

Abstract
The cytokines interleukin (IL)-4 and IL-13 play a critical role in inducing Cɛ germline transcripts and IgE isotype switching in human B cells. The IL-4 receptor (IL-4R) in B cells is composed of two chains, the IL-4-binding IL-4Rα chain, which is shared with the IL-13R, and the IL-2Rγ (γc) chain, which is shared with IL-7R, IL-9R, and IL-15R. IL-4 induces Cɛ germline transcripts and IgE isotype switching in B cells from patients with γc chain deficiency. Induction of Cɛ germline transcripts by IL-4 in B cells that lack the γc chain may involve signaling via the IL-13R. Alternatively, the IL-4Rα chain may transduce intracellular signals that lead to Cɛ gene transcription independently of its association with other chains. We show that ligand-induced homodimerization of chimeric surface receptors consisting of the extracellular and transmembrane domains of the erythropoietin receptor and of the intracellular domain of IL-4Rα induces Janus kinase 1 (Jak1) activation, STAT6 activation, and Cɛ germline transcripts in human B cell line BJAB. Disruption of the Jak1-binding proline-rich Box1 region of IL-4Rα abolished signaling by this chimeric receptor. Furthermore, B cells transfected with a chimeric CD8α/IL-4Rα receptor, which is expressed on the cell surface as a homodimer, constitutively expressed Cɛ germline transcripts. These results suggest that homodimerization of the IL-4Rα chain is sufficient to transduce Jak1-dependent intracellular signals that lead to IgE isotype switching.
IgE synthesis by B cells requires two signals, the cytokine interleukin 4 (IL-4) that induces transcription of the Cɛ gene, targeting it for switch recombination, and interaction of the B cell surface antigen CD40 with its ligand (CD40L) expressed on activated T cells, which results in deletional switch recombination (1). In humans, the cytokine IL-13 duplicates the function of IL-4 in inducing IgE isotype switching (2).
The IL-4 receptor (IL-4R) on lymphoid cells is composed of two chains, the IL-4-binding 140-kDa IL-4Rα chain that is shared with the IL-13R (3), and the IL-2Rγ (γc) chain (4) that is also shared with IL-7R, IL-9R, and IL-15R (5) and that is mutated in boys with X-linked severe combined immunodeficiency (X-SCID) (6).
IL-4Rα and γc respectively associate with the Janus kinases Jak1 and Jak3 (7). Ligand-induced heterodimerization of IL-4Rα and γc chains in lymphoid cells induces phosphorylation and activation of Jak1 and Jak3 and tyrosine phosphorylation of the IL-4Rα chain (8). Phosphorylated tyrosine residues of IL-4Rα chain serve as anchor for the SH2 domain of the signal transducer and activator of transcription STAT6, which in turn gets phosphorylated (9). Phosphorylated STAT6 form homodimers that translocate to the nucleus where they bind to DNA sequences TTCNNN(N)GAA in the promoter region of IL-4 responsive genes, including Cɛ (10, 11).
The IL-13R complex is supposed to consist of two chains, an IL-13-binding IL-13Rα chain (12, 13) and the IL-4Rα chain (3). γc chain is not a component of IL-13R (14). Although IL-13 activates STAT6 and induces Cɛ germline transcripts (15), it does not activate Jak3.
The role of individual IL-4R chains in the induction of Cɛ gene transcription is not very well defined. It was recently shown that IL-4 induces Cɛ germline transcripts and IgE isotype switching in B cells from patients with X-SCID and mutated γc chain (15, 16). Induction of Cɛ gene transcription by IL-4 in B cells with γc chain-deficiency may involve signaling via the IL-4Rα-containing IL-13 receptor. Alternatively, the IL-4Rα chain by itself may transduce intracellular signals that lead to Cɛ gene transcription. In this study, we show that homodimerization of the intracellular domain of the human IL-4Rα chain induces Cɛ germline transcripts. This was accompanied by Jak1 activation and induction of nuclear STAT6 DNA binding activity and was abolished by disruption of the Jak1-binding proline-rich Box1 region of IL-4Rα.
MATERIALS AND METHODS
Generation of Erythropoietin Receptor (EpoR)/IL-4Rα and CD8α/IL-4Rα Chain Chimeras.
To generate a chimeric cDNA encoding the extracellular (EC) and the transmembrane (TM) domains of murine EpoR (muEpoR) fused to the intracellular (IC) domain of the human IL-4Rα chain, overlapping PCR fragments encompassing the appropriate regions of EpoR and IL-4Rα were annealed in a two-step overlap extension PCR as previously described (17). MuEpoR cDNA was a gift of B. Mathey-Prevot (Dana–Farber Cancer Institute, Boston). Human IL-4R cDNA was a gift of P. Leder and D. Seldin (Harvard Medical School). The PCR products that consist of the EpoR-EC and TM domains fused to the IL-4Rα IC domain, EpoREC+TM/IL-4RαIC, were gel purified, cloned into the PCR II vector (Invitrogen), sequenced, and subcloned into the mammalian expression vector pcDNA3 (Invitrogen) containing the neomycin resistance gene. As a control vector, full-length EpoR cDNA was subcloned into pcDNA3.
A chimera consisting of the EC and TM domains of human CD8α and the IC domain of IL-4Rα (CD8EC+TM/IL-4RαIC) was also constructed by the overlap extension PCR using human CD8α cDNA (a gift of L. Clayton, Dana–Farber Cancer Institute) and verified by sequencing. The chimeric CD8/IL-4Rα cDNA was cloned into the mammalian expression vector pCEP4 (Invitrogen) containing the hygromycin resistance gene. As a control, full-length CD8α cDNA was subcloned into pCEP4.
Cell Line and Transfections.
The Epstein–Barr virus-negative human Burkitt lymphoma B cell line BJAB was a gift of E. Kieff (Brigham and Women’s Hospital). BJAB cells were maintained in RPMI medium 1640 supplemented with l-glutamine and 10% fetal bovine serum. Transfection was performed by electroporation, followed 24 hr later by selection with 1.5 mg/ml of G418 (GIBCO/BRL) or 400 μg/ml of hygromycin B (Boehringer Mannheim). Resistant clones were further selected for EpoR surface expression by fluorescence activated cell sorting.
Fluorescence-Activated Cell Sorter (FACS) Analysis.
Surface expression of EpoR was assessed using a rabbit anti-human EpoR antiserum that crossreacts with murine EpoR (a gift of A. D’Andrea, Dana–Farber Cancer Institute), followed by biotin-conjugated sheep anti-rabbit IgG F(ab)′2 antibody (Sigma) and streptavidin-phycoerythrin (Becton Dickinson). Surface CD8 expression was assessed using phycoerythrin-conjugated mouse-anti-human CD8α mAb SK1 (Becton Dickinson). Normal rabbit serum (NRS) or phycoerythrin-conjugated mouse IgG1 was used as a control, respectively. Cells were analyzed by FACSCalibur (Becton Dickinson).
Northern Blot Analysis.
Total RNA was extracted from the cells using TRIzol reagent as described by the manufacturer (GIBCO/BRL) and subjected to Northern blot analysis using 32P-labeled Cɛ probe consisting of a BamHI fragment of genomic DNA that contains all four Cɛ exons, as described (18).
Electrophoretic Mobility-Shift Assay.
Nuclear extracts were prepared and electrophoretic mobility shift assay was performed as previously described (19). The following oligonucleotides were used in electrophoretic mobility shift assay: the interferon gamma-activated sequences (GAS)-like sequence upstream of the human Iɛ exon (Iɛ probe): TCAATCGACTTCCCAAGAACAGAGAGA (STAT site inverted repeats underlined); a mutant Iɛ oligonucleotide (mutant competitor): TCAATCGACGGTCCAAGAACAGAGAGA (mutated nucleotides are underlined); a consensus activating protein 1 (AP-1) sequence (irrelevant competitor): CGCTTGATGAGTCAGCCG. For supershift, nuclear extracts were preincubated for 30 min with 1 μl (0.1 μg) of rabbit anti-human STAT6 antibody (Santa Cruz Biotechnology)
Immunoprecipitation and Western Blotting.
BJAB cells were serum-starved in RPMI 1640 medium containing 0% fetal bovine serum for 6 h, then treated with 300 units/ml of Epo (Amgen) or with 1,200 units/ml of IL-4 (a gift from J. DeVries, DNAX) for the indicated lengths of time. Cell lysates were prepared as previously described (20). The lysates were precleared overnight with NRS coupled to protein G Sepharose beads, then immunoprecipitated for 3 hr with either NRS or with the appropriate antibodies to Jak kinases coupled to protein G Sepharose beads.
The immunoprecipitates were washed five times in lysis buffer and eluted by boiling with SDS-sample buffer. Proteins were resolved by SDS/PAGE and transferred to nitrocellulose membranes. Blots were probed with the desired antibody and developed with enhanced chemiluminescence (Amersham). Mouse anti-human phosphotyrosine mAb 4G10, rabbit anti-Jak1 and anti-Jak2 antibodies were purchased from Upstate Biotechnology (Lake Placid, NY). Rabbit anti-Jak3 antibody and rabbit anti-Tyk2 antibody were from Santa Cruz Biotechnology. Goat anti-mouse IgG linked to horseradish peroxidase was from Bio-Rad. Protein A linked to horseradish peroxidase was from Amersham.
Construction of EpoR/IL-4Rα Box1 Mutant.
PCR-mediated mutagenesis was used to introduce two deletions, 8 and 3 aa each, in the proline-rich Box1 region of the IL-4Rα in the chimeric EpoR/IL-4Rα by the Stratagene Quick Change kit (Stratagene). Box1 mutant 1 (Box1 mut 1) and Box1 mutant 2 (Box1 mut 2) of the chimeric EpoR/IL-4Rα lacked the sequence 237 WWDQIPNP 244 and 242 PNP 244, respectively. The amino acid number denote the amino acid position of the mature IL-4Rα protein. In each case, the entire DNA sequence of the chimeric construct was verified by sequencing.
RESULTS
Homodimerization of EpoR/IL-4Rα Chimeras by Epo Induces Expression of Cɛ Gene Transcripts in BJAB Cells.
Fig. 1A shows that there was no detectable endogenous EpoR expression on BJAB cells transfected with empty pcDNA3 vector. In contrast, EpoR was readily detected on the surface of BJAB cells transfected with the native muEpoR construct as well as on cells transfected with the chimeric EpoR/IL-4Rα construct. The intensity of EpoR staining on cells expressing EpoR and on cells expressing EpoR/IL-4Rα chimeric protein was roughly equivalent.
Figure 1.
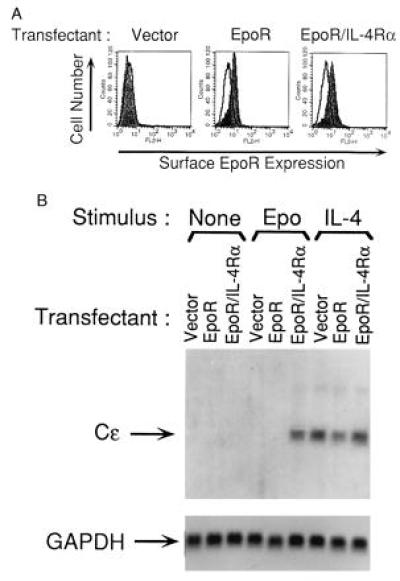
FACS analysis of surface EpoR expression and induction of Cɛ germline transcripts in BJAB transfectants. (A) Surface EpoR expression: BJAB cells transfected with pcDNA3 vector alone, EpoR, or EpoR/IL-4Rα constructs were subjected to G418 selection and fluorescence activated cell sorting, then analyzed for murine EpoR surface expression by FACS. Background fluorescence was determined by staining with normal rabbit serum (—). Filled fields show staining with rabbit anti-EpoR antiserum. Similar results were obtained in two independent transfections. (B) Expression of Cɛ germline transcripts: Transfectants were left unstimulated (None), or stimulated with Epo (50 units/ml) or IL-4 (200 units/ml) for five days then assessed for expression of Cɛ germline transcripts by Northern blot analysis. Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) mRNA expression was used as standard for equal loading. Similar results were obtained in three independent experiments.
We therefore examined whether homodimerization of the EpoR/IL-4Rα chimeric receptor by Epo in BJAB cells mimics IL-4 signaling via native IL-4R in causing Cɛ gene transcription. We used recombinant human Epo in our studies because it is readily available and has been shown to bind to muEpoR and to signal via muEpoR (21). Fig. 1B shows that incubation with IL-4 for five days induced Cɛ gene transcripts in BJAB cells transfected with empty vector, with EpoR, or with EpoR/IL-4Rα (lanes 7–9), whereas no Cɛ gene transcripts were detected in cells incubated with medium alone (lanes 1–3). Epo induced Cɛ gene transcripts only in BJAB cells that were transfected with EpoR/IL-4Rα chimeric receptor (lane 6), and not in BJAB cells transfected with empty vector or with full-length EpoR (lanes 4 and 5). These results strongly suggest that homodimerization of the IL-4Rα chain is sufficient to transduce an intracellular signal that results in Cɛ gene transcription.
Expression of CD8/IL-4Rα Homodimers in BJAB Cells Results in Constitutive Expression of Cɛ Gene Transcripts.
CD8α is expressed at the cell surface as a homodimer as long as it has its own EC and TM domains (22, 23). To confirm that homodimerization of IL-4Rα is sufficient to induce Cɛ gene transcripts in B cells, we examined Cɛ germline transcripts in BJAB cells transfected with a CD8EC+TM/IL-4RαIC chimeric construct, or with a full-length CD8, or with a vector alone. The cells were grown in hygromycin containing medium and were examined from day 13 on for surface CD8 expression by FACS analysis and for expression of Cɛ gene transcripts by Northern blotting.
Fig. 2A shows that 13 days after transfection, CD8 was expressed on the surface of BJAB cells transfected with the native CD8 construct as well as on the surface of BJAB cells transfected with the chimeric CD8/IL-4Rα construct. Subsequent to day 13, CD8 surface expression progressively diminished until the end of the experiment on day 21 (data not shown).
Figure 2.
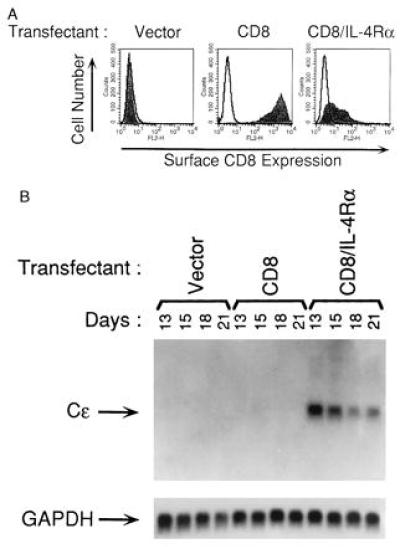
FACS analysis of surface CD8 expression and induction of Cɛ germline transcripts in BJAB transfectants. (A) Surface CD8 expression: BJAB cells transfected with pCEP4 vector alone, CD8, or CD8/IL-4Rα constructs and selected in hygromycin were examined 13 days after transfection for surface expression of human CD8 by FACS. Background fluorescence was determined by staining with isotype control antibody (—). Filled fields show staining with anti-CD8 mAb. Similar results were obtained in two independent transfections. (B) Expression of Cɛ germline transcripts: expression of Cɛ germline transcripts and of glyceraldehyde-3-phosphate dehydrogenase (GAPDH) were examined by Northern blot analysis on the indicated days after transfection. Similar results were obtained in three independent experiments.
Fig. 2B shows that Cɛ gene transcripts were constitutively expressed on day 13 in BJAB cells that expressed the CD8/IL-4Rα chimeric receptor. Expression of the Cɛ gene transcripts in these cells persisted at days 15, 18, and 21, although its level progressively diminished with time. Cɛ gene transcripts were not detected at any time in BJAB cells transfected with empty vector or with the full-length CD8α construct, although both transfectants expressed Cɛ gene transcripts following treatment with IL-4 for five days (data not shown). These data support the notion that homodimerization of the IL-4Rα chain is sufficient for the induction of Cɛ gene transcripts.
Homodimerization of IL-4Rα Chains Induces STAT6 Activation.
We examined whether homodimerization of the IL-4Rα chain induced nuclear STAT6 activity. Fig. 3A shows that IL-4 induced a nuclear complex in BJAB cells transfected with empty vector, EpoR and EpoR/IL-4Rα (lanes 7–9) whereas no nuclear complex was detected in cells incubated with medium alone (lanes 1–3). Epo induced a nuclear complex only in BJAB cells that were transfected with EpoR/IL-4Rα (lane 6) and not in BJAB cells transfected with EpoR or empty vector. The nuclear complex induced in BJAB transfected with EpoR/IL-4Rα by Epo comigrated with the nuclear complex induced in BJAB cells by IL-4 signaling via the native lL-4R, suggesting that it contained STAT6.
Figure 3.
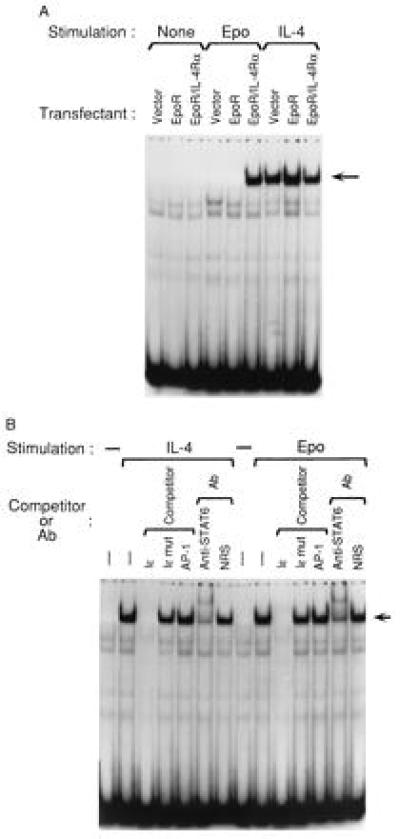
Induction of STAT6 nuclear binding activity to Iɛ oligonucleotide. (A) Electrophoretic mobility-shift assay. BJAB cells transfected with pcDNA3 vector alone, EpoR, or EpoR/IL-4Rα constructs were left unstimulated (None), or were stimulated for 30 min with Epo (50 units/ml), or IL-4 (200 units/ml). The cells were lysed and nuclear extracts were incubated with 32P-labeled Iɛ oligonucleotide, then subjected to PAGE. (B) Specificity and identity of the Iɛ oligonucleotide binding nuclear complex. Nuclear extracts from EpoR/IL-4Rα transfectants left unstimulated or stimulated for 30 min with Epo (50 units/ml) or IL-4 (200 units/ml) were incubated with 32P-labeled Iɛ oligonucleotide and 100-fold molar excess of the following cold competitors: unlabeled Iɛ (Iɛ), Iɛ mutant (Iɛ mut), and an AP-1 consensus sequence (AP-1). For supershift assay, nuclear extracts were pre-incubated with rabbit anti-STAT6 antiserum or with NRS. Similar results were obtained in three independent experiments.
Fig. 3B shows that the nuclear complex induced by Epo in BJAB cells expressing EpoR/IL-4Rα chimeric receptors is specific and, like the nuclear complex induced by IL-4, contains STAT6. Both the IL-4-induced complex and the Epo-induced complex (lanes 2 and 9) were competed out by a 100-fold excess of unlabeled self oligonucleotide (lanes 3 and 10) but not by 100 fold excess of cold Iɛ mutant oligonucleotide (lanes 4 and 11) nor by an irrelevant oligonucleotide containing the AP-1 consensus sequence (lanes 5 and 12). More importantly, both the IL-4-induced complex and the Epo-induced complex were disrupted by preincubation of the nuclear extracts with rabbit antibody to STAT6 protein (lanes 6 and 13) and not with control NRS (lanes 7 and 14). Taken together, these data indicate that homodimerization of the IL-4Rα chain induces STAT6 nuclear activity.
Homodimerization of IL-4Rα Chains Induces Jak1 Activation.
We examined whether homodimerization of IL-4Rα chain by itself results in tyrosine phosphorylation of the IL-4Rα-associated Jak1, in the absence of recruitment of the γc chain and of its associated Jak3.
Fig. 4A shows that engagement of the native IL-4R complex by IL-4 results in tyrosine phosphorylation of Jak3. In contrast, homodimerization of the chimeric EpoR/IL-4Rα receptors resulted in no detectable tyrosine phosphorylation of Jak3 even up to 30 min. following addition of Epo. Fig. 4B shows that tyrosine phosphorylation of Jak1 was readily detectable following engagement of the native IL-4R complex by IL-4 (lane 3). More importantly, homodimerization of the chimeric EpoR/IL-4Rα receptors by Epo also resulted in tyrosine phosphorylation of Jak1 (lane 2). Homodimerization of the chimeric EpoR/IL-4Rα receptors did not lead to detectable tyrosine phosphorylation of Jak2, or Tyk2 (data not shown). These results strongly suggest that homodimerization of IL-4Rα chains results in the activation of Jak1, with no detectable activation of other known Jak kinases.
Figure 4.
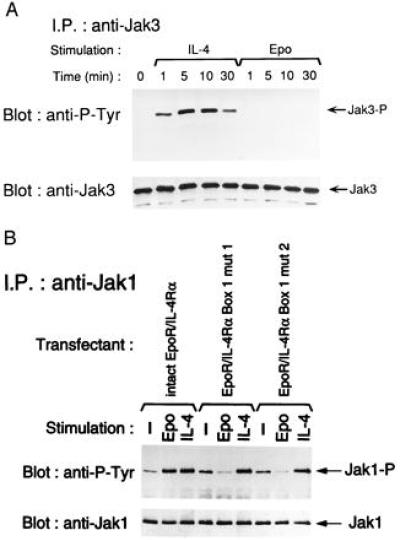
Effect of homodimerization of the EpoR/IL-4Rα chimeric receptor and its Box1 mutants on Jak3 and Jak1 activities. Cells from intact EpoR/IL-4Rα transfectants (A and B) and EpoR/IL-4Rα Box1 mutants transfectants (B) were stimulated with Epo (300 units/ml) or IL-4 (1,200 units/ml) for the indicated periods of time (A) or for 10 min (B). Then cells were lysed as described in Materials and Methods. Cell lysates were immunoprecipitated with anti-Jak3 antibody (A) or anti-Jak1 antibody (B) and the immunoprecipitates were electrophoresed on SDS/PAGE, transferred to a nitrocellulose membrane, and immunoblotted with anti-phosphotyrosine (P-Tyr) mAb 4G10. The membrane was then stripped and reimmunoblotted with anti-Jak3 antibody (A) or anti-Jak1 antibody (B). Similar results were obtained in two independent experiments.
Disruption of Box1 in the EpoR/IL-4Rα Chimeric Receptor Abolishes Epo-Mediated Induction of Cɛ Germline Transcripts.
The membrane proximal proline-rich Box1 region of the IC domain of cytokine receptors is critical for association with Jak kinases (24), including Jak1 (25). To ascertain that Jak1 is involved in signal transduction following homodimerization of IL-4Rα, we examined the effect of disruption of Box1 in the EpoR/IL-4Rα chimeric construct on the induction of Cɛ transcripts. Two independent EpoR/IL-4Rα Box1 deletion mutants that lacked the sequences 237 WWDQIPNP 244 (Box1 mut 1) and 242 PNP 244 (Box1 mut 2) respectively, were stably transfected in BJAB cells. Expression of the mutated EpoR/IL-4Rα chimeric receptor was equivalent to that shown in Fig. 1A for BJAB cells stably transfected with EpoR/IL-4Rα (data not shown).
Fig. 4B shows that BJAB cell lines transfected with EpoR/IL-4Rα Box1 deletion mutants had high baseline tyrosine phosphorylation (lanes 4 and 7). Engagement of the native IL-4R complex by IL-4 induced increased Jak1 tyrosine phosphorylation in both of these cell lines (lanes 6 and 9). In contrast, homodimerization of the mutated EpoR/IL-4Rα chimeric receptors by Epo failed to induce tyrosine phosphorylation of Jak1 (lanes 5 and 8). These results strongly suggest that disruption of Box1 abolished the capacity of the mutant receptors to associate with and activate Jak1.
Fig. 5 shows that ligation of the mutated chimeric receptors with Epo failed to induce Cɛ germline transcripts, whereas ligation of the native IL-4R with IL-4 in the same cells induced it. These results suggest that Jak1 is critical for the induction of Cɛ germline transcripts following homodimerization of IL-4Rα chains.
Figure 5.
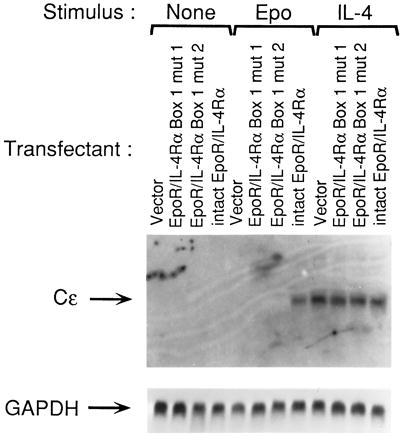
Disruption of the Jak1 binding proline-rich Box1 region of IL-4Rα abolishes the induction of Cɛ germline transcripts following homodimerization of the EpoR/IL-4Rα chimeric receptor . The cells transfected with empty pcDNA3 vector, EpoR/IL-4Rα Box1 deletion mutants (Box1 mut 1 or Box1 mut 2), or intact EpoR/IL-4Rα were left unstimulated (None), or stimulated with Epo (50 units/ml) or IL-4 (200 units/ml) for five days and assessed for induction of Cɛ germline transcripts and expression of glyceraldehyde-3-phosphate dehydrogenase (GAPDH) by Northern blot analysis. Similar results were obtained in two independent experiments.
DISCUSSION
Our data demonstrates that homodimerization of the IL-4Rα chain induces Cɛ germline transcripts in human B cells. This was accompanied by activation of Jak1 kinase and by induction of STAT6 nuclear activity and was abolished by disruption of the membrane proximal Box1 proline-rich region of IL-4Rα .
The present study was prompted by the observation that IL-4 induces Cɛ gene transcription and causes IgE synthesis in B cells from patients with X-SCID (15, 16). Induction of Cɛ gene transcription by IL-4 in these B cells may have involved signaling via the IL-13R complex, which shares the IL-4Rα chain with the IL-4R complex. We considered the possibility that the IL-4Rα chain by itself may be able to transduce an intracellular signal that leads to Cɛ gene transcription independently of its association with other chains. This was shown to be the case because ligation of a chimeric EpoR/IL-4Rα receptor expressed on B cells by Epo resulted in Cɛ gene transcription (Fig. 1B). This induction was dependent on the IL-4Rα chain in the chimeric receptor, because ligation by Epo of a transfected muEpoR failed to induce Cɛ gene transcripts in BJAB cells.
Since the rabbit anti-EpoR antibody we used in the FACS analysis to detect muEpoR expression was raised against and recognizes human EpoR, the lack of detectable EpoR expression on BJAB cells (Fig. 1A) strongly suggests that these cells do not express endogenous human EpoR protein. Moreover, we were unable to detect human EpoR mRNA in BJAB cells by reverse transcription–PCR analysis (data not shown). Thus, it is unlikely that induction of Cɛ gene transcripts by Epo-ligation of the chimeric EpoR/IL-4Rα receptor involved cooperative signaling by an endogenous EpoR and by the muEpoR component of the chimeric receptor.
Homodimerization of IL-4Rα was essential for induction of Cɛ gene transcripts because these transcripts were detected in cells expressing the chimeric EpoR/IL-4Rα receptor only after treatment with Epo. In contrast, Cɛ gene transcripts were constitutively expressed in B cells that expressed a chimeric CD8/IL-4Rα receptor as a homodimer (Fig. 2). The fact that Cɛ gene transcripts were induced by two independent models of IL-4Rα homodimerization strongly suggests that homodimerization of the intracellular domain of the IL-4Rα chain is sufficient to transduce a signal that results in Cɛ gene transcription.
STAT6 plays a critical role in Cɛ germline transcription in response to IL-4 as evidenced by the failure of B cells from mice homozygous for disruption of the STAT6 gene to transcribe the Cɛ gene in response to IL-4 (26). STAT6 binding to the sequence TTCCCAAGAA of the human germline ɛ promoter situated between −159 and −133 bp of the Iɛ exon is essential for the activation of Cɛ germline transcription in B cells, possibly by derepressing the promoter (27). Induction of STAT6 by IL-4 has been observed in B cells from patients with X-SCID by some authors (28) but not others (15). Induction of STAT6 activity by IL-4 in X-SCID B cells is consistent with the ability of IL-4 to induce IgE switching in these B cells (16). In our study, STAT6 nuclear activity was readily induced by homodimerization of the chimeric EpoR/IL-4Rα receptor (Fig. 3). Thus homodimerization of the IL-4Rα chain is sufficient to induce STAT6 activity.
As previously reported (29, 30, 31), ligation of the native IL-4R resulted in tyrosine phosphorylation of Jak3 (Fig. 4A) and Jak1 (Fig. 4B). Not surprisingly, homodimerization of the chimeric EpoR/IL-4Rα by Epo resulted in no detectable tyrosine phosphorylation of Jak3 (Fig. 4A). However it caused an increase in Jak1 tyrosine phosphorylation equivalent to that seen with ligation of the native IL-4R (Fig. 4B).
Dimerization of receptor chains that are associated with Jak kinases is thought to result in transphosphorylation and activation of these kinases. To date, receptor-mediated Jak1 activation has been accompanied by activation of other members of the Jak kinase family. We were unable to detect activation of any other known member of the Jak kinase family following ligation of the chimeric EpoR/IL-4Rα receptor. Our data suggests that following homodimerization of chimeric EpoR/IL-4Rα chains, Jak1 molecules would transphosphorylate and activate each other. Thus, Jak1 in the IL-4Rα homodimer may replace the function of Jak3 in the IL-4Rα/γc heterodimer. This is consistent with recent observations that the Jaks are interchangeable and indiscriminative in the Jak-STAT signaling pathway (32–34).
Two mutations that disrupted the presumptive Jak1 anchoring site of the proline-rich Box1 region in IL-4Rα abrogated Jak1 phosphorylation (Fig. 4B), STAT6 activation (data not shown) and Cɛ germline transcription following receptor homodimerization. These results suggest that Jak1 is required for the induction of Cɛ transcription following homodimerization of the IL-4Rα chain. We cannot formally rule out a role for proteins other than Jak1 that may bind to the Box1 region of IL-4Rα. Experiments in B cells transfected with a dominant negative Jak1 mutant or deficient in Jak1 are needed to definitively establish that Jak1 is essential for the induction of Cɛ gene transcription following homodimerization of the IL-4Rα chain.
While this manuscript was in preparation, two groups showed that homodimerization of human IL-4Rα chains transfected in 32D cells expressing the IRS-1 (murine promyeloid cell line) or Ba/F3 cells (murine pro-B cell line) result in cellular proliferation (28, 35). Thus, the IL-4Rα chain is principally responsible for directing at least two signaling events triggered by IL-4, proliferation and Cɛ gene transcription. Although there is no direct evidence for homomeric IL-4Rα receptors, homodimerization of the IL-4Rα chain by IL-4 could occur. The cytokine IL-2 has been shown to spontaneously form covalent dimers in vitro, possibly by transglutamination (36), and self-association of IL-2 bound to its receptor has been demonstrated (37). Given the structural similarity between IL-4 and IL-2, it is possible that IL-4 may also under certain conditions form dimers.
Acknowledgments
We thank Drs. A. D’Andrea, B. Mathey-Prevot, P. Leder, D. Seldin, E. Kieff, J. DeVries, and L. Clayton for their gifts of reagents and are grateful to Drs. A. D’Andrea, B. Mathey-Prevot, E. Tsitsikov, and Haifa Jabara for helpful suggestions and discussions. This work was supported by National Institutes of Health Grant AI-31136 and by grants from Baxter Healthcare, Quantum, and Alpha Therapeutics Corporations. H.F. is the recipient of a grant from the Japan Research Foundation for Clinical Pharmacology and of an American Academy of Allergy Asthma and Immunology Glaxo–Wellcome Respiratory Diseases Research Award.
ABBREVIATIONS
- IL-4R
IL-4 receptor
- γc
IL-2Rγ
- Epo
erythropoietin
- muEpoR
murine erythropoietin receptor
- Jak
Janus kinase
- STAT
signal transducer and activator of transcription
- X-SCID
X-linked severe combined immunodeficiency
- EC
extracellular
- TM
transmembrane
- IC
intracellular
- NRS
normal rabbit serum
- FACS
fluorescence-activated cell sorter
References
- 1.Vercelli D, Geha R S. Curr Opin Immunol. 1992;4:794–797. doi: 10.1016/0952-7915(92)90064-l. [DOI] [PubMed] [Google Scholar]
- 2.Defrance T, Carayon P, Billian G, Guillemot J-C, Minty A, Caput D, Ferrara P. J Exp Med. 1994;179:135–143. doi: 10.1084/jem.179.1.135. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Zurawski S M, Chomarat P, Djossous O, Bidaud C, McKenzie P, Miossec A N J, Banchereau J, Zurawski G. J Biol Chem. 1995;270:13869–13878. doi: 10.1074/jbc.270.23.13869. [DOI] [PubMed] [Google Scholar]
- 4.Kondo M, Takeshita T, Ishii N, Nakamura M, Watanabe S, Arai K, Sugamura K. Science. 1993;262:1874–1877. doi: 10.1126/science.8266076. [DOI] [PubMed] [Google Scholar]
- 5.De Saint Basile G, Di Santo J P, Fischer A. Clin Immunol Immunopathol. 1995;76:S165–S167. doi: 10.1016/s0090-1229(95)90090-x. [DOI] [PubMed] [Google Scholar]
- 6.Noguchi M, Yi H, Rosenblatt H M, Filipovich A H, Adelstein S, Modi W S, McBride O W, Leonard W J. Cell. 1993;73:147–157. doi: 10.1016/0092-8674(93)90167-o. [DOI] [PubMed] [Google Scholar]
- 7.Miyazaki T, Kawahara A, Fujii H, Nakagawa Y, Minami Y, Liu Z-J, Oishi I, Silvennoinen O, Witthuhn B A, Ihle J N, Taniguchi T. Science. 1994;266:1045–1047. doi: 10.1126/science.7973659. [DOI] [PubMed] [Google Scholar]
- 8.Smerz-Bertling C, Duschl A. J Biol Chem. 1995;270:966–970. doi: 10.1074/jbc.270.2.966. [DOI] [PubMed] [Google Scholar]
- 9.Hou J, Schindler U, Henzel W J, Ho T C, Brasseur M, McKnight S L. Science. 1994;265:1701–1706. doi: 10.1126/science.8085155. [DOI] [PubMed] [Google Scholar]
- 10.Ivashkiv L B. Immunity. 1995;3:1–4. doi: 10.1016/1074-7613(95)90152-3. [DOI] [PubMed] [Google Scholar]
- 11.Fenghao X, Saxon A, Nguyen A, Ke Z, Diaz-Sanchez D, Nel A. J Clin Invest. 1995;96:907–914. doi: 10.1172/JCI118138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Hilton D J, Zhang J-G, Metcalf D, Alexander W S, Nicola N A, Willson T A. Proc Natl Acad Sci USA. 1996;93:497–501. doi: 10.1073/pnas.93.1.497. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Caput D, Laurent P, Kaghad M, Lelias J, Lefort S, Vita N, Ferrara P. J Biol Chem. 1996;271:16921–16926. doi: 10.1074/jbc.271.28.16921. [DOI] [PubMed] [Google Scholar]
- 14.He Y-W, Malek T R. J Immunol. 1995;155:9–12. [PubMed] [Google Scholar]
- 15.Izuhara K, Heike T, Otsuka T, Yamaoka K, Mayumi M, Imamura T, Niho Y, Harada N. J Biol Chem. 1996;271:619–622. doi: 10.1074/jbc.271.2.619. [DOI] [PubMed] [Google Scholar]
- 16.Matthews D J, Clark P A, Herbert J, Morgan G, Armitage R J, Kinnon C, Minty A, Grabstein K H, Caput D, Ferrara P, Callard R. Blood. 1995;85:38–42. [PubMed] [Google Scholar]
- 17.Ho S N, Hunt H D, Horton R M, Pullen J K, Pease L R. Gene. 1989;77:51–59. doi: 10.1016/0378-1119(89)90358-2. [DOI] [PubMed] [Google Scholar]
- 18.Ichiki T, Takahashi W, Watanabe T. Int Immunol. 1992;4:747–754. doi: 10.1093/intimm/4.7.747. [DOI] [PubMed] [Google Scholar]
- 19.Tsytsykova A, Tsitsikov E, Ramesh N, Geha R S. J Biol Chem. 1996;271:3763–3770. doi: 10.1074/jbc.271.7.3763. [DOI] [PubMed] [Google Scholar]
- 20.Yin T, Keller S R, Quelle F W, Witthuhn B A, Tsang M L-S, Lienhard G E, Ihle J N, Yang Y-C. J Biol Chem. 1995;270:20497–20502. doi: 10.1074/jbc.270.35.20497. [DOI] [PubMed] [Google Scholar]
- 21.Miura O, Cleveland J L, Ihle J N. Mol Cell Biol. 1993;13:1788–1795. doi: 10.1128/mcb.13.3.1788. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Hennecke S, Cosson P. J Biol Chem. 1993;268:26607–26612. [PubMed] [Google Scholar]
- 23.Moebius U, Kober G, Griscelli A L, Hercend V, Meuer S C. Eur J Immunol. 1991;21:1793–1800. doi: 10.1002/eji.1830210803. [DOI] [PubMed] [Google Scholar]
- 24.Ihle J N. Adv Immunol. 1995;60:1–35. doi: 10.1016/s0065-2776(08)60582-9. [DOI] [PubMed] [Google Scholar]
- 25.Higuchi M, Asao H, Tanaka N, Oda K, Takeshita T, Nakamura M, Van Snick J, Sugamura K. Eur J Immunol. 1996;26:1322–1327. doi: 10.1002/eji.1830260622. [DOI] [PubMed] [Google Scholar]
- 26.Takeda K, Tanaka T, Shi W, Matsumoto M, Minami M, Kashiwamura S, Nakanishi K, Yoshida N, Kishimoto T, Akira S. Nature (London) 1996;380:627–630. doi: 10.1038/380627a0. [DOI] [PubMed] [Google Scholar]
- 27.Wang D Z, Cherrington A, Famakin-Mosuro B, Boothby M. Int Immunol. 1996;8:977–989. doi: 10.1093/intimm/8.7.977. [DOI] [PubMed] [Google Scholar]
- 28.Lai S Y, Molden J, Liu K D, Puck J M, White M D, Goldsmith M A. EMBO J. 1996;15:4506–4514. [PMC free article] [PubMed] [Google Scholar]
- 29.Witthuhn B A, Silvennoinen O, Miura O, Lai K S, Cwik C, Liu E T, Ihle J N. Nature (London) 1994;370:153–157. doi: 10.1038/370153a0. [DOI] [PubMed] [Google Scholar]
- 30.Johnston J A, Kawamura M, Kirken R A, Chen Y-Q, Blake T B, Shibuya K, Ortaldo J R, McVicar D W, O’Shea J J. Nature (London) 1994;370:151–153. doi: 10.1038/370151a0. [DOI] [PubMed] [Google Scholar]
- 31.Rolling C, Treton D, Beckmann P, Galanaud P, Richard Y. Oncogene. 1995;10:1757–1761. [PubMed] [Google Scholar]
- 32.Kotenko S V, Izotova L S, Pollack B P, Muthukumaran G, Paukkut K, Silvennoinen O, Ihle J N, Pestka S. J Biol Chem. 1996;271:17174–17182. doi: 10.1074/jbc.271.29.17174. [DOI] [PubMed] [Google Scholar]
- 33.Jiang N, He T, Miyajima A, Wojchowski D M. J Biol Chem. 1996;271:16472–16476. doi: 10.1074/jbc.271.28.16472. [DOI] [PubMed] [Google Scholar]
- 34.Lai S Y, Xu W, Gaffen S L, Liu K D, Longmore G D, Greene W C, Goldsmith M A. Proc Natl Acad Sci USA. 1996;93:231–235. doi: 10.1073/pnas.93.1.231. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Kammer W, Lischke A, Moriggl R, Groner B, Ziemiecki A, Gurniak C B, Berg L J, Friedrich K. J Biol Chem. 1996;271:23634–23637. doi: 10.1074/jbc.271.39.23634. [DOI] [PubMed] [Google Scholar]
- 36.Bergmann C A, Landmeier B J, Kaplan D R. Mol Immunol. 1991;28:99–105. doi: 10.1016/0161-5890(91)90092-x. [DOI] [PubMed] [Google Scholar]
- 37.Kaplan D, Smith D, Huang R, Yildirim Z. FASEB J. 1995;9:1096–1102. doi: 10.1096/fasebj.9.11.7649410. [DOI] [PubMed] [Google Scholar]