cyclic evolution. Runaway leads to semi-stable exaggeration of preference for a male ornament, followed by a slow decline in preference due to the cost of choice. This eventually initiates a further runaway to a new semi-stable state, and this process then is repeated. Two conditions are required for cyclic evolutionary change (8). Selection on the male ornament must be weak about the natural selection optimum but increase rapidly beyond a certain value, and female choice must be costly. Cyclic evolution is promoted by strong costs (high b and θ) and weak benefits of choice (low u and a; see Eq. 4).
In our model of cyclic evolution, preference oscillates in evolutionary time between preference for larger than average and preference for smaller than average values of ornamentation. If the male ornament (t) were plumage color, positive t might be red and negative t green. Both are conspicuous and costly compared with gray or brown (t = 0), the optimal ornament value under natural selection. An alternative view is to interpret t as the length of a structure, for example a tail feather. Then, a negative exaggeration means preference for a tail shorter than the viability optima. There are hardly any examples of female preference for small ornaments in this sense. One possibility is the golden-rumped cisticola (Cisticola exilis) for which there is some evidence that females prefer short-tailed males (20). This rarity suggests that some asymmetry must exist that favors evolutionary exaggeration in only one direction (we intend to examine this in a future publication).
We have extended our analysis to consider how two mate preferences evolve for different male ornaments. Again, stable equilibria or cyclic evolution are seen but now in multiple dimensions. In general, preferences subject to greater individual costs (higher λ) and preferences for male ornaments that have low mutation bias (smaller u) show greater instability through time (Fig. 2). So, these preference and ornament pairs are more likely to differ between related species.
The evolutionary outcome depends greatly on the epistatic interaction between the two preferences, measured by the joint cost of choice (θ). Larger values of the joint cost of choice (θ) make cyclic evolution more likely (Table 1) and increase the rate of evolutionary change (Fig. 3). The joint cost of choice (θ) also alters the order in which traits change. When the joint cost of choice (θ) is low, the evolution of both preferences is entrained onto regular intervals between runaway events for both preferences (Figs. 2 and 3). Entrainment breaks down as the joint cost of choice (θ) increases. Finally, at very high values of the joint cost of choice (θ), only a single preference is used by females. These features arise because greater epistasis in the joint cost of choice (θ) increases selection against multiple female preferences.
We only have studied epistatic selection in the cost of choice. Other forms of interaction are possible. For instance, it is easy to imagine that male attraction is amplified (21) when two modes of signaling are used (e.g., song and color), causing epistasis in the effectiveness of male ornamentation (a). We predict that this form of epistasis will produce similar dynamical behavior. Another possible interaction is in the cost of male ornamentation (c). This interaction has been studied in combination with signal error by Johnstone (22), who presented an optimization analysis of equilibrium conditions. However, nonlinearity in the male cost function already is known to cause cyclic evolution rather than a stable equilibrium (8). So, it is more difficult to predict how this form of epistasis will influence the evolutionary dynamics.
These results demonstrate how continual evolutionary change in sexual preferences and ornaments is a natural outcome of Fisher’s runaway process. With multiple preferences and ornaments, the number of exaggerated states increases greatly. So, rather than simple cycles, we find a complex switching between different sexual phenotypes through evolutionary time. The analysis shows variability in the relative stability of different preferences. As before, lower stability and more rapid change is associated with high costs and low benefits of choice.
Our model allows predictions to be made about how allopatric populations evolve under sexual selection. If preferences show cyclic evolution, we predict that small environmental differences are amplified quickly into large population differences (Fig. 4). In particular, small differences in the cost of choice (b, λ, or θ) alter the frequency of runaway and the sequence of male ornament evolution. So, male sexual phenotypes of allopatric populations will diverge within a short period of time. This diversity will contribute to speciation if different preferences result in isolation between allopatric populations that come into secondary contact.
These observations differ in emphasis from traditional views stressing the importance of random genetic drift in triggering runaway (17, 23). Our model does not consider drift explicitly. But we can see already that divergence can be generated without any significant contribution from drift. In our analysis, the cost of choice slowly erodes any current preference, eventually causing runaway in a new direction. In small populations, the effect of drift may be of the same order as the cost of choice. Drift then may significantly alter the time between runaway events and even the direction of the ensuing runaway. Drift is thus another potential contributor to diversity.
In this paper, we have concentrated on the role of Fisher’s runaway process as a cause of diversity. Fisherian ornaments are attractive traits but have no other signaling function. An alternative possibility is that male ornaments are handicaps and act as reliable indicators of male quality (24). Handicap ornaments also show cyclic evolution of female preferences and male ornaments (Y.I. and A.P., unpublished work). However, runaway and cyclic evolution are phenomena that arise because of the coevolution of preference with an attractive ornament caused by the Fisherian process. The handicap process is an important force leading to the stability of preferences for ornaments that reveal male quality. So, preference for ornaments that yield strong handicap benefits for females (either good genes or direct benefits) will tend to remain relatively unchanged through evolutionary time. We can therefore predict that variable sexual ornaments are likely to have weaker associations with male quality (24).
Acknowledgments
We thank Takuya Kubo, Hiroyuki Matsuda, Mark Pagel, and two referees for their help and comments. We are grateful for support received from The Royal Society (A.P.) and from a Grant-in-Aid from the Ministry of Education, Science and Culture (Y.I.). Our collaboration was made possible by Y.I.’s visit to the Natural Environment Research Council Centre for Population Biology, Imperial College and by Fellowships we both held at the Wissenschaftskolleg zu Berlin.
Footnotes
This paper was submitted directly (Track II) to the Proceedings Office.
References
- 1.Darwin C R. The Descent of Man and Selection in Relation to Sex. London: John Murray; 1871. [Google Scholar]
- 2.Andersson M. Sexual Selection. Princeton: Princeton Univ. Press; 1994. [Google Scholar]
- 3.Fryer G, Iles T D. The Cichlid Fishes of the Great Lakes of Africa. Edinburgh: Oliver & Boyd; 1980. [Google Scholar]
- 4.Carson H L. In: Evolutionary Processes and Theory. Karlin S, Nevo E, editors. Orlando, FL: Academic; 1986. pp. 391–409. [Google Scholar]
- 5.Eberhard W G. Sexual Selection and Animal Genitalia. Cambridge, MA: Harvard Univ. Press; 1985. [Google Scholar]
- 6.Coates B. The Birds of Papua New Guinea: Passerines. II. Alderley, Queensland: Dove; 1990. [Google Scholar]
- 7.Young J R, Hupp J W, Bradbury J W, Braun C E. Anim Behav. 1994;47:1353–1362. [Google Scholar]
- 8.Iwasa Y, Pomiankowski A. Nature (London) 1995;377:420–422. doi: 10.1038/377420a0. [DOI] [PubMed] [Google Scholar]
- 9.Møller A P, Pomiankowski A. Behav Ecol Sociobiol. 1993;32:167–176. [Google Scholar]
- 10.Buchanan K L, Catchpole C K. Proc R Soc London Ser B. 1997;264:521–526. [Google Scholar]
- 11.Iwasa Y, Nee S, Pomiankowski A. Evolution (Lawrence, KS) 1991;45:1431–1442. doi: 10.1111/j.1558-5646.1991.tb02646.x. [DOI] [PubMed] [Google Scholar]
- 12.Barton N H, Turelli M. Genetics. 1991;127:229–255. doi: 10.1093/genetics/127.1.229. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Pomiankowski A, Iwasa Y. Proc R Soc London Ser B. 1993;253:173–181. [Google Scholar]
- 14.Iwasa Y, Pomiankowski A. Evolution (Lawrence, KS) 1994;48:853–867. doi: 10.1111/j.1558-5646.1994.tb01367.x. [DOI] [PubMed] [Google Scholar]
- 15.Fisher R A. The Genetical Theory of Natural Selection. Oxford: Clarendon; 1930. [Google Scholar]
- 16.Pomiankowski A, Iwasa Y, Nee S. Evolution (Lawrence, KS) 1991;45:1422–1430. doi: 10.1111/j.1558-5646.1991.tb02645.x. [DOI] [PubMed] [Google Scholar]
- 17.Lande R. Proc Natl Acad Sci USA. 1981;78:37221–37225. [Google Scholar]
- 18.Morita Y. Jpn J Appl Math. 1986;3:223–247. [Google Scholar]
- 19.Taylor M A, Pavlou S, Kevrekidis I G. Math Biosci. 1994;122:25–66. doi: 10.1016/0025-5564(94)90081-7. [DOI] [PubMed] [Google Scholar]
- 20.Cockburn A. An Introduction to Evolutionary Ecology. Oxford: Blackwell; 1991. [Google Scholar]
- 21.Hasson O. Proc R Soc London Ser B. 1989;235:383–406. doi: 10.1098/rspb.1989.0006. [DOI] [PubMed] [Google Scholar]
- 22.Johnstone R A. Phil Trans R Soc Lond Ser B. 1996;351:329–338. [Google Scholar]
- 23.Templeton A R. Genetics. 1980;94:1011–1038. doi: 10.1093/genetics/94.4.1011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Pomiankowski A. Oxf Surv Evol Biol. 1988;5:136–184. [Google Scholar]