

Abstract
Both positive and negative selection of immature T cells rely on engagement of their antigen-specific receptors (TCR) by peptide in association with proteins encoded in the major histocompatibility complex (MHC) protein. The decision made between these two outcomes seems to be determined by the number of TCR engaged by peptide–MHC complexes. It has been unclear how such a mechanism can be reconciled with evidence that positive and negative selection occur in different thymic compartments and are mediated by different antigen-presenting cells (APCs). In this study we demonstrate that the level of class I MHC protein is 10-fold higher on thymic dendritic cells, which mediate the negative selection of immature T cells, than on thymic epithelial cells, which mediate for positive selection. We also demonstrate that as little as a 3-fold increase in the level of a particular cognate peptide–MHC ligand is sufficient to result in negative rather than positive selection. The results suggest that quantitative differences in the level of expression of class I MHC proteins on thymic epithelial and dendritic cells contribute to the opposing roles these cells play in forming the repertoire of mature class I MHC restricted (CD8+) T cells.
Studies of T cell development indicate that T cell maturation in the thymus is largely driven by the extent to which antigen-specific receptors (T cell receptors or TCR) on immature thymocytes are engaged by peptide–major histocompatibility complex (MHC) complexes on thymic antigen-presenting cells (APCs; ref. 1). When TCR interact with complexes that are present at relatively low levels, the corresponding thymocytes are stimulated to proceed along the maturation pathway (positive selection); if, however, the complexes are relatively abundant the thymocytes undergo cell death (negative selection). The thymic APCs also have been the subject of many studies. Most reports indicate that positive selection is promoted by thymic epithelial cells whereas negative selection is mediated largely by hemopoietic dendritic cells (DC; ref. 2). Studies of thymocyte maturation in the absence of thymic DC indicate that some T cells can be positively selected by the thymic epithelial cells even when they would otherwise be negatively selected by DC (3). In an attempt to understand the basis for the different outcomes mediated by thymic epithelial and dendritic cells, we used reaggregated thymic organ cultures to investigate cellular requirements for the selection of T cells that express the antigen-specific receptor (TCR) of the CD8+ CTL clone known as 2C.
The 2C clone was generated from an H-2b mouse (Balb.B) that had been immunized with H-2d-bearing cells (4). It recognizes the allogeneic class I protein Ld in association with a naturally processed peptide, LSPFPFDL (p2Ca; ref. 5). The class I protein Kb has been identified as the positively selecting restriction element (6), and the 2C CTL line was also found to recognize the p2Ca peptide in association with Kb, albeit weakly (7, 8). More recently another naturally processed peptide, EQYKFYSV (dEV8), that can be recognized weakly in association with Kb by the 2C TCR has been identified (9).
In this study we followed the transition of double-positive (CD4+, CD8+) to single-positive (CD4−, CD8+) 2C TCR+ thymocytes from peptide-transport-defective (TAP1−/−) mice, in the presence of various concentrations of peptides, and APCs with different MHC haplotypes. Our results encompass three principal findings. First, although the presence of the allogeneic MHC protein Ld in the thymus normally results in negative selection of 2C thymocytes, we show here that these cells are positively selected by Ld bearing thymic epithelial cells, if Ld-bearing dendritic cells are absent. Second, we have identified a remarkably small range (3-fold or less) of cell surface peptide–MHC (epitope) densities on thymic epithelial cells that define the “epitope density threshold” amount of ligand above which thymocytes are negatively selected and below which they are positively selected. Finally, we found a 10-fold higher expression of class I MHC on thymic dendritic cells than on thymic epithelial cells and propose that this difference contributes to the disparate roles these cells play in thymocyte selection.
MATERIALS AND METHODS
Mice.
C57BL/6 (B6) mice were purchased from Taconic Farms. DBA/2 mice were purchased from The Jackson Laboratory. TAP1 knockout (10) and 2C TCR transgenic mice (6) were bred in our colony (Center for Cancer Research, Massachusetts Institute of Technology).
Flow Cytometric Analyses.
The following mAbs were used: R-phycoerythrin (PE)-labeled RM4–5 (anti-CD4, PharMingen), fluorescein isothiocyanate (FITC)-labeled 53-6.7 (anti-CD8a, PharMingen), PE-labeled MR5–2 (anti-Vβ8.1, 8.2, PharMingen), biotin-labeled anti-1B2 [anti-2C TCR clonotypic (4)], FITC-labeled HL3 (anti-CD11c, PharMingen), PE-labeled AF6–120.1 (anti-I-Ab, PharMingen), PE-labeled AMS-32.1 (anti-I-Ad, PharMingen), FITC-labeled 30-H12 (anti-Thy-1.2, PharMingen), FITC-labeled M1/70 (anti-Mac-1, PharMingen), MTS5 (anti-thymic epithelium, PharMingen), and FITC-labeled G53–238 (anti-rat IgM, PharMingen). Thymocyte and lymphocyte suspensions were prepared from cultured lobes or dissected tissue and analyzed as described previously (10, 11).
Cell Isolation and Reaggregated Thymic Organ Culture (RTOC).
RTOC experiments were performed as described previously (12). TAP1−/−, B6, and DBA/2 thymic epithelial cells (MHC haplotype H-2b, H-2b, and H-2d, accordingly) were prepared by disaggregating deoxyguanosine (dGuo, Sigma)-treated fetal thymic lobes using 0.05% trypsin (GIBCO), 0.02% EDTA in Ca2+, and Mg2+-free Hanks’ balanced salt solution (GIBCO). The resulting cells were analyzed with anti-I-A, anti-Mac-1 (macrophage specific), anti-CD11c (dendritic cell specific), anti-thymic epithelium (TE), and anti-Thy-1.2 antibodies. They were found to be approximately 80% I-A+, 80% TE+, 5% Mac-1+, 10% Thy-1.2+, and less than 1% CD11c+, and are referred to as thymic epithelial cells (TEC).
CD4+CD8+ (“double positive”) 2C+ thymocytes were obtained by gently grinding freshly isolated newborn 2C transgenic TAP1−/− thymus lobes. The resulting suspensions were enriched for double-positive 2C cells with biotinylated anti-CD4 antibody coupled to streptavidin-bound magnetic microbeads (Miltenyi Biotec, Auburn, CA); their purity, checked by fluorescence-activated cell sorter (FACS) analysis, was usually more than 95%. Reaggregates were formed by mixing together the desired epithelial cells and double-positive thymocytes (at a ratio of 1:2). After pelleting the cell mixture by gentle centrifugation and removing the supernatant, the pellet was dispersed into a slurry, drawn into a fine glass pipette, and placed as a standing drop on the surface of a nucleopore filter suspended over complete medium (RPMI 1640 medium supplemented with 10% fetal calf serum) with or without added peptide. Cultures were incubated for 3 days at 37°.
Thymus- and spleen-derived dendritic cells were purified from B6 and DBA/2 tissues as described previously (13). Purity was checked by staining with anti-CD11c, anti-I-Ab, and anti-I-Ad antibodies and analyzing by FACS. Splenic preparations typically were 90% dendritic cells, whereas thymic preparations usually were 70% dendritic cells, the primary contaminants being T cells.
Thymectomized, Irradiated, and Bone Marrow-Reconstituted (TIR) Mice and Thymus Transplants.
B6 mice were thymectomized as described previously (14). They were then lethally irradiated (1,100 rad) and reconstituted with 2C transgenic TAP1−/− bone marrow as described (15). In brief, bone marrow cells were isolated by flushing the tibia and femur of adult mice with ice-cold PBS. After subjecting the cells to two rounds of complement lysis using anti-CD4, anti-CD8, and anti-Thy1 antibodies and rabbit complement, approximately 5 × 106 cells in 0.5 ml PBS were injected intravenously into the TIR mice. Thymic transplants were performed by placing two or three dGuo-treated fetal thymic lobes under the left kidney capsule of these TIR animals as described previously (16). Donor thymi came from E14 embryos of either B6, DBA/2, or TAP1−/− strains.
Quantitation of Kb and Ld Complexes.
To measure the number of Kb and Ld molecules on cells from B6 and DBA/2 mice, we used a monoclonal anti-Kb antibody, Y3 (17), and a monoclonal anti-Ld antibody, 30-5-7 (18), respectively. We used bivalent antibody molecules, whose apparent affinity for the multivalent antigenic determinants on a cell surface is higher than the intrinsic affinity of monovalent Fab fragments (19), to overcome problems of sensitivity and background. We determined apparent equilibrium-binding constants (Ka) for the Y3 antibody to Kb and the 30-5-7 antibody to Ld by using Kb+ EL4 and Ld+ P815 cells, respectively. Various concentrations of 125I-labeled antibody were incubated with 2–4 × 105 cells in a total volume of 50 μl of complete medium for 30 min at room temperature (22–25°C). All reactions were carried out in duplicate. Cell-bound and unbound 125I-antibody were separated by centrifugation on fetal calf serum and measured. The concentration of specifically bound 125I-antibody was determined as the difference between cell-bound radiolabeled antibody in the presence and absence of a 100-fold molar excess of unlabeled antibody. The binding data were analyzed according to Klotz (20). The concentration of specifically bound antibody (B) was plotted as a function of the logarithm of the unbound (free) concentration (F), and experimental points were fitted to the equation:
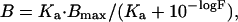 |
1 |
where Bmax is the concentration of class I bound antibody at saturation, i.e., as F approaches infinity. Values of Bmax and Ka were derived from the best fit. The Ka values were found to be 3 × 108 M−1 for the Y3 antibody and 2 × 108 M−1 for the 30-5-7 antibody (the Fab′ fragment of the Y3 antibody to cell surface Kb on the same cells had an approximately 30-fold lower Ka than the bivalent antibody, unpublished observation).
The fractional occupancy (α) of class I for any given free antibody concentration was determined from:
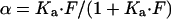 |
2 |
From the specific radioactivity of the radioiodinated antibodies and the number of cells in the analyzed samples we determined the number (Nf) of antibody molecules specifically bound to class I MHC molecules per cell at a given free antibody concentration (F). The maximal number of class I MHC bound antibody molecules per cell was taken to be Nf/α and is reported as a measure of the total number of class I molecules per cell (Table 1); the actual number of the corresponding MHC molecules per cell lies between Nf/α and twice this value.
Table 1.
dGuo-resistant thymic epithelial cells express fewer class I molecules than thymic and splenic dendritic cells
| Thymic epithelial cells
|
Thymic dendritic cells
|
Splenic dendritic cells
|
||
|---|---|---|---|---|
| Kb | Ld | Kb | Ld | Kb |
| 25,581 ± 8,907 | 26,513 ± 1,216 | 375,553 ± 140,195 | 212,600 ± 43,439 | 132,994 ± 30,064 |
Average no. MHC class I molecules per cell and standard deviations shown are based on at least two independent measurements.
To measure the number of peptide–Kb complexes formed on thymic epithelial cells at positively and negatively selecting peptide concentrations, cells from TAP1−/− thymi were incubated for 2 hours at 37° in complete medium with or without peptide followed by analysis with 125I-Y3 as described (Table 2).
Table 2.
Level of Kb expression on TAP1−/− dGuo-resistant thymic epithelial cells in the presence and absence of exogenously added peptides
| Peptide | Concentration, M | Thymic selection* | No. peptide-induced Kb molecules per cell |
|---|---|---|---|
| None | None | None | None |
| LSPFPFDL | 3 × 10−5 | Positive | 718 ± 428 |
| LSPFPFDL | 5 × 10−4 | Negative | 2,318 ± 1,243 |
| EQYKFYSV | 1 × 10−4 | Positive | 12,400 ± 1,500 |
| EQYKFYSV | 3 × 10−3 | Negative | 18,900 ± 4,040 |
| SIYRYYGL | 1 × 10−8 | Positive | ND |
| SIYRYYGL | 1 × 10−7 | Negative | ND |
Average numbers and standard deviations shown are based on at least three independent measurements. ND, not detectable by assay.
As observed in independent RTOC experiments.
RESULTS
Positive Selection of 2C Cells by Alloantigen in Reaggregation Cultures in the Absence of Thymic Dendritic Cells.
Thymocytes carrying the 2C TCR have been shown to be negatively selected in the thymus of H-2d mice (6). To identify the thymic cells involved in 2C cell selection we set up RTOCs. These cultures consisted of two cell populations: (i) immature CD4+ CD8+ 2C thymocytes (>95% pure) and (ii) cells from deoxyguanosine-treated thymic lobes, which consist of trypsinized thymic stromal cells (largely TEC) depleted of dendritic cells. The epithelial cells were obtained from H-2d, H-2b, and TAP1−/− (i.e., MHC class I deficient; ref. 10) H-2b fetuses.
As one should expect, TAP1−/− TEC did not induce either positive or negative selection of 2C TCR+ immature T cells. Interestingly, we found that mature CD8+ 2C cells were generated not only in RTOCs having H-2b epithelial cells, as expected, but also in those with H-2d epithelial cells (Fig. 1). Positive selection was less pronounced in H-2d RTOCs than in H-2b RTOCs, but was clearly above the background level seen in H-2b TAP1−/− RTOCs (Figs. 1 and 2). The observed positive selection was mediated by the TCR, because clonotypic anti-TCR Fab′ fragments (1B2, ref. 4) blocked the appearance of mature CD8+ 2C cells in the RTOC (data not shown). These findings suggest that the negative selection of 2C cells that is observed in vivo in H-2d mice is mediated largely by deoxyguanosine-sensitive cells, presumable thymic dendritic cells. To examine this possibility, we set up the same RTOC as above and added dendritic cells from spleens of H-2d mice. As shown in Fig. 2, the dendritic cells promoted deletion of 2C cells in all cultures. We conclude that 2C cells are positively selected by thymic epithelial cells, which are deoxyguanosine-insensitive, and negatively selected by H-2d dendritic cells.
Figure 1.

CD8+ 2C cells are positively selected in H-2d TAP1+/+ RTOC. Reaggregate thymic organ cultures consisting of H-2b TAP1−/−, H-2b TAP1+/+, or H-2d TAP1+/+ epithelial cells and 2C+ CD4+ CD8+ thymocytes are shown. Numbers indicate percentage of cells in the respective gated areas. Similar numbers of thymocytes were recovered from each lobe.
Figure 2.

Allogeneic dendritic cells mediate deletion of 2C cells in RTOC. Reaggregate thymic organ cultures consisting of dGuo-treated epithelial cells from H-2b TAP1−/−, H-2b TAP1+/+, or H-2d TAP1+/+ mice, and 2C+ CD4+ CD8+ thymocytes, with or without the addition of H-2d TAP1+/+ spleen-derived dendritic cells, were cultured and analyzed as before. Where indicated, the equivalent of 1.5 × 104 dendritic cells per lobe was added to equal 5% of the total epithelial cell number. The total mean number of thymocytes recovered from each lobe (n = 4) in Fig. 2 reading from left to right was (×10−3) 9.0, 10.9, 10.1, 9.4, 2.3, and 4.5, respectively. Error bars indicate the SE in each category.
Positive Selection of 2C Cells in Vivo by Alloantigen in Absence of Class I+ Dendritic Cells.
To exclude the possibility that the positive selection of 2C cells by H-2d epithelial cells was an artifact of the reaggregation cultures, we examined the selection of 2C thymocytes in vivo in thymectomized mice that were transplanted with intact deoxyguanosine-treated lobes. The same types of deoxyguanosine-treated thymus lobes that had been used for the RTOCs (H-2d, H-2b, TAP1−/− H-2b) were transplanted to thymectomized H-2b mice that had been irradiated and reconstituted with 2C transgenic, TAP1−/− bone marrow cells. The bone marrow-derived dendritic cells therefore were deficient in MHC class I expression and of the b haplotype and thus should have no (or minimal) impact on selection. Three months after transplantation very few CD8+ 2C cells were found in the lymph nodes of mice bearing H-2b TAP1−/− thymus lobes (Fig. 3) or mice that had not received any thymus lobes (data not shown). However, a considerable number of CD8+ 2C+ cells accumulated in the lymph nodes of mice receiving intact H-2b or H-2d thymus lobes. These CD8+ 2C cells did not express the activation markers CD25 or CD44 (data not shown), indicating that they have been selected positively but have not been significantly expanded in the periphery by antigenic stimulation.
Figure 3.

Allogeneic thymic epithelium positively selects 2C cells in vivo. Lymph nodes were collected and analyzed (see Materials and Methods) from animals having received either TAP1−/− H-2b (A), TAP1+/+ H-2b (B), or TAP1+/+ H-2d (C) thymic transplants. The high proportion of CD4+ cells observed may be because of either the expansion of some mature CD4 cells present in the transplanted bone marrow or of cells that arose from some surviving host bone marrow. For each type of transplant two mice were studied with similar results.
Differences in the Level of Expression of Class I MHC Proteins on Thymic Epithelial and Dendritic Cells.
Many endogenous peptides are likely to be generated at the same levels in different cells (21), and it is conceivable that differential expression of MHC, and therefore of cell-surface densities of peptide–MHC complexes, may be responsible for the opposing roles that epithelial and dendritic cells play in positive and negative selection, respectively. Dendritic cells are well known to express MHC proteins at high levels (3). However, a direct quantitative comparison of MHC expression on thymic epithelial and dendritic cells has not been reported. Using 125I-labeled antibodies specific for class I proteins, we determined the number of Kb and Ld MHC molecules on thymic dendritic cells and dGuo-resistant thymic epithelial cells. Both proteins were found at about 10-fold higher densities on thymic dendritic cells (2.8 × 105/cell) than on epithelial cells (2.5 × 104/cell) (Table 1). We also examined the number of Kb MHC molecules on splenic dendritic cells and found that these cells express class I MHC proteins at an intermediate level (Table 1), which is consistent with previous observations made with flow cytometry (13).
Peptide-Induced Class I MHC Expression Required for Positive and Negative Selection.
Both positive and negative selection of 2C thymocytes can be achieved with the addition of appropriate amounts of peptide ligand. Fig. 4 shows FACS analyses of RTOCs in which optimal positive and negative selection of 2C cells occurred at 10 and 100 nM of SIYRYYGL peptide (22), respectively, whereas no selection occurred without added peptide or with an irrelevant peptide (data not shown). The concentrations were determined from several peptide titration experiments. Peptide concentrations were considered to be optimal for positive selection at the dose at which the most CD8+ 2C T cells developed and optimal for negative selection at the lowest dose that eliminated nearly all double-positive cells and resulted in near-background levels of CD8+ 2C T cells. Two other peptides also were found to elicit positive and negative in RTOC although at very different concentrations. The peptide p2Ca (LSPFPFDL) could positively and negatively select at 30 and 500 μM, respectively, whereas dEV8 (EQYKFYSV) did so at 100 and 3,000 μM (data not shown). We compared the levels of peptide-stabilized Kb on purified thymic epithelium at these concentrations by using 125I-labeled antibody specific for Kb. The increase in the number of peptide-stabilized Kb complexes from positively to negatively selecting concentrations of peptide was only 3-fold for p2Ca and even less for dEV8 (Table 2). We could not reliably detect SIYRYYGL-induced Kb expression above the background level with the peptide at a total concentration of either 10 or 100nM (Table 2). Thus, as little as a 3-fold difference in the density of a particular peptide–MHC ligand on the APC surface can determine whether a cell is positively or negatively selected.
Figure 4.

Peptide-induced positive and negative selection in RTOC. Reaggregate thymic organ cultures consisting of H-2b TAP1−/− epithelial cells and 2C+ CD4+ CD8+ thymocytes with or without positively and negatively selecting concentrations peptide as indicated. Numbers indicate the percentage of cells in the respective gated areas.
DISCUSSION
Studies of thymocyte development have shown that the fate of double-positive thymocytes can be altered in peptide transport-deficient thymic organ cultures by varying concentrations of peptide, which presumably stabilize proportional amounts of MHC proteins on thymic APCs (1). At relatively low concentrations peptides promote positive selection and at high concentrations they lead to negative selection. These effects can be accounted for by an “avidity” model, which assumes that the cell’s fate is determined by the extent to which its TCR are engaged by peptide–MHC complexes in the thymic environment (1). This view stresses the importance of the affinity of the developing cell’s TCR for peptide–MHC complexes and the abundance of these complexes in the thymic environment. At first glance, it does not account for the finding that different types of APC have divergent effects on thymocytes; i.e., thymic epithelial cells mediate positive selection and dendritic cells mediate negative selection (3). Indeed, thymic epithelium-bearing H-2 Ld, an alloreactive restriction element for the 2C TCR, will positively select 2C thymocytes, whereas H-2 Ld-bearing dendritic cells will induce negative selection both in vitro and in vivo (Figs. 1–3). The disparate effects of epithelial and dendritic cells have been attributed to differences in various cell surface molecules involved in cell–cell interactions, such as LFA-1/ICAM-1 (23–25), CD28/B7.1 (26, 27), CD40/gp39 (28, 29), Fas/FasL (30), and CD30/CD30L (31). Experiments with knockout mice and specific inhibitors have suggested a role for these molecules in thymic selection, but also revealed that they are not always absolutely necessary (23–32).
The most important finding in this study is that the cell-surface level of class I MHC molecules on dendritic cells is approximately 10-fold higher than on epithelial cells (Table 1). Taken in conjunction with the avidity model, this finding could account for the different roles of these APC in shaping the TCR repertoire of newly developed single-positive CD8+ T cells. Thus, if the pool of peptides produced in these APC was similar, those with the higher total MHC level would be expected to have a correspondingly higher level of any particular peptide–MHC complex and to be more likely to elicit negative selection than APC with lower levels. The MHC class I level on thymic dendritic cells is not only higher than that on thymic epithelial cells, but also higher than that on splenic dendritic cells (Table 1). It is conceivable that the very high MHC class I expression in the thymus increases the likelihood that potentially autoreactive immature T cells are eliminated by negative selection in the thymus. There are other precedents for the natural exploitation of differential MHC expression. The down-regulation of MHC levels by a variety of mechanisms helps many cancer cells and virus-infected cells to escape destruction by cytotoxic T cells (33–35). Large differences in the levels of MHC proteins between cells in the periphery may favor antigen presentation by some cells and minimize the risks of inadvertent immune injury of other cells (36).
Although thymic dendritic cells are largely responsible for negative selection, it is important to note that negative selection can occur in their absence (2). Thus, using thymic cell preparations consisting essentially of just double-positive thymocytes and epithelial cells, we observed both positive and negative selection, depending on the concentration of added peptide. With this simplified system, we were able to examine closely the effects of small changes in the abundance of peptide–Kb complexes (“epitope density’) on the surface of thymic epithelial cells. The results are of particular interest with peptides p2Ca and dEV8, because both are naturally processed peptides derived from ubiquitous proteins (5, 9). The finding that approximately 700 p2Ca-Kb complexes per cell was associated with positive selection of 2C TCR+ thymocytes (Table 2) may be compared with an earlier estimate of about 100–200 p2Ca peptide molecules per mouse thymus cell (5). The former value (700 complexes) is probably an overestimate (because it is based on incubation of cells with peptide for 2 hr whereas the RTOC assays were carried out over 3 days, during which time peptide concentrations drop, perhaps because of proteases in the culture medium), and the latter value (100–200 peptide molecules) was probably too low, because it did not take into account losses incurred during peptide isolation. It is thus possible that the p2Ca peptide normally exists in the thymus at a level where it contributes to positive selection under natural conditions. The natural abundance of the dEV8 peptide has not been estimated (9), but it is doubtful that it is anywhere near the levels needed to approach the epitope density values shown in Table 2.
Surprisingly, the difference between an epitope level that led to optimal positive selection and one that led to near-total negative selection was only 3-fold in the case of the peptide p2Ca and even less for the peptide dEV8. These remarkably small differences suggest that we have defined for each of these peptides a narrow epitope density zone, below which thymocytes are stimulated to make the transition from double-positive to single-positive cells and above which they are triggered to undergo cell death. If we consider the midpoint between positively and negatively selecting values for a given peptide as a “threshold” value for the corresponding peptide–Kb complex, we see that the threshold is about 1,500 complexes per cell for the p2Ca peptide and around 16,000 complexes per cell for the dEV8 peptide. For the SIYRYYGL peptide, the epitope density threshold could not be defined because the concentrations used to elicit positive and negative selection were too low. It is apparent, nevertheless, that in terms of required peptide concentrations, the order of effectiveness was SIYRYYGL > p2Ca > dEV8. This order matches the efficacy of these peptides in cytolytic assays (Y.S. and H.E., unpublished data).
For the SIYRYYGL and p2Ca peptides the differences are in accord with the large differences in the affinity of the 2C TCR for these peptide–MHC complexes. Measured on intact CD8+ 2C cells, the values are 1 × 107 M−1 for SIYRYYGL-Kb (Y.S. et al., unpublished data) and 3 × 103 M−1 for p2Ca-Kb (8). The corresponding affinity value for dEV8-Kb has not been measured under the same conditions, but recently has been reported to be 1.2 × 104 M−1, as determined by surface plasma resonance using immobilized 2C TCR and soluble dEV8-Kb complexes (37). Equilibrium constants measured by the two methods so far have differed considerably, being 10- to 300-fold higher on intact cells (presumably because of the contribution of CD8 coreceptors, which are absent in the cell-free system). Thus, the affinity of the 2C TCR on intact cells is likely to be much greater for dEV8-Kb than for p2Ca-Kb. Given this difference, it is notable that dEV8-Kb is far less effective than p2Ca-Kb in promoting the maturation of 2C thymocytes. A possible explanation is suggested by the relative instability of the complex formed by the 2C TCR with dEV8-Kb (37). If the corresponding TCR bond with p2Ca-Kb were more stable, the disparity between the epitope density thresholds for p2Ca-Kb and dEV8Kb might be accounted for. Alternatively, the explanation may be structural rather than kinetic: it is possible that different conformations of the TCR, resulting from ligation with different peptide–MHC ligands, could have confounding effects that are responsible for the dEV8/p2Ca disparity.
Acknowledgments
We thank Dennis Y. Loh for contribution of 2C transgenic mice, Channel Lovett and Carol McKinley for technical assistance, Christiaan Levelt and Suzana Marusic-Galesic for advice and discussion, Emily Rossie for secretarial assistance, and Werner Haas for discussion and critical reading of the manuscript. This work was supported in part by grants from the National Institutes of Health/National Cancer Institute (R35-CA53874, CA-60686, AI-34247) and a training grant in immunology (R37-AI17879).
ABBREVIATIONS
- RTOC
reaggregated thymic organ culture
- dGuo
2′-deoxyguanosine
- TEC
thymic epithelial cell
- DC
dendritic cell
- TCR
T cell receptor
- APC
antigen-presenting cell
- MHC
major histocompatibility complex
References
- 1.Ashton-Rickardt P, Bandeira A, Delaney J, van Kaer L, Pircher H-P, Zinkernagel R, Tonegawa S. Cell. 1994;76:651–663. doi: 10.1016/0092-8674(94)90505-3. [DOI] [PubMed] [Google Scholar]
- 2.Anderson G, Moore N, Owen J, Jenkinson E. Annu Rev Immunol. 1996;14:73–99. doi: 10.1146/annurev.immunol.14.1.73. [DOI] [PubMed] [Google Scholar]
- 3.van Ewijk K. Annu Rev Immunol. 1991;9:591–615. doi: 10.1146/annurev.iy.09.040191.003111. [DOI] [PubMed] [Google Scholar]
- 4.Kranz D, Tonegawa S, Eisen H. Proc Natl Acad Sci USA. 1984;81:7922–7926. doi: 10.1073/pnas.81.24.7922. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Udaka K, Tsomides T, Eisen H. Cell. 1992;69:989–998. doi: 10.1016/0092-8674(92)90617-l. [DOI] [PubMed] [Google Scholar]
- 6.Sha W, Nelson C, Newberry R, Kranz D, Russel J, Loh D. Nature (London) 1988;336:73–76. doi: 10.1038/336073a0. [DOI] [PubMed] [Google Scholar]
- 7.Dutz J, Tsomides T, Kageyama S, Rasmussen M, Eisen H. Mol Immun. 1994;31:967–975. doi: 10.1016/0161-5890(94)90091-4. [DOI] [PubMed] [Google Scholar]
- 8.Sykulev Y, Brunmark A, Jackson M, Cohen R, Peterson P, Eisen H. Immunity. 1994;1:15–22. doi: 10.1016/1074-7613(94)90005-1. [DOI] [PubMed] [Google Scholar]
- 9.Tallquist M, Yun T, Pease L. J Exp Med. 1996;184:1017–1026. doi: 10.1084/jem.184.3.1017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Van Kaer L, Ashton-Rickardt P, Pleogh H, Tonegawa S. Cell. 1992;71:1205–1214. doi: 10.1016/s0092-8674(05)80068-6. [DOI] [PubMed] [Google Scholar]
- 11.Ashton-Rickardt P, van Kaer L, Schumacher T, Ploegh H, Tonegawa S. Cell. 1993;73:1041–1049. doi: 10.1016/0092-8674(93)90281-t. [DOI] [PubMed] [Google Scholar]
- 12.Anderson G, Jenkinson E, Moore N, Owen J. Nature (London) 1993;362:70–73. doi: 10.1038/362070a0. [DOI] [PubMed] [Google Scholar]
- 13.Crowley M, Inaba K, Witmer-Pack M, Steinman R. Cell Immunol. 1989;118:108–125. doi: 10.1016/0008-8749(89)90361-4. [DOI] [PubMed] [Google Scholar]
- 14.Sjodin K, Dalmasso A, Smith J, Martinez C. Transplantation. 1963;1:521–525. doi: 10.1097/00007890-196301040-00011. [DOI] [PubMed] [Google Scholar]
- 15.Ildstad S, Sachs D. Nature (London) 1984;307:168–170. doi: 10.1038/307168a0. [DOI] [PubMed] [Google Scholar]
- 16.Von Boehmer H, Schubiger K. Eur J Immunol. 1984;14:1048–1052. doi: 10.1002/eji.1830141116. [DOI] [PubMed] [Google Scholar]
- 17.Hammerling G, Rusch E, Tada N, Kimura S, Hammerling U. Proc Natl Acad Sci USA. 1982;79:4737–4741. doi: 10.1073/pnas.79.15.4737. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Ozato K, Hansen T, Sachs D. J Immunol. 1980;125:2473–2477. [PubMed] [Google Scholar]
- 19.Hornick C, Karush F. Immunochemistry. 1972;9:325–340. doi: 10.1016/0019-2791(72)90096-1. [DOI] [PubMed] [Google Scholar]
- 20.Klotz I. Science. 1982;217:1247–1249. doi: 10.1126/science.6287580. [DOI] [PubMed] [Google Scholar]
- 21.Marrack P, Ignatowicz L, Kappler J, Boymel J, Freed J. J Exp Med. 1993;178:2173–2183. doi: 10.1084/jem.178.6.2173. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Udaka K, Wiesmüller K, Kienle S, Jung G, Walden P. J Immunol. 1996;157:670–678. [PubMed] [Google Scholar]
- 23.Fine J, Kruisbeek A. J Immunol. 1991;147:2852–2859. [PubMed] [Google Scholar]
- 24.Carlow D, Teh S, Teh H. J Immunol. 1992;148:2988–2995. [PubMed] [Google Scholar]
- 25.Kishimoto H, Cai Z, Brunmark A, Jackson M, Peterson P, Sprent J. J Exp Med. 1996;184:531–537. doi: 10.1084/jem.184.2.531. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Degermann S, Suhr C, Glimcher L, Sprent J, Lo D. J Immunol. 1994;152:3254–3263. [PubMed] [Google Scholar]
- 27.Punt J, Osborne B, Takahama Y, Sharrow S, Singer A. J Exp Med. 1994;179:709–713. doi: 10.1084/jem.179.2.709. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Foy T, Aruffo A, Bajorath J, Buhlmann J, Noelle R. Annu Rev Immunol. 1996;14:591–617. doi: 10.1146/annurev.immunol.14.1.591. [DOI] [PubMed] [Google Scholar]
- 29.Foy T, Page D, Waldschmidt T, Schoneveld A, Laman J, Masters S, Tygrett L, Ledbetter J, Aruffo A, Claassen E, et al. J Exp Med. 1995;182:1377–1388. doi: 10.1084/jem.182.5.1377. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Nagata S, Suda T. Immunol Today. 1995;16:39–43. doi: 10.1016/0167-5699(95)80069-7. [DOI] [PubMed] [Google Scholar]
- 31.Amakawa R, Hakem A, Kundig T, Matsuyama T, Simard J, Timms E, Wakeham A, Mittreucker H, Griesser H, Takimoto H, et al. Cell. 1996;84:551–562. doi: 10.1016/s0092-8674(00)81031-4. [DOI] [PubMed] [Google Scholar]
- 32.Shahinian A, Pfeffer K, Lee K, Kündig T, Kishihara K, Wakeham A, Kawai K, Ohashi P, Thompson C, Mak T. Science. 1993;261:609–612. doi: 10.1126/science.7688139. [DOI] [PubMed] [Google Scholar]
- 33.Restifo N, Esquivel F, Asher A, Stotter H, Barth R, Bennink J, Mule J, Yewdell J, Rosenberg S. J Immunol. 1991;147:1453–1459. [PMC free article] [PubMed] [Google Scholar]
- 34.Hill A, Jugovic P, York I, Russ G, Bennink J, Yewdell J, Ploegh H, Johnson D. Nature (London) 1995;375:411–415. doi: 10.1038/375411a0. [DOI] [PubMed] [Google Scholar]
- 35.Wiertz E, Jones T, Sun L, Bogyo M, Geuze H, Ploegh H. Cell. 1996;84:769–779. doi: 10.1016/s0092-8674(00)81054-5. [DOI] [PubMed] [Google Scholar]
- 36.Joly E, Mucke L, Oldstone M. Science. 1991;253:1283–1285. doi: 10.1126/science.1891717. [DOI] [PubMed] [Google Scholar]
- 37.Garcia K, Tallquist M, Pease L, Brunmark A, Scott C, Degano M, Stura E, Peterson P, Wilson I, Teyton L. Proc Natl Acad Sci USA. 1997;94:13838–13843. doi: 10.1073/pnas.94.25.13838. [DOI] [PMC free article] [PubMed] [Google Scholar]