
Figure 2. Quinine increases Cl− permeability in airway epithelia through a paracellular pathway associated with decreased epithelial resistance.

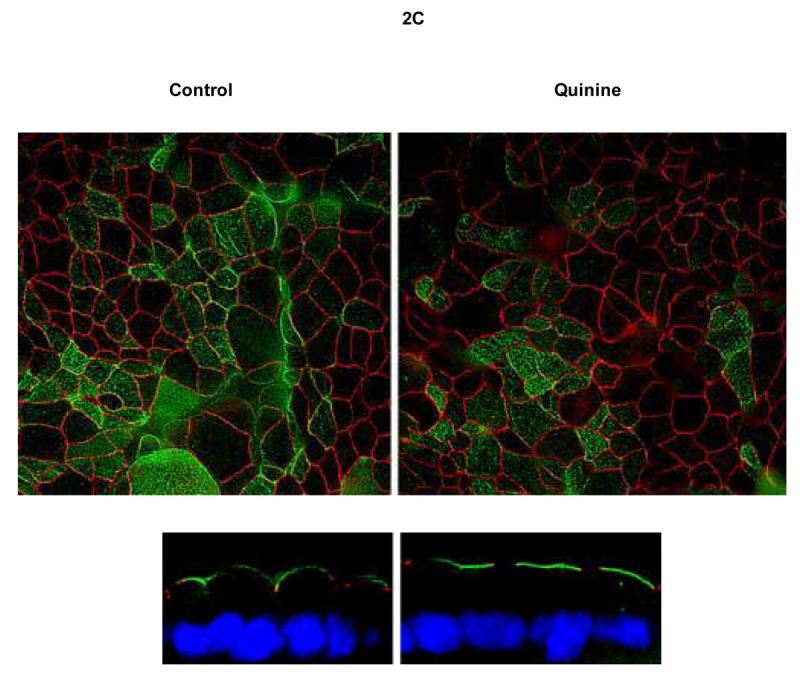

A: In Calu-3 monolayers, apical quinine (1 mg/mL) increased the monolayer permeability (19.4 ± 3.4 μA/cm2, as also shown in Figure 1A,* p < 0.001 compared with untreated controls, n = 16, ± SEM) in the presence of a Cl− secretory gradient (low Cl− on mucosal surface) and was only partially blocked by the addition of glybenclamide (change of 6.9 ± 1.8 μA/cm2, **p< 0.05 vs. apical quinine, n=16, ± SEM). The effect of quinine applied to the basolateral surface was less pronounced than apical administration (**p<0.05 vs. apical quinine, n=16, ± SEM). Bumetanide applied to the basolateral surface several minutes after quinine had no effect compared to apical quinine alone. The change attributable to quinine decreased significantly upon omission of the chloride gradient (‡ p <0.001 vs. apical quinine in Cl− secretory gradient, n=16, ± SEM). In a reversed chloride gradient, the direction of apparent Cl− movement was reversed (‡‡ p <0.001 vs. apical quinine in Cl− secretory gradient, n = 12, ±SEM). B: In Calu-3 monolayers, apical quinine (1 mg/mL) added in a Cl− secretory gradient caused a decrease in airway epithelial resistance (*p= 0.004, n=16, ±SEM) in Calu-3 cells. C: Quinine (1 mg/mL) had no effect on gross epithelial morphology in Calu-3 monolayers studied for localization of tight junctions (Z0-1, green) and CFTR (red). D: Effects of quinine (1 mg/mL) on Ussing chamber recording and transepithelial electrical resistance (E) in primary airway surface epithelial cell monolayers (n = 8 filters tested, ±SEM).