

Abstract
In response to hypoxia the hypoxia-inducible factor-1 (HIF-1) mediates transcriptional activation of a network of genes encoding erythropoietin, vascular endothelial growth factor, and several glycolytic enzymes. HIF-1 consists of a heterodimer of two basic helix–loop–helix PAS (Per/Arnt/Sim) proteins, HIF-1α and Arnt. HIF-1α and Arnt mRNAs are constitutively expressed and were not altered upon exposure of HeLa or HepG2 cells to hypoxia, suggesting that the activity of the HIF-1α–Arnt complex may be regulated by some as yet unknown posttranscriptional mechanism. In support of this model, we demonstrate here that Arnt protein levels were not increased under conditions that induce an hypoxic response in HeLa and HepG2 cells. However, under identical conditions, HIF-1α protein levels were rapidly and dramatically up-regulated, as assessed by immunoblot analysis. In addition, HIF-1α acquired a new conformational state upon dimerization with Arnt, rendering HIF-1α more resistant to proteolytic digestion in vitro. Dimerization as such was not sufficient to elicit the conformational change in HIF-1α, since truncated forms of Arnt that are capable of dimerizing with HIF-1α did not induce this effect. Moreover, the high affinity DNA binding form of the HIF-1α–Arnt complex was only generated by forms of Arnt capable of eliciting the allosteric change in conformation. In conclusion, the combination of enhanced protein levels and allosteric change by dimerization defines a novel mechanism for modulation of transcription factor activity.
Oxygen tension is an important regulator of mammalian gene expression (1), yet despite much attention, the cellular sensoring mechanism for low oxygen remains unidentified. Hypoxia-inducible genes can also be activated by exposure of target cells to certain transition metals such as cobalt or to iron-chelating agents such as desferrioxamine, suggesting that at least one key component in this pathway is a heme protein (2). One physiologically important oxygen-regulated system is induction of erythropoietin (EPO) gene expression in response to hypoxia due to, for instance, anemia. In hypoxic cells, transcription of the EPO gene is rapidly elevated, and this induction response has been localized to enhancer regions of the gene; sequences collectively termed hypoxia response elements. In transient transfection assays using reporter genes, the induction response conferred by these response elements has been demonstrated in a number of cell lines. Moreover, other hypoxia-responsive genes such as vascular endothelial growth factor and a number of glycolytic enzymes have also been documented to be regulated by hypoxia response elements (reviewed in ref. 3).
Hypoxia response elements of target genes are recognized and regulated by the hypoxia-inducible factor 1 (HIF-1), a heterodimeric complex comprising the factors HIF-1α and Arnt (4). These two proteins are transcription factors that contain a basic helix–loop–helix (bHLH) DNA binding and dimerization motif contiguous with a conserved region, PAS (Per/Arnt/Sim), that is also found in the Drosophila circadian rhythm regulator Per, the Drosophila neurodevelopmental factor Sim and its mammalian homologues, and the mammalian dioxin (aryl hydrocarbon) receptor (5–7). Thus, the PAS domain defines a novel subclass, bHLH/PAS, of the broad family of bHLH transcription factors (see ref. 8 for a review).
The PAS domain consists of two hydrophobic repeat motifs, A and B, and has been demonstrated to function as a dimerization interface in Per (9), Arnt (10, 11), and the dioxin receptor (11, 12). In the case of the dioxin receptor, the ligand binding domain has been demonstrated to span the PAS B motif (13, 14). The Arnt factor, in addition to being important for hypoxia signaling (4, 15, 16) is also a critical partner factor of the dioxin receptor (reviewed in refs. 5 and 6). In addition, it appears that Sim also requires Arnt to recognize asymmetric target DNA (17, 18). Moreover, Arnt is a constitutively active transcriptional regulator on symmetric E box motifs, possibly as a homodimeric complex (19, 20). A distinct hierarchy in affinity for Arnt has been documented between different bHLH/PAS factors, resulting in, under certain conditions, competition for recruitment of Arnt between hypoxia and dioxin signaling pathways (16).
Previously, it has been suggested that in response to hypoxia (or CoCl2), both mRNA and protein levels of HIF-1α and Arnt are induced from very low basal levels in Hep3B cells (4), However, contrary to this model of regulation, we and others have detected significant levels of constitutively expressed HIF-1α mRNA in a number of cell lines and human tissues (16, 21), and these steady-state levels are not altered by exposure to hypoxia. In our efforts to understand the mechanism of regulation of bHLH/PAS factors, we have examined whether the activity of HIF-1α, similarly to that of the dioxin receptor (5, 6), can be regulated at the posttranscriptional level. In support of this model we demonstrate here that HIF-1α protein levels are dramatically and rapidly elevated after induction of the hypoxic response. Clearly, recruitment of Arnt is critical to enable HIF-1α to bind to hypoxia response elements in vitro. In addition, upon dimerization with HIF-1α, Arnt produced an allosteric change in the conformation of its partner factor to generate a DNA binding complex.
MATERIALS AND METHODS
Cell Culture, and Preparation and Analysis of Cell Extracts.
Human HeLa and HepG2 cells and mouse Hepa 1c1c7 c4 cells were routinely maintained in Eagle’s minimal essential medium supplemented with 10% fetal calf serum, l-glutamine (2 mM), penicillin (50 units/ml), and streptomycin (50 μg/ml). Cytosolic extracts were prepared from Hepa 1c1c7 c4 cells as described (11). To mimic hypoxic induction, HeLa were seeded at a density of 1.5 × 106 cells onto 10-cm plates and after 16 h were treated either with 100 μM CoCl2 or vehicle only for an additional 24 h. Whole cell extracts were prepared as described (22). Briefly, cells were harvested in TEN buffer (40 mM Tris-HCl, pH 7.9/10 mM EDTA, pH 8.0/150 mM NaCl) followed by a wash with ice-cold PBS. The cell pellet was immediately frozen in liquid nitrogen, thawed, and resuspended in 200 μl of whole cell extract buffer [10 mM Hepes, pH 7.9/400 mM NaCl/0.1 mM EDTA/5% (vol/vol) glycerol/1 mM dithiotreitol/1 mM phenylmethylsulfonyl fluoride], followed by centrifugation 4°C for 30 min at maximal velocity in a microcentrifuge. The resulting supernatant was stored at −70°C until required. Protein concentrations of the extracts was measured using the Bio-Rad protein assay reagent. For Western blot analysis, 20 μg of extract was separated on a 7.5% SDS/polyacrylamide gel and transferred to a nitrocellulose filter. Immobilized proteins were incubated for 2 h at room temperature with rabbit anti-Arnt (23), diluted 1:200 in PBS/0.1% Tween-20. Horseradish peroxidase-conjugated swine anti-rabbit antiserum (Dako) was used as a secondary antibody (1:500 dilution in 1% nonfat milk in PBS). After extensive washing with PBS/0.1% Tween-20, the complexes were visualized using enhanced chemiluminiscence (Amersham) according to the manufacturer’s instructions. HIF-1α antiserum was generated in rabbits against a bacterially expressed C-terminal fragment encompassing amino acids 529–650 of human HIF-1α (4). For Northern (RNA) blot analysis, total RNA was isolated by acid guanidium isothiocyanate-phenol-chloform extraction as described (24), and RNA levels were prehybridized, hybridized, and washed using standard procedures. 32P-labeled HIF-1α and β-actin probes (16) were prepared by random priming.
Partial Proteolysis of HIF-1α.
Wild-type HIF-1α was synthesized and labeled with [35S]methionine in a coupled cell-free transcription–translation kit (Promega) according to the manufacturer’s instructions. HIF-1α was incubated with either bacterially expressed, purified glutathione S-transferase (GST) or GST–Arnt fusion protein (I.P. and L.P., unpublished data), or with vaccinia virus-expressed, histidine-tagged, purified Arnt (19) at room temperature for 1 h in a reaction buffer containing 40 mM Hepes (pH 7.5), 120 mM KCl, 5 mM MgCl2, and 0.5 mM EDTA. After the initial incubation, Staphylococcus aureus endoproteinase Glu-C (V8 protease; Boehringer Mannheim) was added, and the samples were incubated for a further 30 min at 30°C. The reaction products were separated on a 12.5% SDS/polyacrylamide gel, fixed in 45% (vol/vol) methanol, 10% (vol/vol) acetic acid, treated with Amplify (Amersham), and dried, and radioactive peptides were detected by fluorography.
Interaction Assays in Vitro.
[35S]Methionine-labeled HIF-1α was incubated with various GST–Arnt fusion constructs at room temperature for 1 h before addition of glutathione-Sepharose (Pharmacia), and incubation continued on ice for 1 h. Resulting complexes were washed four times with 1 ml of PBS containing 0.5% (vol/vol) Triton X-100 before analysis on 7.5% SDS/polyacrylamide gels. To study in vitro association of HIF-1α with hsp90, coimmunoprecipitation experiments were performed as described (16, 23). Briefly, full-length HIF-1α, synthesized either in reticulocyte or wheat germ lysates and labeled with [35S]methionine, was incubated with monoclonal anti-hsp90 IgM antibody 3G3 (Affinity Bioreagents, Golden, CO) and subsequently immunoprecipitated using goat anti-mouse IgM (Sigma) coupled to CNBr-activated Sepharose 4B (Pharmacia). After washing four times with buffer (25 mM Mops, pH 7.5/1 mM EDTA/0.02% NaN3/10% glycerol) supplemented with 20 mM molybdate and 2 mM dithiotreitol, the precipitated proteins were analyzed by SDS/PAGE and fluorography.
Electrophoretic Mobility Shift Assay.
Electrophoretic mobility shift assay was performed using standard procedures. Briefly, in vitro-translated unlabeled HIF-1α was allowed to interact with bacterially expressed wild-type Arnt or Arnt fragments (in the nanogram range) at room temperature for 30 min before addition of buffer (final concentrations: 10 mM Hepes, pH 7.9/100 mM KCl/0.1 mM EDTA/3 mM MgCl2/4 mM spermidine/0.5 mM dithiothreitol/10% glycerol/20 ng/μl tRNA/1 ng/μl salmon sperm DNA), and the hypoxia response element-containing 18-bp oligonucleotide (5′-GCC CTA CGT GCT GTC TCA-3′) from the erythropoietin enhancer (25) as radiolabeled probe. The incubation was then continued for an additional 30 min at 4°C. Complexes were resolved on a 4% nondenaturing polyacrylamide (1:29) gel run in 1× TGE buffer (50 mM Tris/2.7 mM EDTA/380 mM glycine) at 4°C. Antibody supershift analysis was performed using rabbit antibodies raised against HIF-1α or control (nonimmune) antiserum. For oligonucleotide competition experiments, either hypoxia response element- or glucocorticoid response element-containing oligonucleotides (26) were used. The DNA-binding activity of the dioxin receptor–Arnt complex was monitored by gel mobility shift analysis as described (11, 16).
RESULTS
Analysis of HIF-1α and Arnt Expression Levels in HeLa Cells.
We have previously shown that HIF-1α mRNA levels are constitutively expressed in a large number of human tissues and human and rodent cell lines and are thus unaltered upon treatment with hypoxia (16), suggesting that induction of functional HIF-1α activity could be regulated by a posttranscriptional mechanism. We therefore analyzed whether HIF-1α protein expression levels were altered under conditions that mimic the hypoxic response. To this end, HeLa cells were treated either with 100 μM CoCl2 [i.e., conditions under which hypoxia-inducible genes are stimulated (2)] or with vehicle only. Cells were harvested after 4 or 24 h of treatment, and both whole cell protein extracts or total RNA were prepared. No changes in Arnt protein levels were detected as assessed by immunoblot analysis (Fig. 1; compare lanes 5 and 6). In a similar fashion, Arnt mRNA levels remain unaltered in a number of CoCl2-treated cell lines (16). In contrast, when HeLa cells were assayed for HIF-1α protein expression levels, a dramatic increase (more than 20-fold) was detected, with maximal levels observed following 4 h of treatment with CoCl2 (Fig. 1; compare lanes 1 and 2). This induction response was also seen following 2 h of treatment but it was not detected at the 0.5-h time point (data not shown). A very similar, dramatic induction of HIF-1α protein expression levels was also observed in HepG2 cells treated under identical conditions (4 h of treatment with 100 μM CoCl2; data not shown) or in cells exposed to hypoxia for 4 h (1% oxygen; Fig. 1C). Induced HIF-1α levels persisted at least for an additional 20 h. As previously described (4), cellular or in vitro synthesized HIF-1α is detected as a series of isoforms with molecular mass of ≈110–130 kDa (Fig. 1).
Figure 1.
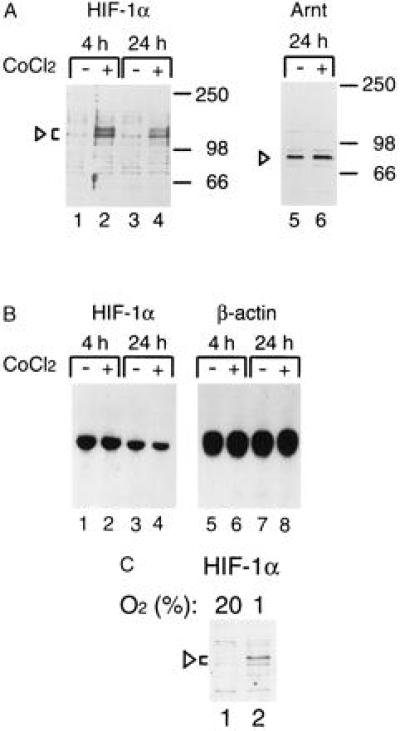
Hypoxic (CoCl2) induction of HIF-1α protein levels in HeLa cells. (A) HeLa cells were either treated with vehicle only (lanes 1, 3, and 5) or induced with 100 μM CoCl2 for 4 or 24 h, and whole cell extracts were prepared. Twenty-microgram aliquots of the extracts were separated on a 7.5% SDS/polyacrylamide gels and transferred onto a nitrocellulose filter. The transferred proteins were detected with rabbit anti-HIF-1α antiserum (1:400, lanes 1–4) or with rabbit anti-Arnt antibody (1:200 dilution, lanes 5 and 6) and visualized by chemiluminescence. Arrowheads indicate the specific HIF-1α and Arnt complexes. (B) Total cellular RNA was prepared from duplicate samples corresponding to lanes 1 to 4 in A and analyzed by RNA (Northern) blot for HIF-1α mRNA levels. As a control, the same blot was rehybridized with a β-actin probe (lanes 5–8). (C) Induction of HIF-1α protein levels by hypoxia. HepG2 cells were incubated under normoxic (20% oxygen, lane 1) or hypoxic (1% oxygen, lane 2) for 4 h, and whole cell extracts were prepared. Immunoblot analysis was performed as in A.
In RNA blot analysis, no up-regulation of HIF-1α mRNA expression levels was observed when untreated and CoCl2-treated HeLa cells were compared over a 24-h period (Fig. 1B, compare lanes 1–2 and 3–4). On the contrary, consistent with earlier observations in HepG2 cells treated under hypoxic conditions or with CoCl2 (16), a subtle (maximally 2-fold) down-regulation of HIF-1α mRNA expression levels was detected after 24 h of treatment. Taken together, these data suggest that a key regulatory step in the induction of HIF-1α activity may occur via a posttranscriptional mechanism(s).
Recruitment of Arnt Induces a Conformational Change in HIF-1α.
As documented for certain transcription factors (27), a plausible mechanism for inducing functional activity, resulting in enhanced transcription of a target promoter, may be provided by allosteric regulation of the activator. For instance, in the case of heterodimeric transcriptional activators, it is possible that the dimerization event may induce the formation of novel interaction surface(s) to facilitate functional communication with positive or negative cofactors and/or components of the basal transcription machinery. We therefore examined whether interaction with Arnt could produce detectable changes in the HIF-1α protein conformation. In these experiments we allowed in vitro synthesized, [35S]methionine-labeled HIF-1α to dimerize with either vaccinia virus-expressed or bacterially produced GST–Arnt fusion proteins before subjecting the complexes to limited proteolysis by S. aureus endoproteinase Glu-C (V8 protease) (Fig. 2). In the absence of any added protein (data not shown), or in the presence of control vaccinia virus extracts or purified GST protein alone, in vitro synthesized HIF-1α was rapidly degraded in the presence of Glu-C (Fig. 2, lanes 2, 4, and 6). However, in the presence of either vaccinia virus-expressed (Fig. 2A, lanes 3 and 5) or bacterially expressed (lane 7) Arnt, a major ≈50 kDa protease-resistant fragment was generated. Given the presence of more than 50 cleavage sites for Glu-C in the primary sequence of HIF-1α, these results suggest a marked alteration in the conformation upon dimerization with Arnt.
Figure 2.
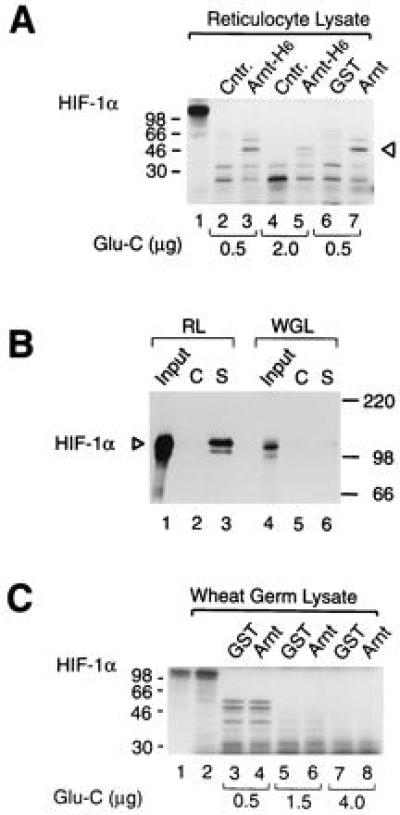
Arnt induces a conformational change in HIF-1α. (A) HIF-1α was in vitro translated in reticulocyte lysate and allowed to interact with vaccinia virus-expressed, histidine-tagged, and purified Arnt (Arnt-H6) or control (Cntr.) extracts (lanes 2–5) or with bacterially expressed, purified GST, or GST–Arnt fusion protein (lanes 6 and 7) at room temperature for 1 h. A total of 0.5 μg (lanes 2, 3, 6, and 7) or 2 μg (lanes 4 and 5) of S. aureus Glu-C protease was added, and the samples were incubated for 30 min at 30°C. Reaction products were run on a 12.5% SDS/polyacrylamide gel and visualized by fluorography. The Arnt-induced protease-resistant product is indicated by an arrowhead. Lane 1 represents full-length HIF-1α with no added protease. (B) Wheat germ lysate-expressed HIF-1α is not associated with hsp90. [35S]Methionine labeled full length HIF-1α, synthesized either in reticulocyte (RL) or in wheat germ lysate (WGL), was coimmunoprecipitated with either monoclonal anti-hsp90 (lanes S) IgM antibody or control (lanes C) IgM antibody as described. Immunoprecipitated proteins were analyzed by 7.5% SDS/PAGE and fluorography. Lanes 1 and 4 represent 10% of the inputs of in vitro synthesized HIF-1α. (C) Wheat germ lysate-expressed HIF-1α does not undergo an allosteric change in the presence of Arnt. Wheat germ lysate-translated HIF-1α (lane 2) was treated with either 0.5 μg (lanes 3 and 4), 1.5 μg (lanes 5 and 6), or 4.0 μg (lanes 7 and 8) Glu-C protease for 30 min at 30°C in the presence of GST (lanes 3, 5, and 7) or full-length GST–Arnt (lanes 4, 6 and 8). The reaction products were separated on a 12.5% SDS/polyacrylamide gel and analyzed by fluorography. Lane 1 represents HIF-1α expressed in reticulocyte lysate.
In view of the similarities in the general domain structure and the high degree of amino acid identity between the dioxin receptor and HIF-1α within these domains, we have previously demonstrated that in vitro translated HIF-1α, in striking analogy to the dioxin receptor, is associated with the molecular chaperone hsp90 (16). hsp90 appears to be critical for folding of a functional form of the dioxin receptor since association with hsp90 is correlated with maintenance of a high affinity ligand binding and ligand responsive conformation of the receptor (28–30). We next expressed HIF-1α by in vitro translation in wheat germ lysate which does not contain a functional homologue of mammalian hsp90 that is able to form a complex with the dioxin receptor (ref. 31 and references therein). Consistent with these observations, we were unable to detect any association of [35S]methionine-labeled HIF-1α with the wheat germ homologue of hsp90 in coimmunoprecipitation assays (Fig. 2B). As expected (16), under identical conditions, reticulocyte lysate-synthesized HIF-1α was efficiently coprecipitated by the hsp90-specific antibodies directed against hsp90 but not by the control antibodies (Fig. 2C, compare lanes 2 and 3). We next tested wheat germ lysate-expressed [35S]methionine-labeled HIF-1α for the appearance of any Arnt-induced, protease-resistant fragment. Under all experimental conditions tested we failed to observe the allosteric change in HIF-1α conformation in the presence of Arnt (Fig. 2C), suggesting that hsp90 may play an important role in formation of the Arnt-sensitive form.
To identify the domains of Arnt required to induce the conformational change in HIF-1α we expressed certain Arnt deletion mutants as GST fusion proteins in Escherichia coli. As schematically represented in Fig. 3A, Arnt1-140 contains only the bHLH region of Arnt, whereas Arnt1-407 spans both the bHLH and PAS domains. Both the bHLH and PAS domains of Arnt serve as dimerization interfaces with the ligand-activated form of the dioxin receptor (10, 11), and, in a similar fashion, both are also important for dimerization with HIF-1α (15, 16). Whereas the bHLH motif is critical for interaction of Arnt with HIF-1α, the PAS domain of Arnt appears to stabilize complex formation with HIF-1α (16). Thus, in dimerization experiments, the bHLH motif encompassed within the Arnt fragment Arnt1-140 was sufficient to mediate significant, albeit weak, complex formation with HIF-1α (Fig. 3B). In these experiments in vitro-synthesized HIF-1α was incubated with either GST alone or with the various GST–Arnt fusion proteins before precipitation with glutathione-Sepharose. Although the bHLH domain was sufficent to mediate dimerization with HIF-1α, complex formation was significantly enhanced in the presence of the PAS motif, consistent with the postulated role of the PAS domain in protein dimerization (9, 11). In fact, the dimerization activity of HIF-1α with the bHLH/PAS fragment Arnt1-407 was virtually identical to that of the full-length Arnt–GST fusion protein (Fig. 3B, compare lanes 2–5), suggesting that the PAS domain of Arnt is of critical importance to stabilize complexes with HIF-1α. In contrast, GST alone did not recognize HIF-1α (Fig. 3B).
Figure 3.
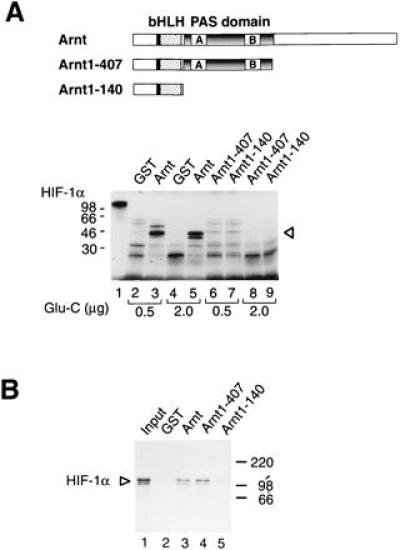
HIF-1α does not undergo an allosteric change in response to the GST–Arnt1-407 or GST–Arnt1-140 deletion mutants. (A) The Upper panel shows a schematic representation of full-length Arnt and the bacterially expressed Arnt bHLH and bHLH/PAS fragments Arnt1-140 and Arnt1-407, respectively. Full-length, [35S]methionine-labeled HIF-1α was expressed in reticulocyte lysate and incubated with either GST (lanes 2 and 4), full-length GST–Arnt (lanes 3 and 5), GST–Arnt1-407 (lanes 6 and 8), or GST–Arnt1-140 (lanes 7 and 9). Glu-C protease (0.5 μg: lanes 2, 3, 6, and 7; or 2.0 μg: lanes 4, 5, 8, and 9) was added and the incubation continued for an additional 30 min before analysis by SDS/PAGE and fluorography. (B) The bHLH and PAS regions of Arnt are sufficient for dimerization with HIF-1α. Ten microliters aliquots of in vitro synthetized and [35S]methionine-labeled HIF-1α expressed in reticulocyte lysate were incubated with either purified GST (lane 2) or GST–Arnt fusion proteins (lanes 3–5) for 1 h at room temperature. Twenty microliters of glutathione-Sepharose was added, and the samples were incubated on ice for 1 h. The precipitated pellet was analyzed on a 7.5% SDS/polyacrylamide gel. Lane 1 represents 10% of the HIF-1α input.
In limited proteolysis studies of HIF-1α we used under identical conditions either the full-length Arnt–GST fusion protein or the Arnt deletion fragments Arnt1-140 or Arnt1-407 (Fig. 3B). Surprisingly, although both Arnt1-140 and Arnt1-407 dimerized with HIF-1α, neither of these proteins were able to induce a conformational change in HIF-1α, as monitored by proteolytic digestion (Fig. 3B). These results suggest that protein–protein interaction per se between Arnt and HIF-1α is not sufficient to induce the allosteric change. It therefore appears that regions C terminal to the PAS domain of Arnt are required to produce this effect.
To further correlate these observations to activation of HIF-1α to a DNA-binding form we performed electrophoretic mobility shift assays. In analogy to the experiments above, we used in vitro translated HIF-1α in combination with bacterially expressed full-length Arnt or the Arnt deletion proteins. As shown in Fig. 4, neither Arnt nor HIF-1α were able to recognize the erythropoietin HIF-response element containing probe (Fig. 4A, lanes 2 and 3). However, in the presence of both HIF-1α and the full-length Arnt protein, a specific, novel protein–DNA complex was generated (Fig. 4A, compare lanes 2–4; complex indicated by an arrowhead). Formation of this complex was completely inhibited by the presence of an excess of specific HIF-response element containing competitor oligonucleotide but not by an identical excess of an oligonucleotide containing the unrelated glucocorticoid response element (compare lanes 5 and 6). Addition of antisera raised against HIF-1α to the binding reactions resulted in disruption of the protein–DNA complex, whereas the control serum had no effect (lanes 10 and 9, respectively). However, in contrast to full-length Arnt, incubation of in vitro translated HIF-1α with either GST–Arnt1-407 or GST–Arnt1-140 resulted in only very low levels of protein–DNA complex formation (Fig. 4, lanes 7 and 8). Thus, the failure of these mutants to induce a conformational change correlated with their inability to generate a DNA binding form of HIF-1α. Although Arnt1-407 failed to produce a DNA binding complex with HIF-1α, it could generate a DNA binding complex with the ligand activated form of the dioxin receptor. In these experiments, both full-length Arnt and Arnt1-407 showed a similar potency in generating a protein–DNA complex with the ligand activated form of the dioxin receptor (Fig. 4B, lanes 3 and 4; complexes indicated with arrows). Consistent with the results obtained with HIF-1α, Arnt1-140 did not induce any detectable levels of DNA-binding activity of the dioxin receptor. We were unable to detect any alterations in the digestion pattern when the dioxin receptor—either liganded or unliganded—was used instead of HIF-1α in proteolytic cleavage assay (data not shown). In conclusion, Arnt1-407 showed DNA binding activity in the presence of the ligand-occupied dioxin receptor, demonstrating functional activity of the bacterially overexpressed protein. However, the same protein failed to generate a protein–DNA complex with HIF-1α, supporting the idea that dimerization-induced conformational alteration is required for the activation of HIF-1α function.
Figure 4.
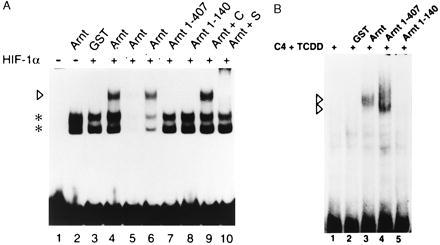
Full-length HIF-1α forms a DNA-binding complex with bacterially expressed Arnt. Six microliters of reticulocyte lysate-expressed HIF-1α was allowed to interact with either GST (lane 3), full-length Arnt (lanes 4–6, 9, and 10), Arnt1-407, or Arnt1-140 (lanes 7 and 8), respectively. In lanes 5 and 6, a 100-fold molar excess of unlabeled competitor DNA was added (specific and nonspecific, respectively), whereas in lanes 9 and 10 control (C) or specific (S) anti-HIF-1α antibodies were added to the reactions. The arrowhead indicates the HIF-1α–Arnt complex, whereas the asterisks denote nonspecific protein DNA complexes formed with unprogrammed reticulocyte lysate only (lane 2). Lane 1 represents free probe in the absence of any added protein. (B) The dioxin receptor forms a DNA binding complex with either full-length Arnt or Arnt1-407. Hepa 1c1c7 C4 cell cytosol was treated with 5 nM dioxin for 2 h to activate the dioxin receptor. Forty micrograms of dioxin-treated cytosol was incubated with GST (lane 3), full-length Arnt, Arnt1-407, or Arnt1-140 (lanes 4–6, respectively) at room temperature for 45 min. A 32P-labeled xenobiotic response element (XRE)-containing oligonucleotide was added as a probe and incubation continued for additional 15 min before electrophoresis. Lane 1, 32P-labeled XRE alone; lane 2, 32P-labeled XRE in the presence of the cytosolic extract without any added recombinant protein. Dioxin receptor–Arnt complexes are indicated by arrowheads.
DISCUSSION
Several recent studies indicate that the bHLH/PAS transcription factor HIF-1α mediates transcriptional activation of genes essential for erythropoiesis and angiogenesis in response to decreased levels of tissue or cell oxygenation (see ref. 3 for a review). Although HIF-1α is structurally related to the dioxin receptor, a ligand-activated transcription factor (reviewed in refs. 5 and 6), the mechanism of conditional regulation of HIF-1α function has not been elucidated. What mechanism triggers activation of HIF-1α and cellular responses to hypoxia? Given the structural relatedness to the dioxin receptor, it is plausible that HIF-1α may be regulated by an as yet unidentified class of ligands—e.g., metabolites generated in response to changes in tissue oxygenation. Alternatively, activation can be achieved by other means, such as phosphorylation. We have previously demonstrated that HIF-1α and dioxin receptor mRNA levels are constitutively expressed in a large number of human cell lines and tissues (16). In contrast to these observations, we detected in the present study only very low levels of expression of HIF-1α protein in normoxic cells. Upon initiation of the hypoxic response by stimulation of cells with CoCl2, however, HIF-1α protein levels were dramatically (more than 20-fold) up-regulated, suggesting that HIF-1α expression, and possibly function, is regulated by a posttranscriptional mechanism. It will now be important to investigate whether this mechanism involves mRNA and/or protein stabilization or a distinct translational regulatory process.
We do not know whether induction of HIF-1α protein levels is the only critical determinant of HIF-1α activity, or whether there exist additional levels of regulation of HIF-1α function. In the case of the dioxin receptor, the protein is functionally repressed and localized in the cytoplasmic compartment of the cell in the absence of ligand (ref. 32 and references therein). However, although cytoplasmic retention of HIF-1α in the normoxic cell is a plausible mechanism, this may be of only limited functional significance since the HIF-1α protein is barely detectable under normoxic conditions. Yet it is also possible that HIF-1α protein, when expressed at elevated levels in hypoxic cells, may also be inactive and require further modification for conversion to a functional form. Interestingly, we observed that dimerization with Arnt produced a conformational change in HIF-1α, rendering HIF-1α more resistant to proteolysis in vitro. Moreover, induction of the conformational change correlated with activation of DNA binding activity of HIF-1α, since mutants of Arnt, most notably the bHLH/PAS fragment Arnt1-407, showed wild-type levels of dimerization activity but failed to generate a DNA binding HIF-1α–Arnt complex and to induce the conformational change within HIF-1α. These results strongly argue that the DNA binding bHLH/PAS domain may be subject to conformational regulation by a region of Arnt C terminal of the PAS domain. It remains to be determined whether this structural change is transduced over a long range to other functional domains of HIF-1α. In the case of members of the steroid hormone receptor family, both DNA and ligand binding allosterically modulate transcriptional regulatory functions by possibly inducing long-range conformational changes (33–35). These allosteric effects may, in turn, modulate the interaction of nuclear receptors with transcriptional coactivators and corepressors (see ref. 36 for a review). Interestingly, recent studies indicate that CBP/p300, and its associated protein SRC-1, are critical cofactors both for HIF-1α activity and for signaling by certain members of the steroid receptor family including the glucocorticoid and retinoic acid receptors (37–39).
In conclusion, as summarized in the model in Fig. 5, there appear to exist two distinct levels of regulation of HIF-1α: induction of HIF-1α protein levels and an allosteric change upon recruitment of Arnt. We have previously demonstrated that HIF-1α is associated with the molecular chaperone hsp90 (16). In the case of certain members of the steroid receptor family and the dioxin receptor hsp90 is important for folding of the ligand binding domain and possibly other functional domains (reviewed in ref. 6). HIF-1α synthesized in wheat germ lysate did not show any association with hsp90, and, interestingly, was not allosterically modified in the presence of Arnt, consistent with a role of hsp90 in folding of HIF-1α. However, the specific target of any chaperoning function of hsp90 on HIF-1α has not yet been identified. As Arnt appears to predominantely reside in the cell nucleus in a constitutive manner (32), and given the very high affinity of Arnt for HIF-1α in vitro (16), it is likely that HIF-1α heterodimerizes with Arnt as soon as these factors are exposed to one another. In both HeLa and HepG2 cells induced levels of HIF-1α exceeded those of Arnt, raising the possibility that there may exist two pools of HIF-1α: HIF-1α/Arnt heterodimers, and a pool of HIF-1α that may either be functionally inert or interact with other cofactors. Conceivably therefore, Arnt expression levels, and/or expression of Arnt-2 (40), may constitute important determinants of hypoxia responsiveness, further illustrating the central role of Arnt in regulation of HIF-1α function.
Figure 5.
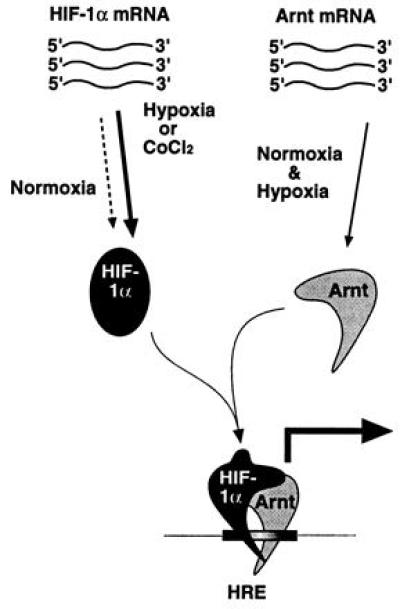
Model for activation of HIF-1 function. Both Arnt mRNA and protein are constitutively expressed. In contrast, hypoxia treatment results in dramatically elevated HIF-1α protein levels generated from a constitutively expressed pool of mRNA. HIF-1α–Arnt heterodimers then form, and dimerization induces a conformational change that allows the HIF-1α–Arnt complex to bind DNA. It is also possible that the structural change is propagated onto other functional domains of HIF-1α, most notably the transactivation domain. The present data indicate that the molecular chaperone hsp90 associates with de novo synthesized HIF-1α and gives rise to a conformation that is critical for dimerization with Arnt.
Acknowledgments
This work was supported by grants from the Swedish Cancer Society. P.J.K. is financially supported by a Marie Curie Research Training Grant from the European Commission.
ABBREVIATIONS
- HIF-1
hypoxia-inducible factor 1
- bHLH
basic helix–loop–helix
- PAS
Per/Arnt/Sim
- GST
glutathione S-transferase
References
- 1.Bunn H F, Poyton R O. Physiol Rev. 1996;76:839–885. doi: 10.1152/physrev.1996.76.3.839. [DOI] [PubMed] [Google Scholar]
- 2.Goldberg M A, Dunning S P, Bunn H F. Science. 1988;242:1412–1415. doi: 10.1126/science.2849206. [DOI] [PubMed] [Google Scholar]
- 3.Wang G L, Semenza G L. Curr Opin Hematol. 1996;3:156–162. doi: 10.1097/00062752-199603020-00009. [DOI] [PubMed] [Google Scholar]
- 4.Wang G L, Jiang B H, Rue E A, Semenza G L. Proc Natl Acad Sci USA. 1995;92:5510–5514. doi: 10.1073/pnas.92.12.5510. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Hankinson O. Annu Rev Pharmacol Toxicol. 1995;35:307–340. doi: 10.1146/annurev.pa.35.040195.001515. [DOI] [PubMed] [Google Scholar]
- 6.Poellinger L. In: Inducible Gene Expression. Baeuerle P A, editor. Vol. 1. Boston: Birkhäuser; 1995. pp. 177–205. [Google Scholar]
- 7.Ema M, Morita M, Ikawa S, Tanaka M, Matsuda Y, Gotoh O, Saijoh Y, Fujii H, Hamada H, Kikuchi Y, Fujii-Kuriyama Y. Mol Cell Biol. 1996;16:5865–5875. doi: 10.1128/mcb.16.10.5865. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Littlewood T D, Evan G I. Protein Profile. 1995;2:621–702. [PubMed] [Google Scholar]
- 9.Huang Z J, Edery I, Rosbash M. Nature (London) 1993;364:259–262. doi: 10.1038/364259a0. [DOI] [PubMed] [Google Scholar]
- 10.Reisz-Porszasz S, Probst M R, Fukunaga B N, Hankinson O. Mol Cell Biol. 1994;14:6075–6086. doi: 10.1128/mcb.14.9.6075. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Lindebro M C, Poellinger P, Whitelaw M L. EMBO J. 1995;14:3528–3539. doi: 10.1002/j.1460-2075.1995.tb07359.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Fukunaga B N, Probst M R, Reisz-Porszasz S, Hankinson O. J Biol Chem. 1995;270:29270–29278. doi: 10.1074/jbc.270.49.29270. [DOI] [PubMed] [Google Scholar]
- 13.Dolwick K M, Swanson H I, Bradfield C A. Proc Natl Acad Sci USA. 1993;90:8566–8570. doi: 10.1073/pnas.90.18.8566. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Whitelaw M L, Göttlicher M, Gustafsson J-Å, Poellinger L. EMBO J. 1993;12:4169–4179. doi: 10.1002/j.1460-2075.1993.tb06101.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Wood S M, Gleadle J M, Pugh C W, Hankinson O, Ratcliffe P J. J Biol Chem. 1996;271:15117–15123. doi: 10.1074/jbc.271.25.15117. [DOI] [PubMed] [Google Scholar]
- 16.Gradin K, McGuire J, Wenger R H, Kvietikova I, Whitelaw M, Toftgård R, Tora L, Gassmann M, Poellinger L. Mol Cell Biol. 1996;16:5221–5231. doi: 10.1128/mcb.16.10.5221. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Wharton K A, Jr, Franks R G, Kasai Y, Crews S T. Development (Cambridge, UK) 1994;120:3563–3569. doi: 10.1242/dev.120.12.3563. [DOI] [PubMed] [Google Scholar]
- 18.Swanson H I, Chan W K, Bradfield C A. J Biol Chem. 1995;270:26292–26302. doi: 10.1074/jbc.270.44.26292. [DOI] [PubMed] [Google Scholar]
- 19.Antonsson C, Arulampalam V, Whitelaw M L, Pettersson S, Poellinger L. J Biol Chem. 1995;270:13968–13972. doi: 10.1074/jbc.270.23.13968. [DOI] [PubMed] [Google Scholar]
- 20.Sogawa K, Nakano R, Kobayashi A, Kikuchi Y, Ohe N, Matsushita N, Fujii-Kuriyama Y. Proc Natl Acad Sci USA. 1995;92:1936–1940. doi: 10.1073/pnas.92.6.1936. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Tian H, McKnight S L, Russel D W. Genes Dev. 1997;11:72–82. doi: 10.1101/gad.11.1.72. [DOI] [PubMed] [Google Scholar]
- 22.Berghard A, Gradin K, Pongratz I, Whitelaw M L, Poellinger L. Mol Cell Biol. 1993;13:677–689. doi: 10.1128/mcb.13.1.677. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.McGuire J, Whitelaw M L, Pongratz I, Gustafsson J-Å, Poellinger L. Mol Cell Biol. 1994;14:2438–2446. doi: 10.1128/mcb.14.4.2438. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Chomczynski P, Sacchi N. Anal Biochem. 1987;162:156–159. doi: 10.1006/abio.1987.9999. [DOI] [PubMed] [Google Scholar]
- 25.Semenza G L, Wang G L. Mol Cell Biol. 1992;12:5447–5454. doi: 10.1128/mcb.12.12.5447. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Pongratz I, Strömstedt P-E, Mason G F, Poellinger L. J Biol Chem. 1991;266:16813–16817. [PubMed] [Google Scholar]
- 27.Roberts S G E, Green M G. Nature (London) 1994;371:717–720. doi: 10.1038/371717a0. [DOI] [PubMed] [Google Scholar]
- 28.Pongratz I, Mason G F, Poellinger L. J Biol Chem. 1992;267:13728–13734. [PubMed] [Google Scholar]
- 29.Carver L A, Jackiw V, Bradfield C A. J Biol Chem. 1994;269:30109–30112. [PubMed] [Google Scholar]
- 30.Whitelaw M L, McGuire J, Picard D, Gustafsson J-Å, Poellinger L. Proc Natl Acad Sci USA. 1995;92:4437–4441. doi: 10.1073/pnas.92.10.4437. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Antonsson C, Whitelaw M L, McGuire J, Gustafsson J-Å, Poellinger L. Mol Cell Biol. 1995;15:756–765. doi: 10.1128/mcb.15.2.756. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Pollenz R S, Sattler C A, Poland A. Mol Pharmacol. 1994;45:428–438. [PubMed] [Google Scholar]
- 33.Lefstin J A, Thomas J R, Yamamoto K R. Genes Dev. 1994;8:2842–2856. doi: 10.1101/gad.8.23.2842. [DOI] [PubMed] [Google Scholar]
- 34.Kurokawa R, DiRenzo J, Boehm M, Sugarman J, Gloss B, Rosenfeld M G, Heyman R A, Glass C K. Nature (London) 1994;371:528–531. doi: 10.1038/371528a0. [DOI] [PubMed] [Google Scholar]
- 35.Forman B M, Umesono K, Chen J, Evans R M. Cell. 1995;81:541–550. doi: 10.1016/0092-8674(95)90075-6. [DOI] [PubMed] [Google Scholar]
- 36.Horwitz K B, Jackson T A, Bain D L, Richer J K, Takimoto G S, Tung L. Mol Endocrinol. 1996;10:1167–1177. doi: 10.1210/mend.10.10.9121485. [DOI] [PubMed] [Google Scholar]
- 37.Arany Z, Huang L E, Eckner R, Bhattacharya S, Jiang C, Goldberg M A, Bunn H F, Livingston D M. Proc Natl Acad Sci USA. 1996;93:12969–12973. doi: 10.1073/pnas.93.23.12969. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Kamei Y L, Heinzel T, Torchia J, Kurokawa R, Gloss B, Lin S, Heyman R A, Rose D W, Glass C K, Rosenfeld M G. Cell. 1996;85:403–414. doi: 10.1016/s0092-8674(00)81118-6. [DOI] [PubMed] [Google Scholar]
- 39.Chakravarti D, LaMorte V J, Nelson M C, Nakajima T, Schulman I G, Juguilon H, Montminy M, Evans R M. Nature (London) 1996;383:99–103. doi: 10.1038/383099a0. [DOI] [PubMed] [Google Scholar]
- 40.Hirose K, Morita M, Ema M, Mimura J, Hamada H, Fujii H, Saijo Y, Gotoh O, Sogawa K, Fujii-Kuriyama Y. Mol Cell Biol. 1996;16:1706–1713. doi: 10.1128/mcb.16.4.1706. [DOI] [PMC free article] [PubMed] [Google Scholar]