

Abstract
The lpr gene encodes a defective form of Fas, a cell surface protein that mediates apoptosis. This defect blocks apoptotic deletion of autoreactive T and B cells, leading to lymphoproliferation and lupus-like autoantibody production. The effects of the lpr Fas mutation on other kinds of physiologically relevant apoptosis are largely undocumented. To assess whether some of the apoptosis known to occur after ionizing radiation might be mediated by Fas/Fas ligand (FasL) interactions, we quantitated in vitro apoptosis by flow cytometry measurement of DNA content in splenic T and B cells from irradiated 5- to 8-month-old B6/lpr mice. Total apoptosis of both lpr and control cells was substantial after treatment; however there was a significant difference between B6 (73%) and lpr (25%) lymphocyte apoptosis. Thy1, CD4, CD8, and IgM cells from lpr showed much lower levels of apoptosis than control cells after irradiation. Apoptosis induced by heat shock was also impaired in lpr. The finding that γ-irradiation increased Fas expression on B6 cells and that irradiation-induced apoptosis could be blocked with a Fas–Fc fusion protein further supported the possible involvement of Fas in this form of apoptosis. Fas/FasL interactions may thus play an important role in identifying and eliminating damaged cells after γ-irradiation and other forms of injury.
Fas (CD95) is a cell surface protein that mediates rapid apoptosis when crosslinked by antibody or by Fas ligand (FasL) (1). Recently, the autoimmune and lymphoproliferative disease of lpr and gld mice has been attributed to defects in the genes encoding Fas and FasL, respectively (2). Mice which are homozygous for the autosomal recessive lpr gene produce autoantibodies similar to those seen in human systemic lupus erythematosus and develop massive lymphadenopathy (1). The lymph nodes and spleens of lpr mice are characterized by progressive infiltration with CD4−CD8−Thy1+ T cells that bear surface molecules which are not characteristic of normal resting T cells, such as B220, PC.1, CD69, Ly-6C, and Ly-24 (1).
The involvement of Fas in antigen-specific apoptosis, both through the T cell antigen receptor (TCR) (3–7) and through membrane Ig (8, 9), is well documented. The consequences of the Fas and FasL mutations might be solely due to defective antigen-driven apoptosis. We undertook this study to use the Fas and FasL mutant mice to ask whether some of the apoptosis known to occur after other stimuli, such as ionizing radiation, might also be mediated in part by Fas/FasL interactions. Apoptosis after exposure to γ-radiation occurs rapidly and is mediated both by nuclear and membrane events (10). Because lpr mice are deficient in surface Fas, their sensitivity to cell death induced by γ-irradiation could be compared with that of normal mice to determine the participation of Fas in this phenomenon.
MATERIALS AND METHODS
Mice.
C57BL/6 (B6), C57BL/6-lpr/lpr (B6/lpr), and C57BL/6-gld/gld (B6/gld) mice were bred in our colony. The B6/lpr and B6/gld strains were originally obtained from The Jackson Laboratory. By 5 months of age, the B6/lpr mice develop lymphadenopathy, splenomegaly, and autoantibodies to chromatin and to IgG (11). Mice used in the experiments were 5–8 months old, except for the Fas expression studies in which 2- to 8-month-old animals were used and the Fas–Fc blocking experiments in which 3- to 4-month-old animals were used.
Irradiation-Induced Apoptosis.
Immediately prior to removal of spleen cells for culture, mice were exposed to 150 rad of whole body irradiation from a 137Cs source. In some experiments, cell suspensions were irradiated in 15 cc or 50 cc conical test tubes.
Heat Shock-Induced Apoptosis.
Spleen cells were harvested and washed several times and maintained for 30 min at 43°C in a shaking water bath (12). Temperature was constantly monitored and cells were shaken during this treatment to ensure uniform heating. The cells were then cultured at 37°C after the hyperthermic exposure and examined after 18 h.
Preparation of Cell Suspensions.
Spleen cell suspensions were made by mashing the splenic capsule between frosted ends of glass slides and washing twice with Hanks’ balanced salt solution containing 15 mM Hepes, 0.1% l-glutamine, calcium and magnesium, and 3% fetal calf serum (HBSS complete) (University of North Carolina Cancer Center Tissue Culture Facility, Chapel Hill). Erythrocytes were lysed with ammonium chloride for 5 min at 4°C. In most experiments, three spleens were pooled per group.
Cell Culture.
Cells were cultured for 18 h at 37°C in a 5% CO2/95% air humidified atmosphere in 24-well plates at a concentration of 2 × 106/ml medium (RPMI 1640 medium/10% fetal calf serum/15 mM Hepes/100 units/ml penicillin/100 μg/ml streptomycin/2 mM l-glutamine/1 mM sodium pyruvate/nonessential amino acids). To some cultures Fas–Fc, a fusion protein composed of the extracellular domain of Fas linked to the Fc region of human IgG1 (13), or a purified human IgG1 myeloma protein was added. As a control for Fas-mediated killing, anti-Fas (Jo2, hamster IgG; PharMingen) was added to short-term cultures of thymocytes. To some cultures, dexamethasone at a concentration of 10−6 M was added to induce apoptosis.
Immunofluorescence Staining.
Fluoresceinated anti-IgM (Il/41, rat IgG2a), anti-CD4 (RM4–5, rat IgG2a), anti-CD8 (53–6.7, rat IgG2a), anti-Fas (Jo2, hamster IgG), anti-2,4,6-trinitrophenyl (TNP) (UC8–4B3, hamster IgG), and anti-Thy1.2 (53–2.1, rat IgG2a) were obtained from PharMingen. For staining, samples were washed twice in complete HBSS and labeled with fluoresceinated antibodies as described (14). Briefly, fluoresceinated antibodies were added to 1 × 106 cells for 30 min at 4°C in 96-well microtiter plates. The cells were washed once with complete HBSS followed by two washes in PBS with 0.1% NaN3. To permeabilize cell membranes for DNA staining, 3 ml of unlabeled 70% ethanol (per sample of 1 × 106 cells) was added to the pelleted cells, which were then vortex mixed, incubated for 1 h at 4°C, and washed twice. For DNA staining (15), 0.1 ml of 1 mg/ml RNase A (Sigma) was added per sample with vortex mixing, followed by 0.2 ml of 100 μg/ml propidium iodide (PI) (Sigma). Cells were incubated for 20 min in the dark at 4°C and were then analyzed with a FACScan (Becton Dickinson) with Cytomation data acquisition and software (Fort Collins, CO) for green and red fluorescence. Anti-Fcγ receptor antibody (2.4G2, mouse IgG2b; American Type Culture Collection) was used in most experiments to block nonspecific staining.
Analysis of Apoptotic Cells.
Apoptotic cells appeared in the <2N DNA peak identified by PI immunofluorescence (16). They could also be distinguished from necrotic cells by analyzing the light scatter profile. Cell death by necrosis resulted in a large decrease in forward-angle light scatter (FSC) and side scatter (SSC), while apoptotic cells showed a smaller decline in FSC and an increase in SSC (17).
Samples were collected in list mode, so that when cell surface phenotypic markers were used, the percentage of cells undergoing apoptosis could be determined by gating on fluorescein isothiocyanate (FITC)-stained cells and subsequently analyzing DNA staining (16). Data were plotted on a 4-decade logarithmic scale, except for DNA staining, which was always plotted linearly. At least 20,000 events were collected per sample in all experiments, and 30,000 events were collected for most experiments.
Statistical Analysis.
Student’s t-test was used to determine the statistical significance of differences between groups.
RESULTS
Lymphocytes from 5- to 8-Month-Old lpr Mice Showed Less Radiation-Induced Apoptosis When Compared with Cells from +/+ Mice.
To test if some of the apoptosis known to occur after ionizing radiation might be mediated by Fas/FasL interactions, we quantitated apoptosis using two-color flow cytometry of splenic lymphocytes from irradiated B6/lpr and B6 mice in a short-term in vitro culture system. Total apoptosis from both strains was substantial after irradiation, yet B6 cells underwent significantly more apoptosis than cells from B6/lpr mice (73% versus 25% apoptotic cells using PI staining). Fig. 1 shows data from a representative experiment. It is evident in the forward angle versus side scatter histograms that the majority of cells in the irradiated lpr sample fell into the live region, while in contrast, the majority of cells in the irradiated B6 sample fell in the apoptotic region. This difference is also reflected in the DNA histograms. In seven experiments, cells from irradiated B6 mice showed significantly less apoptosis than cells from B6/lpr (P = 0.0019, Fig. 2). Because the amount of apoptosis was variable from experiment to experiment, data within a single experiment are paired (Fig. 2).
Figure 1.
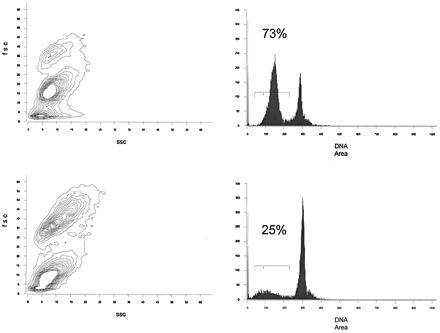
Spleen cells from 8-month-old B6 (Upper) and B6/lpr mice (Lower) were cultured overnight after 150 rads of γ-irradiation. (Left) Forward angle versus side scatter histograms. (Right) DNA histograms. Numbers above bracketed areas indicate the percent of cells with subdiploid DNA content, representing cells undergoing apoptosis. These data are from an experiment representative of seven experiments. Three spleens were pooled in each group per experiment.
Figure 2.
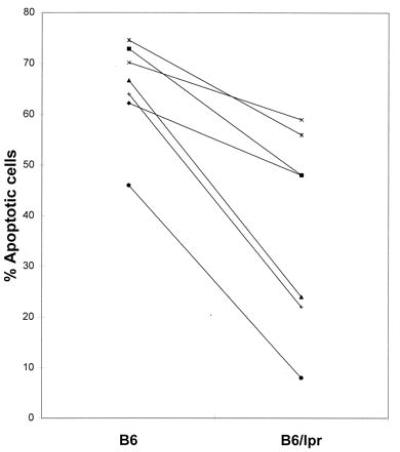
The percent apoptosis determined by PI staining of 5- to 8-month-old B6 (Left) and lpr (Right) irradiated spleen cells that were cultured as in Fig. 1 is shown. Data points represent individual experiments where three spleens were pooled in each group per experiment. Values determined for cells from mice in the same individual experiments are connected with a line. The mean % apoptotic cells from B6 was 68.9 ± 4.4 and 40.1 ± 7.8 for B6/lpr. Numbers after the mean represent the standard error. The difference between the groups was significant to a P value of 0.0019.
T Cells from lpr Mice Showed Markedly Diminished Apoptosis.
Two-color immunofluorescence was utilized to study apoptosis in specific cell populations. Cells were stained for both DNA content and for cell surface markers (Thy1.2, CD4, and IgM). Thy1.2+ cells from lpr mice showed less apoptosis after γ-irradiation compared with T cells from normal mice (Fig. 3A). In multiple experiments, the amount of apoptosis of Thy1.2+ cells after radiation and overnight culture was significantly less in lpr than in B6 cells (Fig. 3B). Data within a single experiment are paired as in Fig. 2 to account for variability between experiments.
Figure 3.
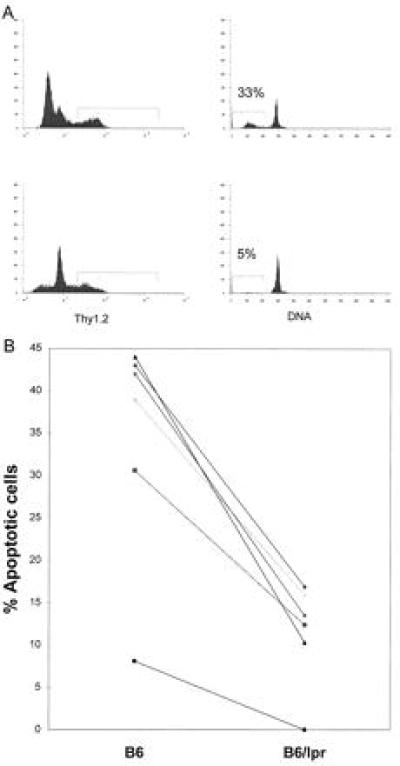
(A) Spleen cells from 8-month-old B6 (Upper) and lpr mice (Lower) were cultured overnight after irradiation and stained with FITC-labeled monoclonal antibody to Thy1.2. (Left) Surface expression of FITC. Positive cells appear under the gates. (Right) DNA content of Thy1.2+ cells. Apoptotic cells appear under the gates to the left of the 2N DNA peak. (B) The percent of Thy1.2+ cells undergoing apoptosis for B6 (Left) and lpr (Right) irradiated spleen cells is shown. Data points represent individual experiments where three spleens were pooled in each group per experiment. Results from individual experiments are connected with a line. The mean % apoptotic T cells from B6 was 34.4 ± 5.6 and 11.5 ± 2.5 for B6/lpr. Numbers after the mean represent the standard error. The difference between the groups was significant to a P value of 0.0015.
Decreased Radiation Sensitivity Was Not Limited to the Abnormal CD4−CD8− T Cells in lpr Mice.
To test whether the diminished radiation-induced apoptosis seen in lpr cells was a property of the abnormal double negative (DN) population (2), lpr T cells were stained for CD4 and the DN T cells excluded from the analysis. CD4+ T cells from lpr mice showed much less apoptosis upon γ-irradiation, indicating that the apoptotic differences seen in lpr cells were not merely due to a special radiation resistance of the DN population (Fig. 4). Variability between experiments is accounted for by pairing data within a single experiment. Parallel results were obtained when the apoptosis differences between CD8+ cells from lpr and B6 mice after radiation and overnight culture were compared (data not shown).
Figure 4.
(A) Spleen cells stained with anti-CD4 from B6 (Left) and lpr mice (Right) were cultured overnight after irradiation and assessed for apoptosis by quantitating DNA content. Data shown represent the DNA content of CD4+ cells from a 7-month-old animal and are from a representative experiment. (B) The percent apoptosis of CD4+ B6 (Left) and lpr (Right) irradiated spleen cells that were cultured as in Fig. 1 is shown. Data points represent individual experiments where three spleens were pooled in each group per experiment. Data from the same experiment are connected with a line. The mean % apoptotic CD4+ T cells from B6 was 19.1 ± 6.3 and 5.4 ± 1.8 for B6/lpr. Numbers after the mean represent the standard error. The difference between the groups was significant to a P value of 0.0383.
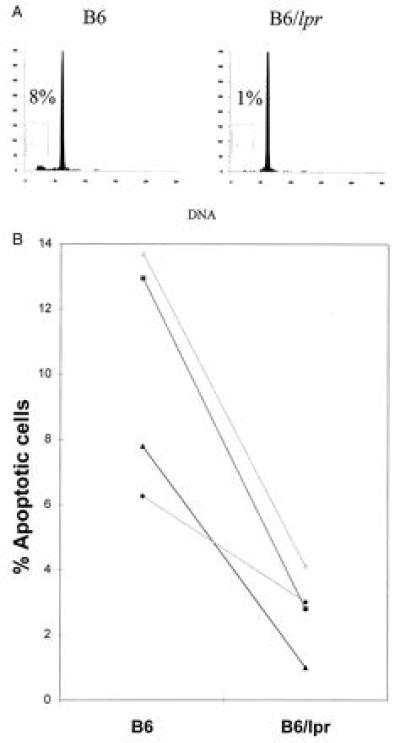
B Cells from lpr Mice Show Decreased Radiation Sensitivity Compared with B6 B Cells.
Because B cells are also known to be relatively radiation sensitive (18), spleen cells were stained with an antibody to surface IgM and apoptosis was quantitated in the positive population after irradiation and overnight culture. Similar to the results obtained after staining with T cell antibodies, lpr B cells showed much less apoptosis upon exposure to ionizing radiation (Fig. 5). Data within a single experiment are paired as in Fig. 2 to account for variability between experiments.
Figure 5.
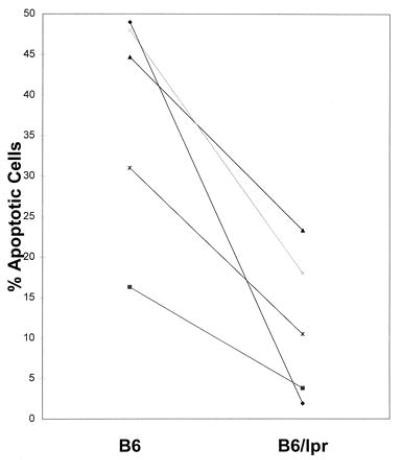
The percent of B cells undergoing apoptosis in B6 (Left) and lpr (Right) irradiated spleen cells that were cultured as in Fig. 1 is shown. Data points represent individual experiments where three spleens were pooled in each group per experiment. Results from individual experiments are connected with a line. The mean % apoptotic B cells from B6 was 37.8 ± 6.3 and 11.5 ± 4.1 for B6/lpr. Numbers after the mean represent the standard error. The difference between the groups was significant to a P value of 0.0111.
In Vitro Irradiation of Spleen Cells Parallels Results of in Vivo Irradiation of Spleen Cells.
To determine if in vitro irradiation of spleen cells would produce the same results as in vivo whole body irradiation of mice, spleen cell suspensions were first prepared and then cells were irradiated. Results obtained paralleled the results of the in vivo experiments (see below).
Fas Expression Is Up-Regulated in Normal Mice After γ-Irradiation.
Our results demonstrated that lpr cells, which are defective in Fas, showed decreased apoptosis after γ-irradiation. It was possible that Fas/FasL interactions, which could take place in the normal but not the lpr cell cultures, were responsible for the consistently greater degree of apoptosis seen in irradiated normal cells compared with those from lpr. We wondered whether irradiation might even increase the expression of Fas on the B6 cells, thus enhancing Fas/FasL apoptosis. We thus quantitated Fas expression on cells from normal B6 mice before and after irradiation. As a positive control, B6 thymocytes were stained with Fas–FITC, and as expected 100% of the cells were positive for Fas.
By immunofluorescence staining, Fas was up-regulated in B6 spleen cells after γ-irradiation (Fig. 6). Fig. 6 Upper shows the FITC histograms for live cells before and after irradiation. Fig. 6 Lower shows FITC histograms for Fas expression on the apoptotic cells. It can be seen in both cell populations that Fas was up-regulated. No positive staining was found on the same cell populations stained with a monoclonal hamster anti-TNP of the same IgG isotype. In 10 experiments, the percent of Fas+ cells increased significantly (P = 0.0002) after γ-irradiation of B6 mice (24.8 ± 5.2 before irradiation compared with 40.0 ± 7.9 after irradiation). Thus, normal B6 spleen cells contain increased numbers of Fas+ cells after irradiation.
Figure 6.
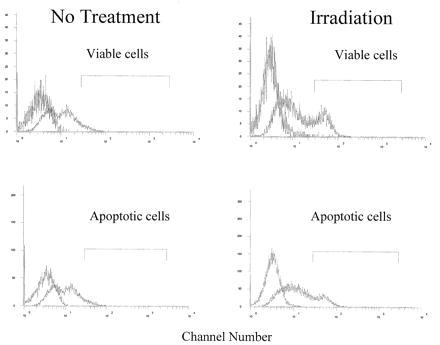
Unirradiated B6 splenocytes (Left) and in vivo irradiated (150 rads) B6 splenocytes (Right) from 5-month-old mice were cultured overnight and stained for Fas using FITC hamster IgG anti-Fas (right fluorescence profile in each histogram). In each histogram, the peak on the far left represents the same cells stained with the isotype control hamster IgG anti-TNP–FITC and was negative on all populations. (Upper) Histograms represent the FITC profile of viable cells. (Lower) FITC profiles for apoptotic cells from the same B6 splenocytes. Apoptotic and viable cells were identified by their forward angle versus side scatter profile. These histograms are representative of 10 experiments. Anti-Fc receptor antibody was used to block nonspecific binding.
In additional experiments, lymphocytes from +/+ and lpr mice were treated with dexamethasone (Dex) and apoptosis and Fas expression was measured. In contrast to γ-irradiation, Dex did not increase Fas expression on B6 cells after overnight culture nor was apoptosis significantly diminished in cells from lpr mice after Dex culture compared with cells from normal mice (data not shown).
lpr Cells Show Diminished Apoptosis in Response to Heat Shock Treatment.
To determine if the relative resistance to injury-induced apoptosis of lpr lymphoid cells was unique to γ-irradiation, cells were exposed to heat shock. Heat shock-induced apoptosis was also impaired in lymphocytes from lpr mice compared with normal cells (Fig. 7).
Figure 7.
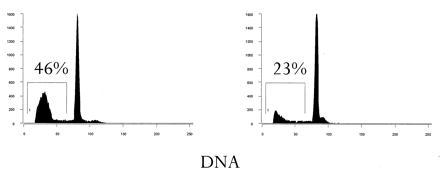
The DNA profiles of cells from 6 month old B6 (Left) and lpr (Right) after heat shock is shown. This experiment was repeated six times with similar results.
Lymphocytes from gld Mice Paralleled the Results Found with Cells from lpr Mice.
To determine if cells from Fas ligand mutant gld mice were also relatively resistant to irradiation-induced apoptosis, spleen cells from 8-month-old gld mice were put into short-term in vitro culture. Apoptosis was quantitated after γ-irradiation as described above. Gld cells showed less radiation-induced apoptosis when compared with cells from +/+ mice and thus paralleled the results found with lpr cells (Fig. 8).
Figure 8.
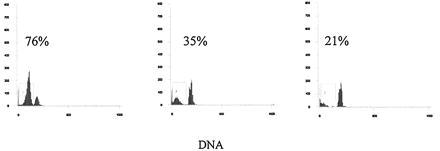
Spleen cells from 8-month-old B6 (Left), B6/lpr (Center), and B6/gld (Right) were cultured overnight after 150 rads of γ-irradiation. DNA histograms are shown and numbers above bracketed areas indicate the percent of cells undergoing apoptosis. These experimental data are representative of four separate experiments.
Irradiation-Induced Apoptosis Was Blocked with High Concentrations of Fas–Fc Fusion Protein.
To assess more directly the role of Fas in radiation-induced apoptosis, a Fas–Fc fusion protein (13) was added to irradiated normal cells. This Fas–human IgG1 Fc fusion protein was successful in inhibiting Fas-mediated apoptosis of thymocytes (Fig. 9). When irradiated spleen cells were incubated with Fas–Fc, apoptosis was inhibited by ≈60% (Fig. 10); a human IgG1 myeloma protein had no effect (Fig. 10). It was necessary to use concentrations of Fas–Fc over 100 μg/ml, as lower concentrations failed to block radiation-induced apoptosis but blocked anti-Fas apoptosis of thymocytes (data not shown). This suggested that the Fas/FasL interactions involved in radiation-induced apoptosis were of high avidity and thus difficult to block.
Figure 9.
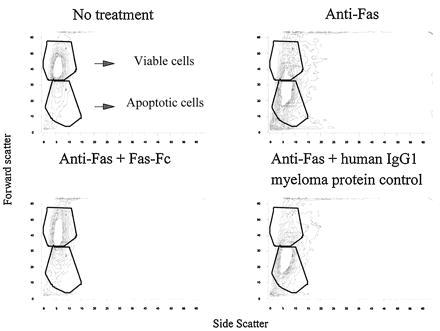
Thymocytes from three pooled 3- to 4-month-old B6 mice were cultured overnight (Upper Left), incubated with 1 μg/ml of anti-Fas during culture (Upper Right), incubated with 1 μg/ml of anti-Fas with the addition of 100 μg/ml of Fas-Fc during culture (Lower Left), or incubated with 1 μg/ml of anti-Fas with the addition of 100 μg/ml of a purified human IgG1 myeloma protein during culture (Lower Right). Viable and apoptotic cells are depicted in the outlined regions as labeled. Viable and apoptotic cells in each panel are as follows: no treatment, 63% viable and 21% apoptotic; anti-Fas treatment, 24% viable and 44% apoptotic; anti-Fas plus Fas–Fc, 57% viable and 22% apoptotic; and anti-Fas plus human IgG1 myeloma protein control, 22% viable and 51% apoptotic.
Figure 10.
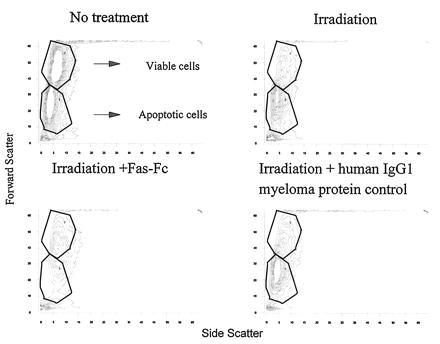
Spleen cells from 3 month-old B6 mice were cultured overnight (Upper Left), irradiated with 150 rads and cultured overnight (Upper Right), irradiated and cultured overnight with 200 μg/ml Fas–Fc (Lower Right), or irradiated and cultured overnight with 200 μg/ml of a purified human IgG1 myeloma protein (Lower Right). Viable and apoptotic cells are depicted in the outlined regions as labeled. Viable and apoptotic cells in each panel are as follows: no treatment, 45% viable and 36% apoptotic; irradiated, 25% viable and 50% apoptotic; irradiated plus Fas–Fc, 35% viable and 20% apoptotic; and irradiated plus human IgG1 myeloma protein control, 26% viable and 51% apoptotic.
DISCUSSION
We have observed that cells from Fas-deficient lpr mice are relatively resistant to the apoptosis induced by ionizing radiation. This resistance was noted for cells bearing markers associated with normal B and T cells, as well as for cells expressing the aberrant “double negative” phenotype associated with the lpr lymphoproliferative state, making it unlikely that the radiation resistance was due to the abnormal differentiation state of the abnormal lpr T cells. Because the lpr mice differed from controls only in their lack of Fas expression, these observations raised the possibility that Fas itself may participate in the apoptosis induced by ionizing radiation.
Ionizing radiation induces apoptosis both by direct effects on the nucleus and through effects on plasma membranes, probably through hydrolysis of sphingomyelin to ceramide (10). As Fas has been shown to signal at least in part via this pathway (19–21), it seemed possible that Fas-deficient lpr cells might be lacking in signaling intermediaries whose levels might be linked to the normal levels of surface Fas. A more intriguing possibility is that a portion of radiation-induced apoptosis might actually be mediated by Fas, and that therefore radiation would induce less apoptosis in lpr. This hypothesis is supported by our finding that surface Fas expression increases on normal lymphocytes exposed to radiation, potentially allowing their destruction by FasL-bearing T cells, and that radiation-induced apoptosis can be blocked in part with high concentrations of a Fas–Fc fusion protein. The Fas-mediated effects of radiation need not only be through increased surface levels of Fas, but might also influence overall functioning of the intracellular arm of this pathway (see below).
It has already been established that Fas expression is induced by lymphocyte activation, both T (22) and B (23), and it has been proposed that the increase in Fas expression may provide a means whereby activated cells may be eliminated once they have served their function in the immune response (24). Our observations suggest an additional function of inducible Fas. Surface expression of this receptor increases after irradiation, and probably also after heat shock. It may be that multiple sources of cellular injury lead to increased Fas expression, and that increased Fas may serve as a marker which targets injured cells for destruction by FasL-bearing T lymphocytes. In effect, induction of Fas expression may serve as a means whereby the immune system can perform a sort of immunological surveillance to identify and eliminate not only activated cells, but also cells that have undergone injury and that require induction of apoptotic death and subsequent elimination. This mechanism would supplement the direct apoptosis-inducing effects of radiation, heat shock, and other forms of physical injury, as well as other pathways which induce apoptosis, such as tumor necrosis factor α.
The proposed role for FasL/Fas interactions in inducing death of damaged cells raises questions about the nature of the interaction between FasL and Fas-bearing cells. In general, these cellular interactions are believed to be cognate, and to be mediated by TCR recognition of major histocompatibility antigen complexes on the surface of Fas-bearing cells (7). It is possible that the interactions we propose also require such TCR-mediated interactions, or that the death of Fas-expressing cells does not require contact with antigen-specific T cells. In this regard, recent data indicate that the level of Fas expression is only one factor regulating the susceptibility to Fas-mediated apoptosis (24). Exposure to radiation or to other forms of stress may lead to apoptosis not only by increasing surface Fas, but also by affecting intracellular signaling pathways that facilitate Fas-mediated killing.
The list of functions for Fas/FasL continues to grow. It includes, beside mediation of TCR-mediated killing, T and B cell tolerance, and immunoregulation (24). These observations provide evidence that this system may be involved in policing against injured cells, and lead to testable notions about the nature and significance of this phenomenon.
Acknowledgments
We thank Mrs. Sylvia Craven for expert technical assistance and Dr. Roberto Caricchio for help with the graphics. We also thank Drs. Alan Zajac and Jeff Frelinger for their generous gift of the Fas–Fc fusion protein. This work was supported by Public Health Service Grants AR42573 and AR33887. E.A.R. is a fellow of the Arthritis Foundation.
ABBREVIATIONS
- FasL
Fas ligand
- TCR
T cell antigen receptor
- HBSS
Hanks’ balanced salt solution
- TNP
2,4,6-trinitrophenyl
- PI
propidium iodide
- FITC
fluorescein isothiocyanate
References
- 1.Cohen P L, Eisenberg R A. Annu Rev Immunol. 1991;9:243–269. doi: 10.1146/annurev.iy.09.040191.001331. [DOI] [PubMed] [Google Scholar]
- 2.Watanabe-Fukunaga R, Brannan C I, Copeland N G, Jenkins N A, Nagata S. Nature (London) 1992;356:314–317. doi: 10.1038/356314a0. [DOI] [PubMed] [Google Scholar]
- 3.Singer G G, Abbas A K. Immunity. 1994;1:365–371. doi: 10.1016/1074-7613(94)90067-1. [DOI] [PubMed] [Google Scholar]
- 4.Russell J H, Rush B, Weaver C, Wang R. Proc Natl Acad Sci USA. 1993;90:4409–4413. doi: 10.1073/pnas.90.10.4409. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Russell J H, Wang R. Eur J Immunol. 1993;23:2379–2382. doi: 10.1002/eji.1830230951. [DOI] [PubMed] [Google Scholar]
- 6.Scott D E, Kisch W J, Steinberg A D. J Immunol. 1993;150:664–672. [PubMed] [Google Scholar]
- 7.Bossu P, Singer G G, Andres P, Ettinger R, Marshak-Rothstein A, Abbas A K. J Immunol. 1993;151:7233–7239. [PubMed] [Google Scholar]
- 8.Garrone P, Neidhardt E, Garcia E, Gailbert L, van Kooten C, Banchereau J. J Exp Med. 1995;182:1265–1273. doi: 10.1084/jem.182.5.1265. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Lagresle C, Mondiere P, Bella C, Krammer P, Defrance T. J Exp Med. 1996;183:1377–1388. doi: 10.1084/jem.183.4.1377. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Haimovitz-Friedman A, Kan C, Ehleiter D, Persaud R G, McLoughlin M, Fuks Z, Kolesnick R N. J Exp Med. 1994;180:525–535. doi: 10.1084/jem.180.2.525. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Sobel E S, Katagiri T, Katagiri K, Morris S C, Cohen P L, Eisenberg R A. J Exp Med. 1992;173:1441–1449. doi: 10.1084/jem.173.6.1441. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Mogil R J, Shi Y, Bissonnette R P, Bromley P, Yamaguchi I, Green D R. J Immunol. 1994;152:1674–2683. [PubMed] [Google Scholar]
- 13.Ju S-T, Panka D J, Cui H, Ettinger R, El-Khatib M, Sherr D H, Stanger B Z, Marshak-Rothstein A. Nature (London) 1995;373:444–448. doi: 10.1038/373444a0. [DOI] [PubMed] [Google Scholar]
- 14.Reap E A, Piecyk M L, Oliver A, Sobel E S, Waldschmidt T, Cohen P L, Eisenberg R A. Clin Immunol Immunopathol. 1996;78:21–29. doi: 10.1006/clin.1996.0004. [DOI] [PubMed] [Google Scholar]
- 15.Reap E A, Leslie D, Abrahams M, Eisenberg R A, Cohen P L. J Immunol. 1995;154:936–943. [PubMed] [Google Scholar]
- 16.Darzynkewicz Z, Bruno S, DelBino G, Gurizyca W, Hotz M A, Lassota P, Traganos F. Cytometry. 1992;13:795–805. doi: 10.1002/cyto.990130802. [DOI] [PubMed] [Google Scholar]
- 17.Illera V A, Perandones C E, Stunz L L, Mower D A, Jr, Ashman R F. J Immunol. 1993;151:2965–2973. [PubMed] [Google Scholar]
- 18.Woodland R T, Schmidt M R, Riggs J E, Korsmeyer S J, Lussier A M, Gravel K A. J Immunol. 1995;155:3453–3463. [PubMed] [Google Scholar]
- 19.Cifone M G, DeMaria R, Roncaioli P, Rippo M R, Azuma M, Lanier L L, Santoni A, Testi R. J Exp Med. 1994;177:1547–1552. doi: 10.1084/jem.180.4.1547. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Gulbins E, Bissonnette R, Mahboubi A, Martin S, Nishioka W, Brunner T, Baier G, Baier-Bitterlick G, Byrd C, Lang F, Kolesnick R, Altman A, Green D. Immunity. 1995;2:341–351. doi: 10.1016/1074-7613(95)90142-6. [DOI] [PubMed] [Google Scholar]
- 21.Tepper C G, Jayadev S, Liu B, Bielawska A, Wolff R, Yonehara S, Hannun Y, Seldin M F. Proc Natl Acad Sci USA. 1995;92:8443–8447. doi: 10.1073/pnas.92.18.8443. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Drappa J, Brot N, Elkon K B. Proc Natl Acad Sci USA. 1993;90:10340–10346. doi: 10.1073/pnas.90.21.10340. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Onel K B, Tucek-Szabo C L, Ashany D, Lacy E, Nikolic-Zugic J, Elkon K B. Eur J Immunol. 1995;25:2940–2947. doi: 10.1002/eji.1830251034. [DOI] [PubMed] [Google Scholar]
- 24.Cohen P L, Eisenberg R A. In: Apoptosis and the Immune Response. Gregory C, editor. New York: Wiley; 1995. pp. 169–186. [Google Scholar]